I Nutrienti. I grassi. Prof. Giorgio Sartor. L04 - Versione 1.3 oct Copyright by Giorgio Sartor. All rights reserved.
|
|
|
- Bruno Fedele
- 8 anni fa
- Visualizzazioni
Transcript
1 Prof. Giorgio artor I utrienti I grassi Copyright by Giorgio artor. All rights reserved. L04 - Versione 1.3 oct 2011 V.1.3 gsartor L04 - I grassi - 2-1

2 Metabolismo dei grassi I grassi sono fra le principali fonti di energia metabolica Grassi Proteine Glicogeno (fegato) Glicogeno (muscolo) Glucosio Energia (kjmole -1 ) V.1.3 gsartor L04 - I grassi I grassi (esteri del glicerolo) C1 C 3 C2 C3 + 3 C C 3 C3 C 3 C 3 2 C C 2 R P C 3 C 3 Trigliceridi (TAG) Fosfolipidi Lisofosfolipidi + 3 C C 3 C3 P C 3 V.1.3 gsartor L04 - I grassi - 4-2
 C1 C 3 C2 C3 + 3 C C 3")
3 I grassi (esteri della sfingosina) C 3 2 C 3 C 3 fingosina Ceramidi fingomielina P C 3 C C C 3 3 C V.1.3 gsartor L04 - I grassi Colesterolo 3 C 3 C C 3 3 C C 3 V.1.3 gsartor L04 - I grassi - 6-3

4 ello spazio! V.1.3 gsartor L04 - I grassi Bilayer I fosfolipidi formano bilayers. Liposomi: V.1.3 gsartor L04 - I grassi - 8-4

5 Le cellule hanno membrane lipidiche! V.1.3 gsartor L04 - I grassi -9- Digestione dei lipidi e trasporto A causa dell insolubilità in acqua dei lipidi la loro digestione ed il trasporto sono un problema chimico. Gli enzimi che agiscono sui lipidi sono enzimi solubili o di membrana che agiscono all interfaccia acqua particella lipidica. I lipidi ed i prodotti di digestione (insolubili o poco solubili in acqua) devono essere trasportati attraverso compartimenti acquosi all interno di cellule e tessuti per essere metabolizzati. V.1.3 gsartor L04 - I grassi

6 Detergenti L accessibilità dei legami esterei dei TAG è favorita dalla presenza di sali di acili biliari che funzionano come emulsionanti: 3 C C 3 Acido colico (colato) C 3 C 3 EI MAMMIFERI R R Acido taurocolico Acido glicocolico V.1.3 gsartor L04 - I grassi Acidi biliari in ambiente marino R" C 3 R' R' = Ac R" = A C 3 R' = R" = A R' = R" = B C 3 C 3 R R = A R = B Derivati dell acido 3-oxo-col-4-en- 24-oico e 3-oxo-col-1,4-dien-24- oico A 3 C B 3 C * * ew 3-xo-chol-4-en-24-oic Acids from the Marine oft Coral Eleutherobia sp. arah C. Lievens, a kon ope, and Tadeusz F. Molinski* J. at. Prod. 2004, 67, V.1.3 gsartor L04 - I grassi

7 Lipasi Catalizza l idrolisi dei triacilgliceroli in posizione 1 e 3 formando 1,2-diacilgliceroli, 2-acilglicerolo e quindi glicerolo C1 C2 C3 C 3 C 3 2 C 3 C 3 2 C 3 C 3 C 3 2 C 3 C 3 V.1.3 gsartor L04 - I grassi Lipolisi: meccanismo generale R' R" R' u+ R" R u: R R' + R" R' u R" R u: 2 R V.1.3 gsartor L04 - I grassi

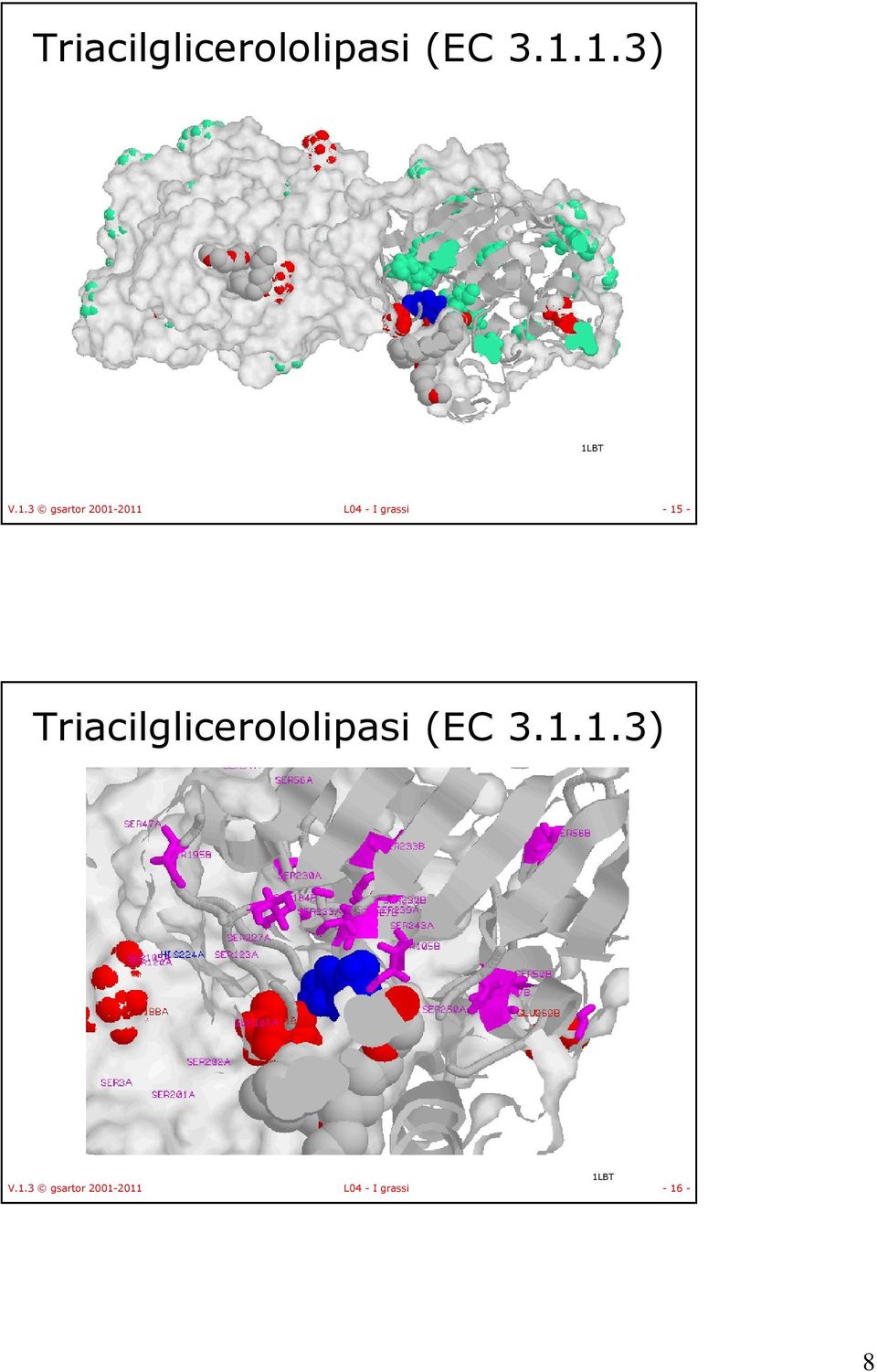
8 Triacilglicerololipasi (EC ) 1LBT V.1.3 gsartor L04 - I grassi Triacilglicerololipasi (EC ) 1LBT V.1.3 gsartor L04 - I grassi
9 Lipasi (EC ) Triade catalitica er is Asp 1CUA V.1.3 gsartor L04 - I grassi Lipasi come antidoto! Alcune alghe della famiglia Caulerpaceae (Caulerpa prolifera, C. taxifolia e C. racemosa) producono caulerpenina, un metabolita tossico (sesquiterpene) in grado di funzionare come deterrente chimico nei confronti di molluschi marini che vivono nella comunità algale. Una mollusco (xynoe olivacea) riesce a rendere inoffensivo il terpene attraverso una lipasi. Lipase-mediated production of defensive toxins in the marine mollusc xynoe olivacea Adele Cutignano, Valentina otti, Giuliana d Ippolito, Anna Domènech Coll, Guido Cimino and Angelo Fontana* rg.biomol.chem.,2004,2, V.1.3 gsartor L04 - I grassi
 riesce a rendere inoffensivo il terpene attraverso una lipasi.")
10 Lipasi come antidoto! Alcune alghe della famiglia Caulerpaceae (Caulerpa prolifera, C. taxifolia e C. racemosa) producono caulerpenina, un metabolita tossico (sequiterpene) in grado di funzionare come deterrente chimico nei confronti di molluschi marini che vivono nella comunità algale. Una mollusco (xynoe olivacea) riesce a rendere inoffensivo, per sé, il terpene attraverso una lipasi. Lipase-mediated production of defensive toxins in the marine mollusc xynoe olivacea Adele Cutignano, Valentina otti, Giuliana d Ippolito, Anna Domènech Coll, Guido Cimino and Angelo Fontana* rg.biomol.chem.,2004,2, V.1.3 gsartor L04 - I grassi Lipasi come antidoto! Alcune alghe della famiglia Caulerpaceae (Caulerpa prolifera, C. taxifolia e C. racemosa) producono caulerpenina, un metabolita tossico (sequiterpene) in grado di funzionare come deterrente chimico nei confronti di molluschi marini che vivono nella comunità algale. Una mollusco (xynoe olivacea) riesce a rendere inoffensivo, per sé, il terpene attraverso una lipasi. Lipase-mediated production of defensive toxins in the marine mollusc xynoe olivacea Adele Cutignano, Valentina otti, Giuliana d Ippolito, Anna Domènech Coll, Guido Cimino and Angelo Fontana* rg.biomol.chem.,2004,2, V.1.3 gsartor L04 - I grassi

11 Lipasi come antidoto! Alcune alghe della famiglia Caulerpaceae (Caulerpa prolifera, C. taxifolia e C. racemosa) producono caulerpenina, un metabolita tossico (sequiterpene) in grado di funzionare come deterrente chimico nei confronti di molluschi marini che vivono nella comunità algale. Una mollusco (xynoe olivacea) riesce a rendere inoffensivo, per sé, il terpene attraverso una lipasi. TIE PER LA DIFEA DAI PREDATRI V.1.3 gsartor L04 - I grassi Meccanismo Lipase-mediated production of defensive toxins in the marine mollusc xynoe olivacea Adele Cutignano, Valentina otti, Giuliana d Ippolito, Anna Domènech Coll, Guido Cimino and Angelo Fontana* rg.biomol.chem.,2004,2, V.1.3 gsartor L04 - I grassi

12 Meccanismo Lipase-mediated production of defensive toxins in the marine mollusc xynoe olivacea Adele Cutignano, Valentina otti, Giuliana d Ippolito, Anna Domènech Coll, Guido Cimino and Angelo Fontana* rg.biomol.chem.,2004,2, V.1.3 gsartor L04 - I grassi Fosfolipasi C 3 PLD + C 3 C 3 P PLC PLA 2 C 3 C 3 PLA 1 V.1.3 gsartor L04 - I grassi

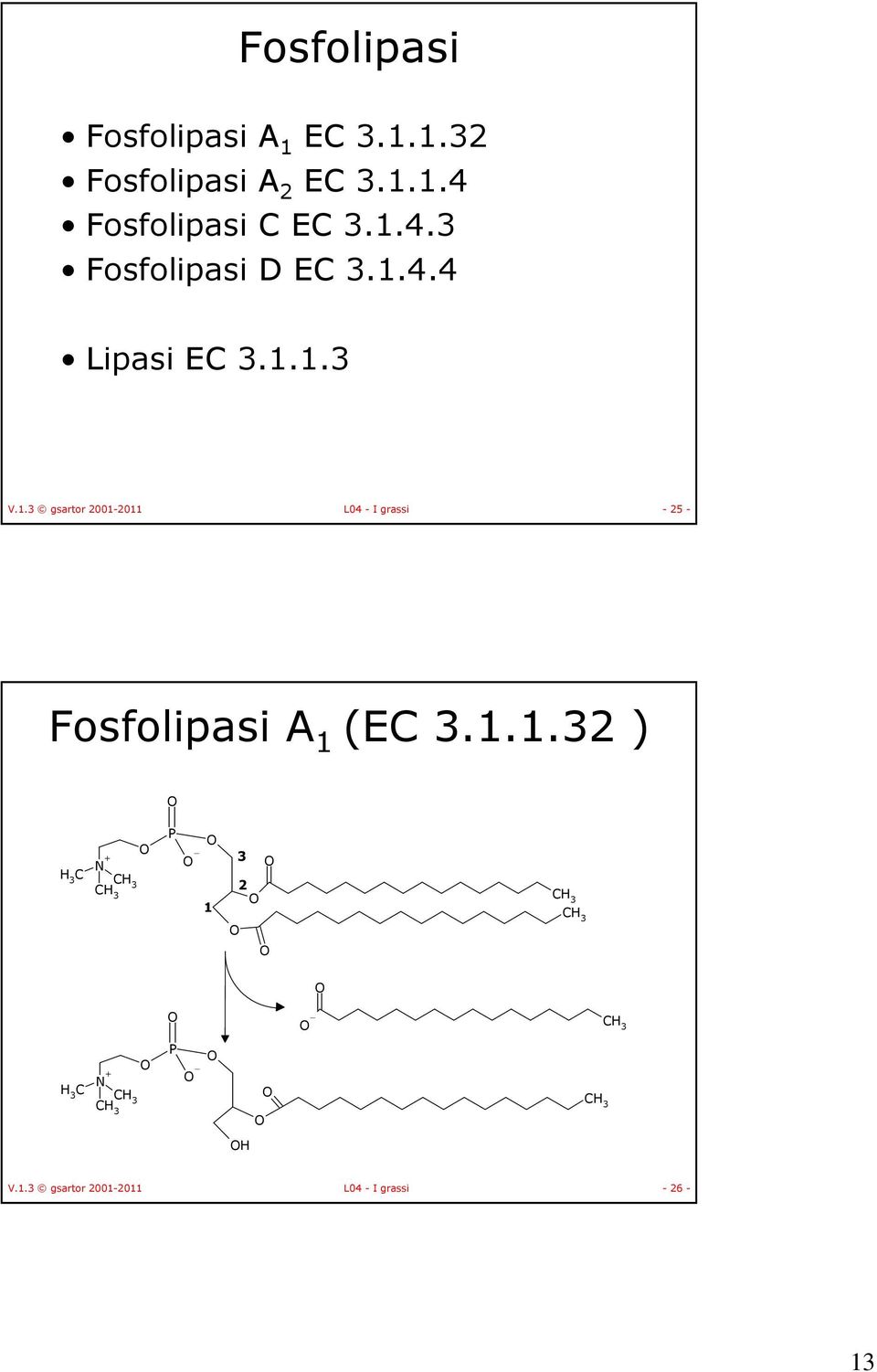
13 Fosfolipasi Fosfolipasi A 1 EC Fosfolipasi A 2 EC Fosfolipasi C EC Fosfolipasi D EC Lipasi EC V.1.3 gsartor L04 - I grassi Fosfolipasi A 1 (EC ) C 3 + C 3 C 3 P C 3 C 3 C 3 3 C + C 3 C 3 P C 3 V.1.3 gsartor L04 - I grassi
14 Fosfolipasi A 1 Ca ++ AP I V.1.3 gsartor L04 - I grassi QD6 Meccanismo (diade catalitica) is48 Asp99 Ca++ R' R" is48 Ca++ R" is48 Ca++ Asp99 R' Asp99 R' R" V.1.3 gsartor L04 - I grassi
 is48 Asp99 Ca++ R' R\"")
15 Fosfolipasi A 2 (EC EC ) C 3 + C 3 C 3 P C 3 C 3 C 3 3 C + C 3 P C 3 C 3 V.1.3 gsartor L04 - I grassi Fosfolipasi A 2 Il lisofosfolipide è un detergente potentissimo, La fosfolipasi A 2 è contenuta nel veleno di serpenti (cobra), insetti (api), probabilmente anche in invertebrati marini (spugne), l effetto è quello di lisare i globuli rossi attraverso l effetto della lisofosfatidilcolina ei mammiferi è secreta dal pancreas e una piccola quantità di lecitina viene secreta dal fegato, e quindi idrolizzata, per aiutare la solubilizzazione dei grassi. V.1.3 gsartor L04 - I grassi

16 Fosfolipasi A 2 Esistono almeno due famiglie di PLA 2 : spla 2 : secreta, che permette l idrolisi dei fosfolipidi all interfaccia, e possiede un dominio che si lega alla membrana (veleno delle api) cpla 2 : citosolica, che viene utilizzata per la produzione di acidi grassi (arachidonato), inositolo fosfato come messaggeri intracellulari. V.1.3 gsartor L04 - I grassi Fosfolipasi A 2 (EC ) 18G V.1.3 gsartor L04 - I grassi
 18G V.1.3 gsartor 2001-2011 L04 - I grassi - 32-16")
17 V.1.3 gsartor L04 - I grassi Diade catalitica Ca ++ AP99 I48 1FXF V.1.3 gsartor L04 - I grassi

18 V.1.3 gsartor L04 - I grassi Ipotetico modo d azione di Fosfolipasi A 2 su di una micella di fosfolipidi V.1.3 gsartor L04 - I grassi

19 FAB V.1.3 gsartor L04 - I grassi FAB V.1.3 gsartor L04 - I grassi

20 Glicerolo L ossidazione del glicerolo a diidrossiacetone è catalizzata dalla glicerolo fosfato (EC ) AD+ AD 1JQ5 V.1.3 gsartor L04 - I grassi Glicerolo ATP ADP P La fosforilazione del diidrossiacetone è catalizzata dalla diidrossiacetone chinasi (EC ). Il diidrossiacetonfosfato così formatosi è convogliato nella glicolisi 1UE Diidrossiacetone mologo ATP Mg V.1.3 gsartor L04 - I grassi

21 Glicerolo ATP ADP P La fosforilazione del diidrossiacetone è catalizzata dalla diidrossiacetone chinasi (EC ). Il diidrossiacetonfosfato così formatosi è convogliato nella glicolisi 1UE Diidrossiacetone mologo ATP Mg ++ V.1.3 gsartor L04 - I grassi β-ossidazione degli acidi grassi 21
22 β-ossidazione degli acidi grassi Ì?? Acidi grassi Acidi grassi saturi C 3 Acido stearico (C 18 ) Acidi grassi insaturi C 3 Acidi grassi polinsaturi Acido oleico (C 18 9 ) C18:1ω C 3 18 Acido linoleico (C 18 9,12 ) C18:2ω6 V.1.3 gsartor L04 - I grassi
23 β-ossidazione acidi grassi Una volta che i TGA sono idrolizzati ad acidi grassi questi ultimi vengono demoliti fondamentalmente attraverso l ossidazione, Il principale meccanismo è quello della β-ossidazione che distacca unità bicarboniose, sottoforma di Acetil-, rompendo il legame tra il Cα e il Cβ, α β C 3 C 3 α β C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi A seconda del numero (n) di atomi di carbonio dell acido grasso di producono n/2 molecole di Acetil-. La reazione avviene nella matrice mitocondriale. Gli acidi grassi vengo attivati attraverso la formazione di un Acil-. L Acil- viene trasportato all interno della membrana interna mitocondriale attraverso il sistema carnitina/acilcarnitina. V.1.3 gsartor L04 - I grassi
24 Attivazione degli acidi grassi L attivazione degli acidi grassi porta alla formazione di Acil- Coa Avviene nel citoplasma ad opera di una l acil- sintetasi (EC ). Il processo è endoergonico ( G = 31.4 kjmole -1 ) Viene reso spontaneo dalla razione di idrolisi di una molecola di ATP a AMP e PPi, il quale fornisce un surplus di energia convertendosi a 2Pi attraverso una pirofosfatasi, per un totale di kjmole -1 di ATP impiegato. C 3 ATP G = kj mol -1 G = kj mol -1 G = kj mol AMP + PPi -1 2Pi C 3 1V25 1V26 V.1.3 gsartor L04 - I grassi Formazione di Acil- Il meccanismo di reazione prevede la formazione di un derivato intermedio aciladenosina, legato all enzima, che subisce l attacco nucleofilo da parte dell atomo di zolfo del -. ATP Acido grasso C 3 P P P n C 3 n + P P P PPi P Aciladenosina Pi C 3 n P V.1.3 gsartor L04 - I grassi Intermedio tetraedrico 2 C 3 P n Acil- + AMP 2 24
25 Trasporto Il trasporto degli acil nell interno degli organelli subcellulari, dove avviene l ossidazione degli acidi grassi, sfrutta il sistema carnitina/acilcarnitina. + C 3 C 3 C 3 V.1.3 gsartor L04 - I grassi Carnitina Il sistema carnitina/acilcarnitina è stato scoperto la prima volta nei mitocondri. Esso contiene carnitina acetiltransferasi (CAT), legata strettamente alla faccia interna della membrana interna mitocondriale e quattro isoenzimi carnitina palmitoil transferasi (CPT, CPT-IA nel fegato, CPT-IB nel muscolo e altre cellule, CPT-IC nel cervello e CPT-II). Gli enzimi CPT-I sono localizzati nella membrana esterna mitocondriale con le loro porzioni catalitica e regolatoria (inibita da malonil) che si affacciano verso il lato citoplasmatico mentre CPT II è localizzata come la CAT. Tale varietà isoenzimatica è legata alla diversa affinità per i diversi gruppi acilici: CAT: C2-C10, CPTI: C6-C20 CPTII: C6-C18 V.1.3 gsartor L04 - I grassi
26 Carnitina Anche i perossisomi contengono una CAT, la carnitina octiltransferasi (CRT) che ha un affinità maggiore per C4-C16, è inibita da malonil ed un altra acilcarnitina transferasi solubile non inibita da malonil. Anche altre membrane (reticolo endoplasmatico, sarcoplasmatico, membrana nucleare e membrana plasmatica) contengono carnitina aciltransferasi. V.1.3 gsartor L04 - I grassi Trasporto nei mitocondri Il trasporto di Acil-Coa attraverso la membrana interna mitocondriale viene quindi mediato dalla carnitina attraverso tre sistemi enzimatici: A. Carnitina aciltransferasi I, nel lato citoplasmatico, CAT, CPT-IA nel fegato, CPT-IB nel muscolo e altre cellule, CPT-IC nel cervello. B. Carnitina acilcarnitina traslocasi (CACT), nella membrana e C. Carnitina aciltransferasi II, nel lato della matrice mitocondriale. CPT-II Carnitina Carnitina Acil- Citoplasma CPTI -acilcarnitina - pazio intermembrana CACT CPTII Matrice Acil- -acilcarnitina Carnitina V.1.3 gsartor L04 - I grassi
27 Trasporto nei mitocondri Carnitina Acil- -acetilcarnitina Citoplasma CPTI Carnitina -acetilcarnitina Carnitina -acilcarnitina - pazio intermembrana CACT CACT Matrice CPTII β-ossidazione CAT -acilcarnitina + - Acil- + Carnitina Acetil- + Carnitina -acetilcarnitina + - Carnitina V.1.3 gsartor L04 - I grassi Trasporto nei perossisomi ei perossisomi il trasporto di Acil-Coa attraverso la membrana viene mediato dalla carnitina attraverso: A. Carnitina aciltransferasi I, nel lato citoplasmatico, CRT e solubile. B. Carnitina acilcarnitina traslocasi, nella membrana e C. Carnitina aciltransferasi II, all interno. CPT-II Carnitina + Acil- CPT solubile CRT Interno del perossisoma - + -acilcarnitina CACT Carnitina + Acil- CPTII Citoplasma - + -acilcarnitina V.1.3 gsartor L04 - I grassi
28 Carnitina aciltransferasi EC C 3 C 3 C 3 + n C3 Carnitina Acil transferasi I (lato matrice) Carnitina Acil transferasi II (lato citoplasma) n C 3 + C 3 3 C C 3 + V.1.3 gsartor L04 - I grassi Carnitina aciltransferasi EC Carnitina V.1.3 gsartor L04 - I grassi
29 Carnitina aciltransferasi EC Carnitina V.1.3 gsartor L04 - I grassi Carnitina aciltransferasi EC V.1.3 gsartor L04 - I grassi
30 Carnitina aciltransferasi EC Carnitina V.1.3 gsartor L04 - I grassi Carnitina aciltransferasi EC Carnitina V.1.3 gsartor L04 - I grassi
31 β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
32 β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
33 β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC β-chetoacil- AD + + L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi La strategia della β-ossidazione consiste nel generare un nuovo carbonile in β a seguito della rotture del legame tra Cα e Cβ. Tale sequenza di reazioni produce: Un Acetil- Un Acil- più corto di due unità carboniose. Un FAD 2 Un AD La reazione procede ciclicamente fino a quando l acil- è ridotto a due unità (Acetil-) o tre unità (Propionil-) carboniose. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
34 Destino dei prodotti I prodotti della β-ossidazione: Acetil- e corpi chetonici: entrano nel ciclo di Krebs per produrre equivalenti riducenti (AD e FAD 2 ) che alimentano la fosforilazione ossidativa per la produzione di ATP. Un Acil- più corto di due unità carboniose: rientra nel ciclo successivo di β- ossidazione. FAD 2 e AD che alimentano la fosforilazione ossidativa per la produzione di ATP. V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
35 β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
36 β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra deidorgenasi riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - AD + + C 3 3-Chetotiolasi EC β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra deidorgenasi riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. FAD Acil- C 3 Acil- (C n ) EC EC EC C 3 FAD 2 C 3 3-Chetotiolasi EC Acil- (C n-2 ) - C 3 E il ciclo si ripete trans- 2 -enoil- C 3 β-chetoacil- AD L-3-idrossiacil- EC AD + L-β-idrossiacil- C 3 Enoil- idratasi EC V.1.3 gsartor L04 - I grassi
37 β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi Acil- EC EC EC C Tre enzimi solubili nella matrice 3 Acil- (C n ) - Contengono un FAD come gruppo C 3 β-chetoacil- mitocondriale con diversa 3-Chetotiolasi specificità per EC acidi grassi con catena idrofobica di diversa lunghezza (corti, medi e lunghi). prostetico. AD + + L-3-idrossiacil- EC AD + C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
38 Acil- EC EC EC Il FAD viene ridotto a FAD 2, gli elettroni vengono poi convogliati alla catena respiratoria attraverso una flavoproteina trasportatrice di elettroni (ETF) e al CoQ per formare CoQ 2. Acil- (C n ) EC EC EC C 3 FAD ETF red CoQ trans- 2 -enoil- FAD 2 ETF ox CoQ 2 C 3 V.1.3 gsartor L04 - I grassi Acil- EC EC EC Meccanismo C 3 C C 3 B B + FAD C 3 C C 3 B + EC I2 V.1.3 gsartor L04 - I grassi
39 β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + L-3-idrossiacil- EC AD + β-chetoacil- Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 C 3 Enoil- idratasi EC V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi Idratazione stereo specifica. L enoil- idratasi catalizza l idratazione stereospecifica al doppio legame trans producendo L-idrossiacil-. Esamero. C 3 β-chetoacil- AD + + L-3-idrossiacil- EC AD + Acil- (C n ) C 3 3-Chetotiolasi Due enzimi: EC EC : maggiore affinità per enoil- a corta - catena. EC : maggiore affinità per enoil- a lunga C catena. 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 C 3 Enoil- idratasi EC EC V.1.3 gsartor L04 - I grassi
40 Enoil- idratasi EC Esanoil- 1MJ3 V.1.3 gsartor L04 - I grassi Enoil- idratasi EC δ ++ R R Glu 164 Glu 144 Glu 164 δ R Glu 144 Glu 164 R Glu164 Glu 144 Glu144 Glu 144 Glu 164 V.1.3 gsartor L04 - I grassi
41 Enoil- idratasi EC δ ++ R R Glu 164 Glu 144 Glu 164 δ R Glu 144 Glu 164 R Glu164 Glu 144 Glu144 Glu 144 Glu 164 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + L-3-idrossiacil- EC AD + β-chetoacil- Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi
42 β-ossidazione acidi grassi L enzima L-3-idrossiacil- catalizza l ossidazione di L-β-idrossiacil-. FAD Acil- C 3 È un enzima specifico per l isomero L. Acil- (C n ) EC EC EC Il AD prodotto entra nella catena C 3 respiratoria a livello del FAD complesso I. 2 C 3 3-Chetotiolasi EC Acil- (C n-2 ) - C 3 C 3 trans- 2 -enoil- β-chetoacil- 2 AD + + L-3-idrossiacil- EC AD + L-β-idrossiacil- C 3 Enoil- idratasi EC V.1.3 gsartor L04 - I grassi L-3-idrossiacil- EC is Glu + δ - C AD + + P P Gly is + Glu C AD P P Gly V.1.3 gsartor L04 - I grassi
43 β-ossidazione acidi grassi In tutto quattro attività enzimatiche catalizzano il processo ciclico: Una riduce il legame CαCβ a doppio legame, Una idratasi addiziona acqua in trans per formare un enolo Un altra riduce il gruppo a carbonile ed infine Una tiolasi permette la rottura del legame CαCβ per formare Acetil- e Acil- più corto di due unità. - C 3 3-Chetotiolasi EC AD + + β-chetoacil- L-3-idrossiacil- EC AD + Acil- (C n ) C 3 C 3 Acil- (C n-2 ) L-β-idrossiacil- C 3 trans- 2 -enoil- C 3 FAD Acil- EC EC EC FAD 2 2 Enoil- idratasi EC C 3 V.1.3 gsartor L04 - I grassi β-ossidazione acidi grassi - β-chetoacil- Acil- (C n ) L-3-idrossiacil- Enoil- idratasi EC i produce un Acil-EC di due atomi di carbonio più corto. AD + C 3 C 3 3-Chetotiolasi EC AD + + C 3 Acil- (C n-2 ) C 3 L-β-idrossiacil- C 3 FAD Acil- EC EC EC FAD 2 trans- 2 -enoil- La chetotiolasi catalizza la scissione del β-chetoacil-. Il ciclo ricomincia. 2 C 3 V.1.3 gsartor L04 - I grassi
44 3-Chetotiolasi EC C n B + C 2 B + C 2 - B + C 3 C 2 B Intermedio enzima tioestere V.1.3 gsartor L04 - I grassi Chetotiolasi EC B B + Intermedio enzima tioestere B + C n-2 V.1.3 gsartor L04 - I grassi
45 3-Chetotiolasi EC Arg Cys 1AWF V.1.3 gsartor L04 - I grassi techiometria Per ogni ciclo si producono: AD che equivale a 2.5 moli di ATP prodotte nella catena respiratoria. FAD 2 che equivale a 1.5 moli di ATP prodotte nella catena respiratoria. Per la degradazione dell acido palmitico (C 16 ): C 3 -(C 2 ) 14 -C-- + 7FAD + 7AD C 3 -C-- + 7FAD 2 + 7AD FAD Pi ADP FAD ATP 320 kjmol -1 7AD Pi ADP AD ATP 534 kjmol -1 8C 3 -C Pi + 80ADP C ATP 2440 kjmol -1 C 3 -(C 2 ) 14 -C Pi +108ADP ATP + 16 C kjmol -1 V.1.3 gsartor L04 - I grassi
46 techiometria Per ogni ciclo si producono: AD che equivale a 2.5 moli di ATP prodotte nella catena respiratoria. FAD 2 che equivale a 1.5 moli di ATP prodotte nella catena respiratoria. Per la degradazione dell acido palmitico (C 16 ): C 3 -(C 2 ) 14 -C-- + 7FAD + 7AD C 3 -C-- + 7FAD 2 + 7AD FAD Pi ADP FAD ATP 320 kjmol -1 7AD Pi ADP AD ATP 534 kjmol -1 8C 3 -C Pi + 80ADP C ATP 2440 kjmol -1 C 3 -(C 2 ) 14 -C Pi +108ADP ATP + 16 C kjmol -1 V.1.3 gsartor L04 - I grassi Rendimento Combustione dell acido palmitico: C 3 -(C 2 ) 14 -C + 2 C G = kjmol -1 ssidazione dell acido palmitico e respirazione cellulare: C 3 -(C 2 ) 14 -C + 2 C (108 2)ATP = G = kjmol -1 Efficienza = 3233/9790 = 33% Produzione di metabolica per mole di acido palmitico che viene β-ossidato. V.1.3 gsartor L04 - I grassi
47 Pari e dispari Dalla β-ossidazione di acidi grassi con atomi si carbonio pari (n) si formano n/2 acetil- attraverso (n/2 1) cicli di reazioni. Dalla β-ossidazione di acidi grassi con atomi si carbonio dispari (n) si formano n/2-1 acetil- + 1 propionil- da (n/2-1) cicli di reazioni. V.1.3 gsartor L04 - I grassi Acidi grassi dispari Acil- β-ossidazione n/2 3 C C 3 Propionil- uccinil- Propionil- carbossilasi ATP + C Metilmalonil- mutasi D-Metilmalonil- C 3 C 3 Metilmalonil- epimerasi L-Metilmalonil- V.1.3 gsartor L04 - I grassi
48 Acidi grassi monoinsaturi 3 C 6 6 C 3 leil- (C 18 9 ) 6 3 x β-ossidazione 3 3 C 2 Enoil- idratasi 6 x β-ossidazione 3 C 6 3 C 6 cis- 3 -dodecanoil- Enoil- isomerasi trans- 3 -dodecanoil- 6 3 C V.1.3 gsartor L04 - I grassi Acidi grassi polinsaturi 3 C C 4 Linoeil- (C ) cis- 4 -decanoil- cis- 3 -cis- 3 -dodecanoil- 3 x β-ossidazione 3 C 4 β-ossidazione 3 3 C 2 Enoil- idratasi 3 C 4 Enoil- isomerasi trans- 3 -cis- 6 -dodecanoil- 3 C 4 trans- 2 -cis- 4 -decanoil- ADP + + ADP + Dienoil- reduttasi 3 C 4 trans- 3 -decanoil- 5 3 C 4 x β-ossidazione Enoil- idratasi 2 V.1.3 gsartor L04 - I grassi
49 Perossisomi La β-ossidazione avviene in molte strutture subcellulari, in particolare nei perossisomi. ono deputati alla formazione di 2 2 che viene utilizzata come sistema di difesa da batteri, virus, ecc. L accettore di elettroni della acil- ossidasi è il FAD: FAD FAD 2 FAD FAD V.1.3 gsartor L04 - I grassi ω-ossidazione el reticolo endoplasmatico può avvenire la ω-ossidazione. n C 3 n n Catalizzata da enzimi che appartengono alla classe delle ossidasi miste (CytP450, CYP) Provocano una idrossilazione e quindi una ossidazione. È un processo aspecifico che converte molecole lipofile in prodotti più idrosolubili più facili da eliminare. È un processo che detossifica le cellule da molecole lipofile. V.1.3 gsartor L04 - I grassi
50 truttura del CYP (1P5) AA basici V.1.3 gsartor L04 - I grassi truttura del CYP (1P5) Cys436 AA basici V.1.3 gsartor L04 - I grassi
51 truttura del CYP (1P5) Cys436 AA basici Cavità V.1.3 gsartor L04 - I grassi Ciclo del CYP (prodotto) R R (substrato) Fe 3+ 2 Fe 3+ R 2 2 Fe 3+ R e - Fe 2+ R Fe 3+ R 2 AD(P)-citocromo P450 reduttasi [Fe 2+ 2 R] e- + [Fe2+ R] -1 [Fe 2+ 2 R] -2 citocromo b5 V.1.3 gsartor L04 - I grassi
52 Meccanismo ciclico del CYP R 2 R R 2 R Fe 3+ Fe 3+ Fe 3+ II I VII 2- Fe 4+ R +. A e VI 2 + R - Fe 3+ + R Fe 2+ III + R 2- R - R 2 Fe 3+ e - Fe 3+ Fe 2+ V IV Fe 2+ V.1.3 gsartor L04 - I grassi Meccanismo ciclico del CYP R 2 I. P450 acquo Fe 3+ (basso spin) Fe 3+ II. Lega R P450 canfora Fe 3+ (alto spin) entra 1e -, riduzione a Fe R III. P450 canfora Fe 2+ VII Lega 2 IV. P450 con 2 legato, Fe 4+ 2 equivalente a Fe R 2 - entra 1e -, riduzione a 2-2 V. P450 perossido VI Fe R A + Fe 3+ I 2 2 R 2 R Fe 3+ e - R Fe 2+ II III + R 2- R - R 2 Fe 3+ e - Fe 3+ Fe 2+ V IV Fe 2+ V.1.3 gsartor L04 - I grassi
53 Meccanismo ciclico del CYP R 2 R R 2 R VI. Entra 1 + P450 idroperossido Entra 1 + esce 2 VII. P450 Fe catione radicale sulla proteina si forma R VII VI 2 + Fe Fe 4+ Fe 3+ - R R +. A + Fe 3+ I 2 2 Fe 3+ e - R Fe 2+ II III A. L idroperossido VI si può formare per reazione di II con Fe R e - R - Fe 3+ R Fe 2+ 2 V IV Fe 2+ V.1.3 gsartor L04 - I grassi Meccanismo L ossigeno è legato non ad angolo retto. Il legame dell ossigeno allontana il ligando (R) solo dopo che I due atomi di ossigeno si sono ridotti il ligando si riavvicina. Ciò previene la formazione di R. Gli elettroni per la riduzione dell ossigeno so forniti da una proteina Fe- (P450 batterica o mitocondriale) o da una ADP-citocromo P450 ossidoreduttasi FAD/FM dipendente (microsomi). V.1.3 gsartor L04 - I grassi
54 Meccanismo generale del CYP 2 ADP + CYTP450 Reduttasi FM 2 /FAD 2 CYP Fe 3+ R ADP + + CYTP450 Reduttasi FM/FAD CYP Fe 2+ R V.1.3 gsartor L04 - I grassi Crediti e autorizzazioni all utilizzo Questo materiale è stato assemblato da informazioni raccolte dai seguenti testi di Biochimica: CAMPE Pamela, ARVEY Richard, FERRIER Denise R. LE BAI DELLA BICIMICA [IB ] Zanichelli EL David L., CX Michael M. I PRICIPI DI BICIMICA DI LEIGER - Zanichelli GARRETT Reginald., GRIAM Charles M. BICIMICA con aspetti molecolari della Biologia cellulare - Zanichelli VET Donald, VET Judith G, PRATT Charlotte W FDAMETI DI BICIMICA [IB ] - Zanichelli E dalla consultazione di svariate risorse in rete, tra le quali: Kegg: Kyoto Encyclopedia of Genes and Genomes Brenda: Protein Data Bank: Rensselaer Polytechnic Institute: Questo ed altro materiale può essere reperito a partire da: oppure da gsartor.org/ Il materiale di questa presentazione è di libero uso per didattica e ricerca e può essere usato senza limitazione, purché venga riconosciuto l autore usando questa frase: Materiale ottenuto dal Prof. Giorgio artor Università di Bologna a Ravenna Giorgio artor - giorgio.sartor@unibo.it 54
Struttura delle proteine
 Struttura delle proteine I II III Copyright 2001-2015 by Giorgio Sartor. All rights reserved. Versione 1.0.2 oct 2015 Struttura quaternaria Èil livello di organizzazione per il quale si formano strutture
Struttura delle proteine I II III Copyright 2001-2015 by Giorgio Sartor. All rights reserved. Versione 1.0.2 oct 2015 Struttura quaternaria Èil livello di organizzazione per il quale si formano strutture
Βeta ossidazione degli acidi grassi BETA OSSIDAZIONE DEGLI ACIDI GRASSI 13/12/2013 SEMINARIO. Acidi grassi (2)
") Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Attenzione : lunedì 29 aprile NON ci sarà lezione
 Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI
 LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
Si possono suddividere in:
 Si possono suddividere in: LIPIDI DI DEPOSITO: si accumulano nelle cellule del tessuto adiposo, sono una riserva energetica, mantengono costante la temperatura del corpo (termogenesi) LIPIDI STRUTTURALI:
Si possono suddividere in: LIPIDI DI DEPOSITO: si accumulano nelle cellule del tessuto adiposo, sono una riserva energetica, mantengono costante la temperatura del corpo (termogenesi) LIPIDI STRUTTURALI:
Prof. Giorgio Sartor. Invertebrati marini. I nutrienti. Fonti di carbonio ed energia. Fonte di carbonio. Fonte di energia. Composti organici.
 Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
SISTEMI ENERGETICI. L ATP privato di uno dei suoi 3 radicali fosforici diventa ADP (adenosindifosfato).
.") SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
GLUCONEOGENESI. Sintesi (GENESI) di nuove (NEO) molecole di glucosio
 di nuove (NEO) molecole di glucosio") GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 31 Lipidi I lipidi si dividono in due gruppi principali: I gruppo: comprende composti a catena aperta con una testa polare
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 31 Lipidi I lipidi si dividono in due gruppi principali: I gruppo: comprende composti a catena aperta con una testa polare
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Metabolismo dei grassi
 Prof. Giorgio Sartor Metabolismo dei grassi Copyright 2001-2019 by Giorgio Sartor. All rights reserved. B12 - Versione 2.0 Mar-19 1 Lipidi Semplici Sono molecole che non contengono legami esterei o amidici
Prof. Giorgio Sartor Metabolismo dei grassi Copyright 2001-2019 by Giorgio Sartor. All rights reserved. B12 - Versione 2.0 Mar-19 1 Lipidi Semplici Sono molecole che non contengono legami esterei o amidici
«Macromolecole» Lipidi
 LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
CELLULE EUCARIOTICHE
 CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
METABOLISMO LIPIDICO. Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva. Lipidi introdotti con gli alimenti.
 METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
METABOLISMO DEI LIPIDI
 METABOLISMO DEI LIPIDI Generalità I lipidi alimentari, per poter essere assorbiti dall organismo umano, devono essere preventivamente digeriti (ad eccezione del colesterolo), cioè, trasformati in molecole
METABOLISMO DEI LIPIDI Generalità I lipidi alimentari, per poter essere assorbiti dall organismo umano, devono essere preventivamente digeriti (ad eccezione del colesterolo), cioè, trasformati in molecole
Da dove prendono energia le cellule animali?
 Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
dotto di Wirsung che si riversa nella papilla maggiore del duodeno;
 Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Metabolismo dei grassi
 Prof. Giorgio artor Metabolismo dei grassi Copyright 2001-2011 by Giorgio artor. All rights reserved. B12 - Versione 1.5 may 2011 Lipidi emplici ono molecole che non contengono legami esterei o amidici
Prof. Giorgio artor Metabolismo dei grassi Copyright 2001-2011 by Giorgio artor. All rights reserved. B12 - Versione 1.5 may 2011 Lipidi emplici ono molecole che non contengono legami esterei o amidici
METABOLISMO E SISTEMI ENERGETICI
 METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI Nelson Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER,
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI Nelson Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER,
Metabolismo degli aminoacidi
 Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di
 IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico 2014-15
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
INSOLUBILI IN ACQUA E SOLUBILI NEI SOLVENTI ORGANICI NON POLARI
 LIPIDI Una delle quattro principali classi di sostanze biologicamente attive. I lipidi (la parola deriva dal greco lípos, grasso) sono costituenti delle piante e degli animali, caratterizzati da particolari
LIPIDI Una delle quattro principali classi di sostanze biologicamente attive. I lipidi (la parola deriva dal greco lípos, grasso) sono costituenti delle piante e degli animali, caratterizzati da particolari
Biosintesi dei triacilgliceroli
 Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE. L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO
 GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
glicogeno Glucosio 6-P Piruvato Acetil CoA Intermedi del ciclo di Krebs
 PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
digestione dei lipidi alimentari duodeno digiuno sali biliari fosfolipidi bile specifiche idrolasi pancreatiche colecistochinina
 La digestione dei lipidi alimentari, in prevalenza trigliceridi, avviene nel duodeno e nel digiuno per azione combinata dei sali biliari e dei fosfolipidi della bile e delle specifiche idrolasi pancreatiche
La digestione dei lipidi alimentari, in prevalenza trigliceridi, avviene nel duodeno e nel digiuno per azione combinata dei sali biliari e dei fosfolipidi della bile e delle specifiche idrolasi pancreatiche
Il ciclo dell acido citrico
 Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
GLICOGENOLISI & GLICOGENOSINTESI
 GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
Modulo 15 metabolismo dei lipidi
 Modulo 15 metabolismo dei lipidi Il metabolismo cellulare dei lipidi (AG= acido grasso) AGE= AG essenziale 7 Digestione ed assorbimento e utilizzazione dei lipidi 90% dieta triacilgliceroli 10% colesterolo,
Modulo 15 metabolismo dei lipidi Il metabolismo cellulare dei lipidi (AG= acido grasso) AGE= AG essenziale 7 Digestione ed assorbimento e utilizzazione dei lipidi 90% dieta triacilgliceroli 10% colesterolo,
I LIPIDI. - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua
 I LIPIDI I LIPIDI - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua - ruolo fondamentale di fornitori di energia (circa 9 kcal/grammo),
I LIPIDI I LIPIDI - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua - ruolo fondamentale di fornitori di energia (circa 9 kcal/grammo),
N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi
 Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
ENZIMI CINETICA ENZIMATICA
 ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
Nota dell editore Presentazione
 00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
dieta vengono convertiti in composti dei corpi chetonici.
 Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
METABOLISMO OSSIDATIVO
 METABOLISMO OSSIDATIVO IDROLISI DEI LIPIDI LIPASI Fosfolipidi Fosfolipasi β ossidazione degli acidi grassi NEL CITOPLASMA: attivazione dell acido grasso Consumo di 2 ATP ΔG idrolisi ATP = -35,9 kj mol
METABOLISMO OSSIDATIVO IDROLISI DEI LIPIDI LIPASI Fosfolipidi Fosfolipasi β ossidazione degli acidi grassi NEL CITOPLASMA: attivazione dell acido grasso Consumo di 2 ATP ΔG idrolisi ATP = -35,9 kj mol
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
unità C2. Le trasformazioni energetiche nelle cellule
 unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
METABOLISMO DEI GRASSI
 Capitolo 27 METABOLISMO DEI GRASSI La maggior parte dell energia conservata in un organismo si trova nei depositi di grasso. In questi corpulenti combattenti di sumo la conservazione di energia sotto forma
Capitolo 27 METABOLISMO DEI GRASSI La maggior parte dell energia conservata in un organismo si trova nei depositi di grasso. In questi corpulenti combattenti di sumo la conservazione di energia sotto forma
MOVIMENTO. Motori molecolari. IV Movimento Miosina e actina. Prof. Giorgio Sartor. P04 - Versione 2.0 oct 2013
 Prof. Giorgio Sartor Motori molecolari IV Movimento Miosina e actina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P04 - Versione 2.0 oct 2013 MOVIMENTO Miosina Actina P04 v. 2.0 gsartor
Prof. Giorgio Sartor Motori molecolari IV Movimento Miosina e actina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P04 - Versione 2.0 oct 2013 MOVIMENTO Miosina Actina P04 v. 2.0 gsartor
Catena di Trasporto degli elettroni
 Chimica Biologica A.A. 2010-2011 Catena di Trasporto degli elettroni Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Respirazione - il catabolismo di tutti i combustibili
Chimica Biologica A.A. 2010-2011 Catena di Trasporto degli elettroni Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Respirazione - il catabolismo di tutti i combustibili
Cosa sono i grassi o lipidi?
 Cosa sono i grassi o lipidi? Lipidi da LIPOS = grasso Sostanze organiche di diversa natura con una caratteristica comune: sono insolubili in acqua Importanti costituenti dei tessuti vegetali e animali
Cosa sono i grassi o lipidi? Lipidi da LIPOS = grasso Sostanze organiche di diversa natura con una caratteristica comune: sono insolubili in acqua Importanti costituenti dei tessuti vegetali e animali
http://sds.coniliguria.it 1
 Aspetti metabolici dell esercizio fisico Genova 22 gennaio 2011 A cura di Attilio TRAVERSO http://sds.coniliguria.it 1 L allenamento produce modificazioni fisiologiche (adattamenti) in quasi tutti i sistemi
Aspetti metabolici dell esercizio fisico Genova 22 gennaio 2011 A cura di Attilio TRAVERSO http://sds.coniliguria.it 1 L allenamento produce modificazioni fisiologiche (adattamenti) in quasi tutti i sistemi
Metabolismo dei lipidi
 Metabolismo dei lipidi 1 Funzione centrale dei lipidi nel metabolismo energetico 1)Assorbimento e trasporto degli acidi grassi 1) Ossidazione degli acidi grassi 2) sintesi degli acidi grassi e del colesterolo
Metabolismo dei lipidi 1 Funzione centrale dei lipidi nel metabolismo energetico 1)Assorbimento e trasporto degli acidi grassi 1) Ossidazione degli acidi grassi 2) sintesi degli acidi grassi e del colesterolo
Il catabolismo dei lipidi e l ossidazione degli acidi grassi
 Il catabolismo dei lipidi e l ossidazione degli acidi grassi Nella maggior parte degli organismi i lipidi si trovano principalmente nella forma di TRIACILGLICEROLI (trigliceridi) In molte piante una grossa
Il catabolismo dei lipidi e l ossidazione degli acidi grassi Nella maggior parte degli organismi i lipidi si trovano principalmente nella forma di TRIACILGLICEROLI (trigliceridi) In molte piante una grossa
CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) scaricato da www.sunhope.it
 FOSFORILAZIONE OSSIDATIVA (FO) scaricato da www.sunhope.it") CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) - Decarb. ossid. piruvato - β-ossidazione acidi grassi - Vie ossidative aa - Ciclo di Krebs avvengono tutte nella matrice mitocondriale In tutte
CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) - Decarb. ossid. piruvato - β-ossidazione acidi grassi - Vie ossidative aa - Ciclo di Krebs avvengono tutte nella matrice mitocondriale In tutte
Niccolò Taddei Biochimica
 Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
BIOLOGIA GENERALE 22-24 ottobre 2007
 Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
LIPIDI COMPLESSI E LIPOPROTEINE
 LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
FUNZIONI DEI MITOCONDRI
 FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
Mais, riso, patate, granozucchero di canna o barbabietola Latte da zucchero Polisaccaride Amido - - Disaccaride Maltosio Saccarosio Lattosio
 Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara
 Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici
 I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
METABOLISMO DEL Glucosio
 METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
Prof. Maria Nicola GADALETA
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Idratazione (introduciamo un gruppo OH sul Cβ) per mezzo della enoil-coa idratasi
 per mezzo della enoil-coa idratasi") β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
Il catabolismo degli acidi grassi
 Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Fosforilazione ossidativa
 Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine
 Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
4) PROTEINE FERRO ZOLFO. Contengono Fe e S in quantità equimolecolari; fanno parte dei complessi respiratori. La NADH DH contiene diversi centri Fe-S.
 PROTEINE FERRO ZOLFO. Contengono Fe e S in quantità equimolecolari; fanno parte dei complessi respiratori. La NADH DH contiene diversi centri Fe-S.") 4) PROTEINE FERRO ZOLFO Contengono Fe e S in quantità equimolecolari; fanno parte dei complessi respiratori. La NADH DH contiene diversi centri Fe-S. Trasportano elettroni attraverso le transizioni Fe
4) PROTEINE FERRO ZOLFO Contengono Fe e S in quantità equimolecolari; fanno parte dei complessi respiratori. La NADH DH contiene diversi centri Fe-S. Trasportano elettroni attraverso le transizioni Fe
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
ANABOLISMO DEI LIPIDI
 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
Degradazione degli acidi grassi
 Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
I LIPIDI. Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi
 Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
I L I P I D I. Lipidi complessi: fosfolipidi e glicolipidi; sono formati da CHO e altre sostanze.
 I L I P I D I ASPETTI GENERALI I Lipidi o grassi sono la riserva energetica del nostro organismo; nel corpo umano costituiscono mediamente il 17% del peso corporeo dove formano il tessuto adiposo. In generale
I L I P I D I ASPETTI GENERALI I Lipidi o grassi sono la riserva energetica del nostro organismo; nel corpo umano costituiscono mediamente il 17% del peso corporeo dove formano il tessuto adiposo. In generale
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato
 2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
Prima di essere ossidati. gli ACIDI GRASSI devono essere attivati. (cioè legati al CoA) e poi entrare nel mitocondrio
 e poi entrare nel mitocondrio") Prima di essere ossidati gli ACIDI GRASSI devono essere attivati (cioè legati al CoA) e poi entrare nel mitocondrio L enzima acil CoA sintetasi (= acido grasso tiokinasi) sta sulla membrana mitocondriale
Prima di essere ossidati gli ACIDI GRASSI devono essere attivati (cioè legati al CoA) e poi entrare nel mitocondrio L enzima acil CoA sintetasi (= acido grasso tiokinasi) sta sulla membrana mitocondriale
Metabolismo degli amminoacidi. Ciclo dell urea
 Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di
 Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di materiali per fare funzionare il nostro corpo È il carboidrato
Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di materiali per fare funzionare il nostro corpo È il carboidrato
Biosintesi non ribosomiale di metaboliti peptidici bioattivi
 Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
ANTIOSSIDANTI Grassi IV LEZIONE
 ANTIOSSIDANTI Grassi IV LEZIONE ANTIOSSIDANTE Sostanza che, pur presente in concentrazione ridotta rispetto al substrato ossidabile, può prevenire o ritardare l ossidazione iniziata da un fattore proossidante.
ANTIOSSIDANTI Grassi IV LEZIONE ANTIOSSIDANTE Sostanza che, pur presente in concentrazione ridotta rispetto al substrato ossidabile, può prevenire o ritardare l ossidazione iniziata da un fattore proossidante.
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido
 Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
Continua. Peptidasi H 2 O
 Continua Peptidasi H 2 O Classificazione delle peptidasi 1. Meccanismo catalitico 2. Tipo di reazione catalizzata 3. Struttura molecolare e omologia 1. Meccanismo catalitico (mostrato per la chimotripsina)
Continua Peptidasi H 2 O Classificazione delle peptidasi 1. Meccanismo catalitico 2. Tipo di reazione catalizzata 3. Struttura molecolare e omologia 1. Meccanismo catalitico (mostrato per la chimotripsina)
scaricato da www.sunhope.it
 Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Metabolismo degli amminoacidi
 Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
Regolazione del metabolismo del glucosio
 Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
Fasi dell ossidazione degli acidi grassi
 Fasi dell ossidazione degli acidi grassi REGOLAZIONE DELLA OSSIDAZIONE DEGLI ACIDI GRASSI La velocità di trasporto nella matrice mitocondriale decide il destino (ossidazione) CARNITINA ACIL-TRANSFERASI
Fasi dell ossidazione degli acidi grassi REGOLAZIONE DELLA OSSIDAZIONE DEGLI ACIDI GRASSI La velocità di trasporto nella matrice mitocondriale decide il destino (ossidazione) CARNITINA ACIL-TRANSFERASI
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Ruolo metabolico delle flavine
 Ruolo metabolico delle flavine Deidrogenasi flaviniche che trasferiscono 2 elettroni Acil CoA deidrogenasi (beta ossidazione acidi grassi) Diidrolipoildeidrogenasi (complessi piruvato dh etc.) Glutatione
Ruolo metabolico delle flavine Deidrogenasi flaviniche che trasferiscono 2 elettroni Acil CoA deidrogenasi (beta ossidazione acidi grassi) Diidrolipoildeidrogenasi (complessi piruvato dh etc.) Glutatione
La fotosintesi: energia dal Sole
 La fotosintesi: energia dal Sole Gli organismi fotosintetici usano la luce del Sole, l acqua del suolo e il CO 2 dell atmosfera per produrre composti organici e liberare O 2 grazie alla fotosintesi. Sadava
La fotosintesi: energia dal Sole Gli organismi fotosintetici usano la luce del Sole, l acqua del suolo e il CO 2 dell atmosfera per produrre composti organici e liberare O 2 grazie alla fotosintesi. Sadava
Il Metabolismo Energetico 1. Bilancio Quantitativo tra Apporto di Energia e Dispendio Energetico
 Il Metabolismo Energetico 1. Bilancio Quantitativo tra Apporto di Energia e Dispendio Energetico Prof. Carlo Capelli Fisiologia Facoltà di Scienze Motorie, Università degli Studi Verona Obiettivi Energia
Il Metabolismo Energetico 1. Bilancio Quantitativo tra Apporto di Energia e Dispendio Energetico Prof. Carlo Capelli Fisiologia Facoltà di Scienze Motorie, Università degli Studi Verona Obiettivi Energia
la struttura tridimensionale può essere ottenuta solo per Un intero dominio in genere da 50 a 300 residui
 Durante la traduzione l informazione di ripiegamento codificata nella sequenza aminoacidica diventa disponibile in maniera vettoriale la struttura tridimensionale può essere ottenuta solo per Un intero
Durante la traduzione l informazione di ripiegamento codificata nella sequenza aminoacidica diventa disponibile in maniera vettoriale la struttura tridimensionale può essere ottenuta solo per Un intero
Valitutti, Taddei, Kreuzer, Massey, Sadava, Hills, Heller, Berenbaum
 Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
ghiandole salivari bocca amilasi stomaco HCl (ph 1) pepsinogeno pepsina
 pepsinogeno pepsina") Metabolismo lipidico Digestione I lipidi della dieta sono Trigliceridi Fosfolipidi Colesterolo processi digestivi ghiandole salivari amilasi bocca HCl (ph 1) pepsinogeno pepsina stomaco processi digestivi
Metabolismo lipidico Digestione I lipidi della dieta sono Trigliceridi Fosfolipidi Colesterolo processi digestivi ghiandole salivari amilasi bocca HCl (ph 1) pepsinogeno pepsina stomaco processi digestivi
>> Il malonil-coa è prodotto nel citosol a partire dall acetil-coa.
 SINTESI DEGLI ACIDI GRASSI >> AVVIENE NEL CITOSOL. Nei mammiferi principalmente negli epatociti e negli adipociti. >> per sintetizzare acidi grassi è necessario avere nel citosol: Acetil-CoA, Malonil-CoA
SINTESI DEGLI ACIDI GRASSI >> AVVIENE NEL CITOSOL. Nei mammiferi principalmente negli epatociti e negli adipociti. >> per sintetizzare acidi grassi è necessario avere nel citosol: Acetil-CoA, Malonil-CoA
Segnalazione cellulare e trasduzione del segnale. Comunicazione fra le cellule
 Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce
Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce