Struttura secondaria, Motivi e Domini nelle Proteine
|
|
|
- Evaristo Franco
- 6 anni fa
- Visualizzazioni
Transcript
1 Struttura secondaria, Motivi e Domini nelle Proteine

2 Proprietà generali Le forme ioniche degli aminoacidi, senza considerare alcuna ionizzazione delle catene laterali.
3 Proprietà generali Tutti gli aminoacidi (ad eccezione della glicina) derivati da proteine sono otticamente attivi, cioè ruotano il piano di polarizzazione della luce. Le molecole otticamente attive sono asimmetriche in modo da non essere sovrapponibili con le loro immagini speculari (enantiomeri). Ciò è caratteristico di molecole contenenti atomi di carbonio tetraedrici con 4 diversi sostituenti. L atomo centrale si dice centro chirale.
4 Il legame peptidico Il gruppo peptidico ha una struttura rigida e planare, dovuta al parziale (~40 %) carattere di doppio legame del legame peptidico. O O - C C N N + H H


5 Il legame peptidico Il legame peptidico C N è 0.13 Å più corto del legame singolo N C e0.08å più lungo di un doppio legame C=N. Il legame peptidico, quindi, presenta per il 60 % una natura di legame singolo e per il 40 % una natura di legame doppio.
 n e(c ) n+1 opposti legame peptidico C N.")

6 Il legame peptidico Generalmente il gruppo peptidico assume la conformazione trans trans -atomi (C ) n e(c ) n+1 opposti legame peptidico C N. rispetto al In alcuni casi il gruppo peptidico può assume la conformazione cis (~8 kj mol -1 meno stabile della conformazione trans. Problemi sterici, perché distanza C -C =2.8Å). cis
7 Il legame peptidico Il gruppo peptidico ha un momento di dipolo. Gli atomi O e N sono rispettivamente più elettronegativi di C ed H. La conseguente delocalizzazione di carica porta alla formazione di due dipoli (CO ed NH) con analoga direzione e verso nel gruppo peptidico. Il momento di dipolo risultante è di circa 3.5 Debye. O C H N
8 Gli aminoacidi Gli aminoacidi sono generalmente classificati a seconda della polarità delle loro catene laterali. Infatti, il ripiegamento della catena polipeptidica nella sua conformazione nativa è dovuto principalmente alla tendenza che hanno le catene laterali idrofobiche a sfuggire il contatto con il solvente e le catene laterali idrofiliche ad essere esposte all acqua. Si possono quindi distinguere 3 gruppi principali di aminoacidi: - gruppo Rnonpolare - gruppo R polare non carico - gruppo R polare carico
9 Gruppo R non polare Gruppo R polare non carico Glicina Gly G Serina Ser S Alanina Ala A TreoninaThr T Valina Val V Asparagina Asn N Leucina Leu L Glutammina Gln Q Isoleucina Ile I Tirosina Tyr Y Metionina Met M Cisteina Cys C Prolina Pro P Fenilalanina Phe F Triptofano Trp W Gruppo R polare carico Lisina Lys K Arginina Arg R Istidina His H Acido Aspartico Asp D Acido Glutammico Glu E
10 Proprietà acido-base pk a degli aminoacidi I valori di pk a del gruppo carbossilico ed aminico degli altri aminoacidi sono simili a quelli della glicina. Alcuni aminoacidi contengono anche catene laterali ionizzabili: Asp, Glu, His, Cys, Tyr, Lys e Arg. Il pk a delle catene laterali ionizzabili degli aminoacidi è compreso tra i valori 3.9 (Asp) e 12.5 (Arg).
 - rotazione intorno al legame C C (angolo ) A ciascun aminoacido viene associata una")
11 Angoli diedri e Proprietà fondamentale del legame peptidico è di essere rigido e planare. Isoligradidilibertàdell unità peptidica rigida sono: - rotazione intorno al legame N C (angolo ) - rotazione intorno al legame C C (angolo ) A ciascun aminoacido viene associata una coppia di angoli diedri (, ), che determina in modo univoco la conformazione della catena principale.
 stericamente permessi si possono determinare calcolando le distanze fra gli atomi di un tripeptide in corrispondenza di tutti i valori (, ) per l unità peptidica centrale.")
12 Angoli diedri e Molte combinazioni di angoli (, ) per gli aminoacidi non sono permesse, a causa delle collisioni steriche fra atomi della catena principale e/o delle catene laterali. I valori di (, ) stericamente permessi si possono determinare calcolando le distanze fra gli atomi di un tripeptide in corrispondenza di tutti i valori (, ) per l unità peptidica centrale. Le conformazioni stericamente proibite sono quelle per cui la distanza interatomica di un interazione non covalente è inferiore alla corrispondente somma di raggi di van der Waals.
13 Plot di Ramachandran Le coppie ( ) permesse per gli aminoacidi sono riportate in un diagramma chiamato plot di Ramachandran, dal nome del fisico indiano G.N. Ramachandran che per primo (fine anni 60) calcolò le regioni stericamente permesse. Ramachandran e colleghi usarono il modello che schematizza gli atomi come sfere rigide e fissa le geometrie dei legami. Misure più precise del plot di Ramachandran risultano dal calcolo delle energie relative a ciascuna conformazione, permettendo una certa flessibilità per i contatti di van der Waals e per gli angoli di legame, e calcolando i contributi di tutte le interazioni favorevoli e non.

14 Plot di Ramachandran Plot di Ramachandran relativo ad un tripeptide costituito da Ala. Qualunque scelta accettabile si faccia del raggio di van der Waals, si ottengono comunque solo 3 regioni del plot di Ramachandran fisicamente accessibili alla catena polipeptidica. Le zone in azzurro sono quelle largamente permesse, mentre le zone in verde indicano le conformazioni, comunque possibili, che hanno distanze di van der Waals al limite inferiore dell accettabilità.

15 Coot Plot di Ramachandran Un caso a parte è rappresentato dalla glicina: le zone permesse occupano fino al 45% del plot di Ramachandran. Il plot di Ramachandran per la glicina è centrosimmetrico, riflettendo il fatto che la glicina è l unico aminoacido a non essere asimmetrico. La glicina gioca un ruolo strutturale molto importante, permettendo conformazioni inusuali alla catena principale delle proteine. Le zone in azzurro sono quelle largamente permesse, mentre le zone in verde indicano le conformazioni, comunque possibili, che hanno distanze di van der Waals al limite inferiore dell accettabilità.
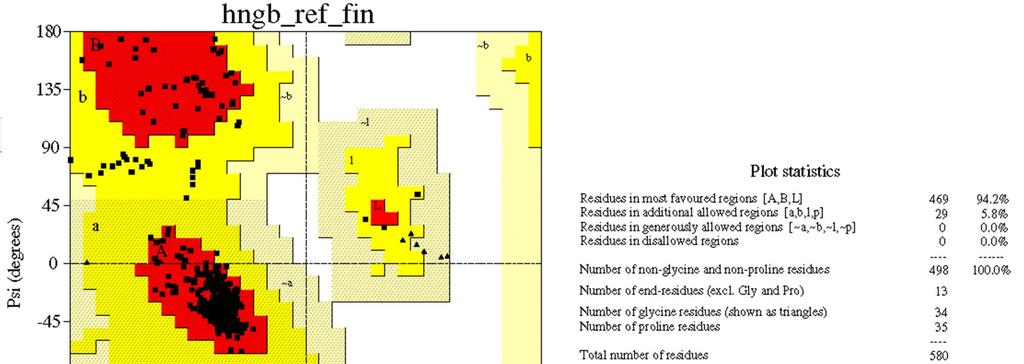
16 Plot di Ramachandran
17 Plot di Ramachandran Le regioni del plot di Ramachandran sono indicate con il nome della conformazione risultante in un peptide se i corrispondenti angoli ( ) sono ripetuti negli aminoacidi successivi lungo la catena polipeptidica. -eliche destrorse filamenti antiparalleli e paralleli -eliche sinistrorse
18 Plot di Ramachandran Valori di (, ) relativi a ciascuna struttura secondaria
19 La struttura secondaria Le strutture secondarie sono disposizioni regolari della catena polipeptidica principale, che vengono classificate senza fare riferimento al tipo di aminoacidi. Esse sono stabilizzate da legami idrogeno fra il gruppo aminico e il gruppo carbonilico della catena principale. La regolarità della conformazione risulta dalla regolarità della struttura atomica della catena polipeptidica, ed è evidenziata dai valori degli angoli diedri (, ) di ciascun aminoacido, che si ripetono quasi costantemente all interno di ogni elemento di struttura secondaria.
20 La struttura secondaria Si distinguono fondamentalmente tre tipi di strutture secondarie: elica foglietto reverse turn Circa il % degli aminoacidi delle proteine globulari assume la conformazione regolare tipica di uno dei tre tipi di struttura secondaria. I segmenti di catena polipeptidica che non sono in elica, foglietto o turn, assumono la conformazione chiamata loop o random coil, struttura non ripetitiva né regolare, spesso priva di legami idrogeno tra gli aminoacidi che la compongono.


21 (D.L. Nelson, M.M. Cox, Lehninger Principles of Biochemistry, 4th ed., W.H. Freeman & Co., 2005)
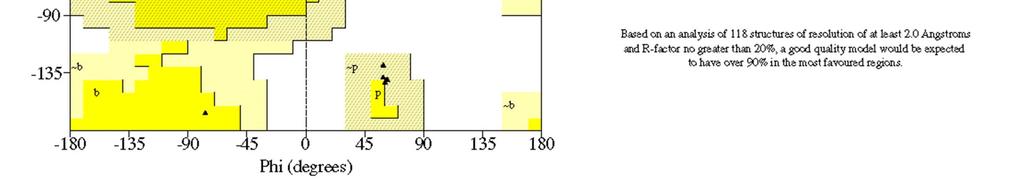
22 elica L elica è la struttura secondaria più frequentemente adottata dalla catena polipeptidica delle proteine: circa il % degli aminoacidi delle proteine globulari di struttura tridimensionale nota assume la conformazione ad elica. Fu descritta per la prima volta nel 1951 da Linus Pauling e Robert Corey (California Institute of Technology), che ipotizzarono una struttura stabile ed energeticamente favorita nelle proteine, sulla base di parametri geometrici accurati per l unità peptidica dedotti dall analisi cristallografica di strutture di piccole molecole. La previsione di Pauling e Corey ricevette molto presto il supporto sperimentale dalla determinazione della struttura tridimensionale della mioglobina (John Kendrew, 1960) e dell emoglobina (Max Perutz, 1961), due proteine i cui elementi di struttura secondaria sono eliche.
, molto rare e limitate a pochi aminoacidi.")
23 elica Un elica destrorsa è caratterizzata da: (, ) = (-57, -47 ), corrispondenti al quadrante in basso a sinistra del plot di Ramachandran; n = 3.6 residui per giro p =5.4Å d =1.5Å Esistono anche eliche di tipo sinistrorso (quadrante in alto a destra del plot di Ramachandran), molto rare e limitate a pochi aminoacidi. Esse sono caratterizzate da valori (, ) = (+57, +47 ) e da stessi valori per n e p (n negativo).
24 (D. Voet, J.G. Voet, Biochemistry, 3 ed., John Wiley & Sons, 2004)
25 elica Le catene laterali degli aminoacidi che appartengono ad un elica destrorsa sono rivolte verso l esterno e verso l estremità N-terminale dell elica. In questo modo si evitano interferenze steriche con la catena polipeptidica principale e tra le catene laterali stesse. Un elica sinistrorsa si trova molto raramente nelle proteine (solo brevi tratti di 3-5 aminoacidi) perché le catene laterali interferirebbero stericamente con la catena principale.
26 elica All elica è associato un macrodipolo. Ad ogni unità peptidica è associato un dipolo; nell elica questi dipoli sono tutti allineati lungo l asse dell elica. I dipoli associati a ciascuna unità peptidica si sommano e un elica costituita da n aminoacidi avrà un momento di dipolo complessivo pari a n 3.5 Debye.

27 (D. Voet, J.G. Voet, Biochemistry, 3 ed., John Wiley & Sons, 2004)
28 Altri tipi di elica Variazioni dell elica, in cui la catena polipeptidica è avvolta più o meno strettamente, con legami idrogeno fra coppie di aminoacidi (n n+3) o (n n+5), sono chiamate elica 3 10 ed elica. Entrambe queste conformazioni si trovano ai limiti della regione permessa per le strutture elicoidali destrorse nel plot di Ramachandran: Elica 3 10 :( ) = (-49, -26 ) Elica : ( ) = (-57, -70 ) Elica 3 10 ed elica ricorrono poco frequentemente nelle proteine. In particolare, le eliche 3 10 possono essere presenti alle estremità delle eliche, come segmenti molto corti (3-4 aminoacidi).

29 Altri tipi di elica

30 Elica 3 10 L elica 3 10 deve il suo nome al numero di aminoacidi per giro (3) e al numero di atomi (10) compresi fra un donatore ed un accettore di legame idrogeno. Secondo questa nomenclatura, l elica sarebbe un elica Le eliche 3 10 sono meno favorite delle eliche: - gli atomi della catena principale sono troppo strettamente impaccati, portando ad interazioni di van der Waals repulsive; - i legami idrogeno non sono lineari; - i dipoli delle unità peptidiche deviano di circa 30 rispetto all asse dell elica; - la posizione delle catene laterali (allineate) porta ad interferenze steriche.


31 Page 233 Coiled-coil di 2 -eliche


32 Page 232 Struttura schematica dell -cheratina
33 Coiled-coil elica: actina e miosina Kinemage Leu_Zipper La molecola di miosina è un dimero costituito da due catene pesanti (MW 230 kda) e quattro catene leggere (MW 20 kda) e forma una coda lunga 1400 Å e due teste. Frammenti di miosina, costituiti da due catene leggere e la parte N-terminale di una delle catene pesanti, si possono separare e costituiscono il sottoframmento S1.

34 Foglietto Nello stesso anno (1951) in cui proposero l elica, Pauling e Corey postularono anche l esistenza di un altra struttura secondaria: il foglietto ( -sheet). Dopo l elica, la conformazione più ricorrente adottata dalla catena polipeptidica delle proteine è il foglietto circa il % degli aminoacidi nelle proteine globulari conosciute ricorre in questa conformazione. Gli angoli diedri ( ) della catena polipeptidica in conformazione foglietto cadono nella zona permessa del plot di Ramachandran (quadrante in alto a sinistra).
35 Filamento L unità costituente i foglietti èilfilamento ( strand), lungo in media da 5 a 10 aminoacidi, con la catena polipeptidica quasi completamente estesa. I foglietti nelle proteine globulari sono costituiti da 2 a 15 filamenti abbinati lateralmente, con un valor medio di 6. Il filamento si può considerare un tipo speciale di elica, con n = 2 residui per giro e una traslazione d = 3.4 Å per aminoacido. I filamenti sono allineati uno vicino all altro, in modo tale che si possano formare legami idrogeno traigruppicodiunfilamentoeigruppinhdel filamento adiacente e viceversa.

36 Foglietto I foglietti formati da un certo numero di filamenti sono pieghettati (pleated), con gli atomi C degli aminoacidi adiacenti alternativamente leggermente sopra e sotto il piano del foglietto dando al foglietto l apparenza di un foglietto pieghettato. Le catene laterali degli aminoacidi che compongono il foglietto seguono lo stesso andamento, per cui puntano alternativamente sopra e sotto il foglietto. Spesso un foglietto può presentare tutte le catene laterali polari su una faccia e tutte le catene laterali non polari sull altra (ideale per costruire la superficie delle proteine).
 I filamenti possono interagire fra loro a formare i foglietti in due modi diversi: -")
 e si parla di foglietto antiparallelo; - gli aminoacidi nei filamenti")
37 Foglietto (v. Animated_Lesson 8_1c) I filamenti possono interagire fra loro a formare i foglietti in due modi diversi: - gli aminoacidi nei filamenti adiacenti hanno direzioni alternate (N-term C-term seguito da C-term N-term seguito da N-term C-term, ) e si parla di foglietto antiparallelo; - gli aminoacidi nei filamenti allineati vanno tutti nella stessa direzione (sempre N-term C-term) e si parla di foglietto parallelo.
38 Foglietto antiparallelo Angoli diedri ( ) = (-139, 135 ). Coppie di legami idrogeno poco spaziate si alternano a coppie largamente spaziate. Tali legami idrogeno sono paralleli fra loro e perpendicolari alla direzione dei filamenti. I dipoli associati alle unità peptidiche costituenti un filamento si annullano.
39 Foglietto parallelo Angoli diedri ( ) = (-119, 113 ). Coppie di legami idrogeno uniformemente spaziati, non sono paralleli fra loro né perpendicolari alla direzione dei filamenti. I dipoli associati alle unità peptidiche costituenti un filamento si annullano. I foglietti paralleli sono meno frequenti di quelli antiparalleli.
40 Foglietto misto I filamenti si possono anchecombinareaformare un foglietto misto, con alcune coppie di filamenti antiparalleli ed altre paralleli. I foglietti misti sono meno frequenti dei foglietti antiparalleli e paralleli.
41 Reverse turn Le proteine globulari hanno una forma compatta, dovuta a numerose inversioni della direzione della catena polipeptidica che le compone. Molte di queste inversioni sono dovute alla presenza di un comune elemento strutturale, chiamato reverse turn. Gli aminoacidi coinvolti nella formazione dei reverse turn si trovano generalmente sulla superficie delle proteine e hanno natura polare e/o carica. Esistono vari tipi di reverse turn, a seconda del numero di aminoacidi che li costituiscono e degli elementi di struttura secondaria che collegano: -turn -turn -loop
42 -turn Tipo I Angoli diedri ( ) 2 = (-60, -30 ) Angoli diedri ( ) 3 = (-90, 0 ) I -turn di tipo I si possono considerare un breve tratto distorto di elica Nei -turn di tipo I l aminoacido in posizione 2 spesso è Pro, poiché facilmente può assumere la conformazione voluta. Essi sono circa 2-3 volte più frequenti di quelli di tipo II. 2 1


43 Kinemages: Bchl_prot / Porin -turn Tipo II Angoli diedri ( ) 2 = (-60, 120 ) Angoli diedri ( ) 3 = (90, 0 ) I -turnditipoiidifferisconoda quelli di tipo I per un flip di 180 dell unità peptidica che collega gli 2 aminoacidi in posizione 2 e 3. 1 Nei -turn di tipo II l aminoacido in posizione 2 spesso è Pro, poiché facilmente può assumere la conformazione voluta, mentre l aminoacido in posizione 3 spesso è una Gly, per evitare che l atomo di carbonio della catena laterale sia troppo vicino all atomo di ossigeno dell aminoacido in posizione
 caratterizzate dalla ripetizione del motivo (-Gly-Ser-Gly-Ala-Gly-Ala) n.")
44 Fibroina della seta L analisi della sequenza delle fibrine della seta ha mostrato la presenza di un motivo comune: domini variabili alle estremità N- e C-terminali, affiancati da ampie regioni (fino a 800 aminoacidi) caratterizzate dalla ripetizione del motivo (-Gly-Ser-Gly-Ala-Gly-Ala) n. Questi tratti ripetitivi di catena polipeptidica formano foglietti,in cui gli aminoacidi Gly si trovano su un lato del foglietto e Ala/Ser sull altro lato.
45

46 elica
47 Foglietto Anim. Fig. 8-7


48 -turn Tipo I Angoli diedri ( ) 2 = (-60, -30 ) Angoli diedri ( ) 3 = (-90, 0 ) I -turn di tipo I si possono considerare un breve tratto distorto di elica Nei -turn di tipo I l aminoacido in posizione 2 spesso è Pro, poiché facilmente può assumere la conformazione voluta. Essi sono circa 2-3 volte più frequenti di quelli di tipo II. 2 1



49 La struttura supersecondaria
50 La struttura supersecondaria Elementi di struttura secondaria si combinano a costituire aggregati locali con geometria specifica, che definiscono la struttura supersecondaria (o motivi). Alcuni di questi motivi si possono associare ad una particolare funzione, come ad esempio il legame del DNA, mentre altri non hanno una funzione biologica specifica, ma sono semplicemente parte di organizzazioni strutturali più ampie e complesse. I principali motivi individuati nelle proteine sono: - motivi - motivi - motivi /
51 Motivi Nonostante siano le strutture secondarie più frequenti nelle proteine, le eliche isolate non sono stabili in soluzione. Nelle strutture terziarie delle proteine le eliche si impaccano in modo adiacente una all altra attraverso interazioni tra le catene laterali idrofobiche. Iprincipalimotivi sono: - -loop- - EF hand
52 Motivi -loop- Il motivo più semplice consiste di 2 eliche antiparallele collegate da una regione di loop, chiamato hairpin. La più breve connessione fra 2 eliche coinvolge 2aminoacidi, di cui il secondo è sempre Gly, orientati perpendicolarmente agli assi delle eliche. Le eliche risultano cosi antparallele, e sono stabilizzate dall interazione dei loro macrodipoli. N C
 In particolare, una di queste 2 eliche si va ad inserire nel solco maggiore del DNA, e riconosce le basi nucleotidiche, mentre l altra interagisce con i gruppi")
53 Motivi -loop- Un particolare motivo -loop- è caratteristico di alcune proteine che riconoscono e legano specifiche zone di DNA. (noto anche come motivo HTH) In particolare, una di queste 2 eliche si va ad inserire nel solco maggiore del DNA, e riconosce le basi nucleotidiche, mentre l altra interagisce con i gruppi fosfato dello scheletro desossiribosio-fosfato. Kinemage... Exercises/E19

54 Motivi EF hand Il secondo motivo è specifico per il legame del calcio ed è presente in proteine che legano il calcio quali parvalbumina, calmodulina e troponina C, che regolano l attività cellulare. Questo particolare motivo, trovato per la prima volta nella parvalbumina, viene chiamato EF hand. Il loop fra le 2 eliche lega lo ione Ca 2+

55 Motivi -hairpin Il motivo più semplice è quello costituito da 2 filamenti antiparalleli adiacenti collegati da un tratto di loop. Questo motivo, chiamato -hairpin o unità, ricorre molto frequentemente nelle strutture antiparallele, come motivo isolato o come parte di un foglietto più complesso. La lunghezza del tratto di loop tra i filamenti è variabile, ma di solito è costituito da 2-5 aminoacidi (v. reverse-turns, come elementi di struttura secondaria). A questo motivo non è associata nessuna funzione specifica.

56 Motivi / Cross over connection Alla base dei motivi / sta il modo in cui 2 filamenti paralleli vengono collegati. Due filamenti paralleli adiacenti di solito sono connessi da un elica, che collega l estremità C-terminale del primo filamento con l estremità N- terminale del secondo filamento in modo tale che l asse dell elica sia parallelo ai filamenti. Questo motivo - - viene chiamato cross-over connection.

57 Motivi / Cross over connection La cross over connection consiste di di due filamenti paralleli, un elica e due loop (che possono variare notevolmente in lunghezza). L elica si impacca con i 2 filamenti, riparando dal solvente gli aminoacidi idrofobici dei filamenti. La cross over connection può essere considerata come un largo giro di superelica, a partire dal primo filamento, attraverso la connessione, fino al secondo filamento. La cross over connection può essere di tipo destrorso (a) o sinistrorso (b). Quasi tutte le proteine presentano una cross over connection destrorsa.
 che si ripiega indipendentemente in una struttura stabile.")
58 I domini I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini. I domini sono definiti come parte di una catena polipeptidica (o al limite l intera catena caso particolare) che si ripiega indipendentemente in una struttura stabile. I domini possono essere unità funzionali; spesso a domini diversi di una proteina sono associate funzioni diverse.
59 I domini Le proteine possono essere costituite da un singolo dominio o da molti (anche diverse dozzine).
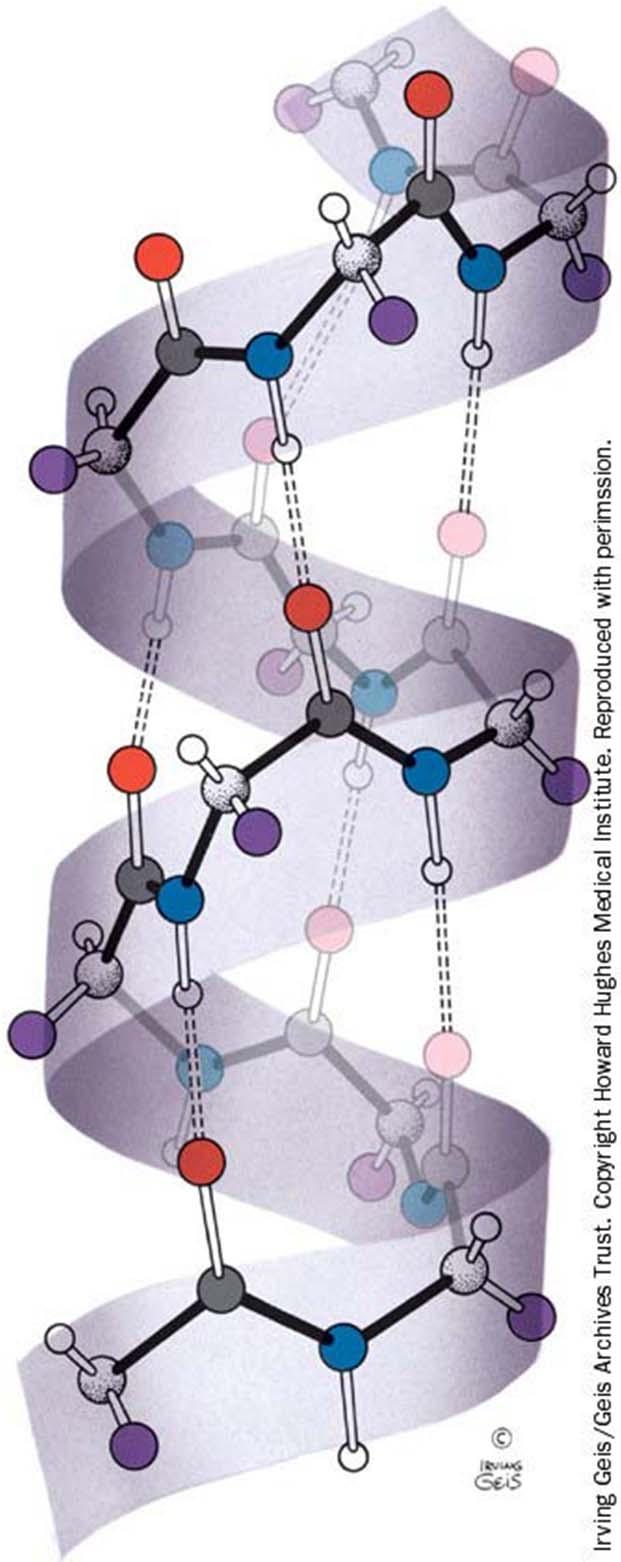
60 I domini Michael Levitt and Cyrus Chothia, sulla base di semplici considerazioni sulla connessione dei motivi, hanno classificato i domini in 3 gruppi principali, a seconda delle strutture secondarie e dei motivi coinvolti nella loro formazione: - domini helical bundle, globin-fold. - domini up-and-down, chiave greca, jelly roll, -helix - domini / barrel, open sheet, fold a ferro di cavallo - domini piccoli, contenenti metalli o ponti S-S


61 Domini helical bundle L helical bundle è costituito da 4 eliche disposte in modo tale che i loro assi siano reciprocamente quasi paralleli. Le catene laterali idrofobiche degli aminoacidi di ciascuna elica sono orientate verso l interno dell helical bundle, mentre le catene laterali idrofiliche degli aminoacidi sono rivolte verso l esterno. Le catene laterali idrofobiche rivolte verso l interno dell helical bundle sono così strettamente impaccate che non c è spazio per molecole d acqua.
.")
62 Domini helical bundle L helical bundle è un tipo di dominio che ricorre in svariate proteine (mioemeritrina, citocromo c' e b 562, ferritina, proteina del capside del virus del mosaico del tabacco). In questi casi le eliche sequenzialmente vicine sono sempre antiparallele.

63 Domini helical bundle L helical bundle si può anche formare con arrangiamenti topologici delle eliche diversi, come nel caso dell ormone umano della crescita. In questo caso, l helical bundle è formato da 2 coppie di eliche parallele che sono disposte in modo antiparallelo nell helical bundle. L impaccamento antiparallelo delle eliche conferisce loro maggiore stabilità in quanto i macrodipoli associati a ciascuna elica si annullano a vicenda.
64 Domini helical bundle Le eliche che costituiscono l helical bundle si impaccano con la modalità cresta-solco. Le creste e i solchi sono costituiti dalle catene laterali di aminoacidi separati da 3-4 residui (fig. c e b rispettivamente). La geometria dei solchi e delle creste di un elica dipende dalla geometria dell elica ma anche dalla sua sequenza aminoacidica.

65 Domini Globin fold Un altro impaccamento tipico delle eliche è quello osservato nel globin fold, caratteristico di emoglobine, mioglobine e ficocianine. Il globin fold è costituito da 8 eliche, indicate con le lettere A-H, collegate da loop piuttosto corti, in modo da disporsi a formare la tasca del sito attivo, che in mioglobina e emoglobina, ospita il gruppo eme. La lunghezza delle eliche varia considerevolmente: da 7 aminoacidi per l elica C a 28 aminoacidi per l elica H. Le eliche sono disposte in direzioni diverse, in modo tale che eliche adiacenti in sequenza non lo sono nella struttura, con l eccezione delle eliche G e H.
66 Domini Nonostante l elevato numero di possibili disposizioni di filamenti (a costituire foglietti antiparalleli) connessi da tratti di loop, i domini più frequentemente osservati sono solo 3: - up and down - chiave greca - jelly roll In generale, i filamenti sono disposti in modo tale da formare 2 foglietti impaccati uno con l altro, a formare un core idrofobico. Un discorso a parte meritano i domini a elica, una struttura recentemente scoperta.
 di 8 filamenti, come nel caso delle proteine che legano il retinolo o acidi grassi.")
67 Domini Up and down La topologia più semplice per foglietti antiparalleli è chiamata up and down: in essa i filamenti adiacenti sono collegati da regioni di loop ( -hairpin). Spesso l ultimo filamento è collegato al primo da legami idrogeno, a formare un barile ( barrel) di 8 filamenti, come nel caso delle proteine che legano il retinolo o acidi grassi. In questi casi, nelle sequenze dei filamenti catene laterali idrofobiche si alternano a catene laterali polari e cariche idrofiliche, in modo tale da formare il core idrofobico all interno del barile e da interagire esternamente con il solvente.

68 Kinemage Exercises/Ig-fold Domini Chiave greca Se si vogliono collegare 8 filamenti antiparalleli con una topologia diversa da quella up and down, ci sono solo 2 alternative: collegare il filamento n a quello n+3 oppure a quello n- 3. I due casi rappresentano le 2 possibili mani, cioè 2 opposte sceltechirali:intuttelestrutturedi proteine finora note si è trovata sempre e solo la topologia a chiave greca indicata in fig. a, quella destrorsa.


69 Domini Chiave greca Il motivo base è costituito da 4 filamenti : 3 presentano topologia up and down e sono collegati da -turns, seguiti da una connessione più lunga al quarto filamento, che è adiacente al primo. Esempi di topologia a chiave greca si osservano nelle superossido dismutasi a Cu,Zn, nei domini delle immunoglobuline e nella proteina cristallino.
.")
70 Domini Jelly roll Un altra topologia caratteristica dei filamenti antiparalleli è chiamata jelly roll. Consideriamo una stringa costituita da 8 filamenti antiparalleli, collegati a 2 a 2 da legami idrogeno (coppie 1-8, 2-7, 3-6 e 4-5). Se si avvolge questa stringa intorno ad un cilindro, in modo tale che i filamenti si trovino sulla superficie del barile mentre le regioni di loop alle 2 estremità, si ottiene la topologia jelly roll.

71 Domini elica Nel 1993 fu scoperta una nuova, inaspettata, struttura di tipo, la elica. La elica, che non è correlabile alle altre strutture, fuosservataper la prima volta nella struttura dell enzima batterico pectato liasi. Successivamente, sono state determinate altre strutture proteiche contenenti eliche (tra cui proteinasi batteriche extracellulari e la proteina tailspike del batteriofago P22). Nella elica la catena polipeptidica si avvolge a formare un ampia elica, costituita da brevi filamenti uniti fra loro da regioni di loop. Esistono 2tipidi elica: - elica a 2 foglietti - elica a 3 foglietti

72 Domini elicaa2foglietti La elica a 2 foglietti èlaformadi elica più semplice, osservata nelle proteinasi extracellulari batteriche. Ciascun giro di elica comprende 2 filamenti e 2 regioni di loop. Questa unità strutturale si ripete piu volte a formare una struttura elicoidale destrorsa costituita da 2 foglietti paralleli costituiti ciascuno da 3 filamenti, con un core idrofobico in mezzo. L unità strutturale base della elica a 2 foglietti è costituita da 18 aminoacidi, 3 in ciascun filamento e 6inciascunloop.Questaunità base è caratterizzata dalla sequenza consenso: (Gly-Gly-X-Gly-X-Asp-X-U-X) 2 [X: qualsiasi aminoacido; U: idrofobico, spesso Leu]
 collegati da 3 loop.")
73 Domini elica a 3 foglietti Una elica più complessa è presente nella pectato liasi e nella proteina tailspike del batteriofago P22. Nelle eliche a 3 foglietti ciascun giro di elica è formata da 3 segmenti (ciascuno costituito da 3 a 5 aminoacidi) collegati da 3 loop. La elica quindi è costituita da 3 foglietti paralleli, che formano i lati di un prisma. La sezione della elica, però, non è triangolare, a causa della disposizione dei foglietti.
74 Domini elica a 3 foglietti Due dei foglietti sono posizionati uno adiacente all altro, come nella elica a 2 foglietti, e il terzo foglietto è quasi perpendicolare agli altri 2. Un loop (a) è corto e quasi sempre formato solo da 2 aminoacidi con conformazione invariante e forma un angolo di circa 120 fra i 2 filamenti che unisce. Gli altri 2 loop (b, c) sono più lunghi e variano in dimensione e conformazione. I loop lunghi protrudono dai foglietti e probabilmente formano il sito attivo sulla superficie esterna della proteina. Kinemage Exercises/PecLyC La variabilità in numero e tipo di aminoacidi che costituiscono i 2 loop lunghi impedisce di determinare una sequenza consenso per la elica a3foglietti.
75 Domini elica a 3 foglietti Il numero di giri di elica nelle eliche a 3 foglietti è maggiore di quello trovato nelle eliche a 2 foglietti. La pectato liasi consiste di 7 giri di elica (d = 4.86 Å), è lunga 34 Å e ha un diametro di Å, mentre la elica della proteina tailspike del batteriofago P22 è formata da 13 giri.
.")
.")
76 Domini elica sinistrorsa Nel 1995 è stata osservata una elica sinistrorsa, nella struttura dell enzima UDP-N acetilglucosammina transferasi (LpxA). La elica sinistrorsa è l unica struttura di tipo parallelo che puo essere interpretata come una cross-over connection fra i filamenti paralleli di tipo sinistroso (vedi strutture supersecondarie). Gli enzimi che presentano la elica sinistrorsa sono caratterizzati dalla ripetizione dell esapeptide: [LIV]-[GAED]-X-X-[STAV]-X

77 I domini
78 Domini / La cross over connection è l unità costitutiva su cui si basa la topologia di 3 tipi di domini / osservati nelle proteine: - / barrel - motivi ricchi di Leu (fold a ferro di cavallo) - / open sheet
 Le eliche stanno tutte da un lato del foglietto parallelo. L ultimo filamento della prima cross over connection è adiacente al primo filamento della seconda cross over connection.")
79 Domini / Due cross over connections possono essere collegate in 2 modi diversi (tramite un elica), per formare un foglietto parallelo di 4 filamenti. 1) Le eliche stanno tutte da un lato del foglietto parallelo. L ultimo filamento della prima cross over connection è adiacente al primo filamento della seconda cross over connection. I filamenti sono nell ordine Impaccamento tipico degli / barrel e del fold a ferro di cavallo.

80 Domini / 2) Le eliche stanno sui lati opposti del foglietto parallelo. I primi filamenti di ciascuna cross over connection sono adiacenti. I filamenti sono nell ordine Impaccamento tipico degli / open sheet.

81 Domini / barrel Un / barrel è costituito da 8 filamenti paralleli, posizionati in modo tale che il filamento 8 sia adiacente e faccia legami idrogeno con quello 1. Di solito le cross over connection fra i filamenti paralleli sono eliche. In più c è un elica dopo l ultimo filamento. Esistono anche / barrel costituiti da 10 filamenti paralleli. L / barrel è un tipo di dominio che coinvolge almeno 200 aminoacidi ed è tipico di molti enzimi. E chiamato anche TIM barrel dalla struttura dell enzima trioso fosfato isomerasi, dove fu osservato per la prima volta.

82 Kinemage Exercises/ab_barrel Domini / barrel Nell / barrel le catene laterali idrofobiche delle eliche si impaccano con le catene laterali idrofobiche dei filamenti. Le interazioni fra le eliche e i filamenti coinvolgono principalmente aminoacidi del tipo Val, Ile, Leu, che costituiscono circa il 40% degli aminoacidi che costituiscono i filamenti del barile. Le altre catene laterali degli aminoacidi (in posizione 1, 3, 5, ) dei filamenti sono rivolte verso l interno del barile e formano un core idrofobico strettamente impaccato. Tra le catene laterali ci sono anche Arg, Lys e Gln, che presentano un gruppo polare, rivolto verso le 2 estremità del barile, in modo da poter interagire con il solvente esterno.

83 Kinemage Exercises/E12 Domini / barrel Tutti gli / barrel osservati nelle proteine conosciute hanno funzioni enzimatiche: isomerizzazione di piccole molecole di zucchero, trasferimento di gruppi fosfato, degradazione di zuccheri polimerici. In tutti gli / barrelilsitoattivositrova nella stessa posizione, in una tasca costituita dagli 8 loop che connettono l estremità C-terminale dei filamenti con l estremità N-terminale delle eliche. Gli aminoacidi che partecipano al legame del substrato e alla catalisi appartengono a queste regioni di loop.

84 Domini / Fold a ferro di cavallo I filamenti formano un foglietto parallelo e tutte le eliche sono su uno stesso lato del foglietto. I filamenti formano una struttura aperta ricurva, che ricorda un ferro di cavallo, conle eliche sul lato esterno e il foglietto che forma la parte interna del ferro di cavallo. Un lato del foglietto si interfaccia con le eliche, formando un core idrofobico, mentre l altro lato del foglietto è esposto al solvente, caratteristica che gli altri motivi / non hanno.

85 Kinemage Exercises/a-b_open_sht Domini / open sheet Nell / open sheet le eliche si trovano su entrambi i lati del foglietto parallelo. Ciascun filamento fornisce le catene laterali degli aminoacidi idrofobici per impaccarsi con le eliche in 2 regioni idrofobiche, una per ciascun lato del foglietto. In questo dominio / ci sono sempre 2 filamenti (1 e 4 in figura) nella parte interna del foglietto le cui connessioni al filamento vicino sono sui lati opposti del foglietto. Uno dei loop uscenti da uno dei filamenti va sopra il foglietto, mentre l altro loop va sotto.


86 Domini / open sheet Il punto in cui si ha l inversione del lato del foglietto che viene coperto dalle eliche è chiamato switch point. La zona dello switch point, all estremità C- terminale del foglietto, definisce una tasca, in cui si trovano sempre i siti di legame di questa classe di proteine. Negli / open sheet non ci sono restrizioni geometriche sul numero dei filamenti coinvolti: il numero varia da 4 a 10.
.")
87 Domini / open sheet Molti enzimi, e in particolare alcune deidrogenasi, hanno il dominio che lega il NAD costituito da due unità aformareildinucleotidebinding fold o Rossmann fold (questo tipo di fold fu identificato per la prima volta nell enzima lattato deidrogenasi da M. Rossmann, nel 1970). Il Rossmann fold è caratterizzato dalla presenza della sequenza consenso: GXGXXG(17X)D coinvolta nel legame del dinucleotide.

88 Domini / open sheet I residui GXGXXG, posti nel loop fra il primo filamento e la susseguente elica, determinano l interazione tra la proteina ed il gruppo pirofosfato. Asp 193 Gly 172 Gly 175 Alla fine del secondo filamento parallelo la catena laterale carbossilica di un acido aspartico o glutammico interagisce con il diolo del gruppo ribosio. Gly 170
89 Web sites for domain analysis home.php
90
91 Domini: altro Alcune proteine presentano domini che non rientrano nella classificazione fatta in precedenza e quindi formano gruppi a parte. Alcuni esempi sono: - proteine costituite dalla combinazione di motivi e a formare un foglietto antiparallelo impaccato contro un certo numero di eliche. Strutture di questo tipo vengono classificate come appartenenti ad un quarto tipo di domini, chiamato. - piccole proteine, ricche di ponti disolfuro o di ioni metallo. La presenza dei ponti disolfuro o degli ioni metallo sembra influenzi fortemente la struttura di queste proteine e spesso sembrano la versione distorta di proteine più regolari.

92 Polypeptide backbone and disulfide bonds of native BPTI.


93 The second zinc finger motif of Zif268
Domini nelle Proteine
 Struttura secondaria, Motivi e Domini nelle Proteine Proprietà generali Le forme ioniche degli aminoacidi, senza considerare alcuna ionizzazione delle catene laterali. Proprietà generali Tutti gli aminoacidi
Struttura secondaria, Motivi e Domini nelle Proteine Proprietà generali Le forme ioniche degli aminoacidi, senza considerare alcuna ionizzazione delle catene laterali. Proprietà generali Tutti gli aminoacidi
formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita i da unoo più domini.
 Idomini(I) I domini I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita i da unoo più domini. I domini sono definiti come parte
Idomini(I) I domini I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita i da unoo più domini. I domini sono definiti come parte
Macromolecole Biologiche. I domini (II)
") I domini (II) Domini β Nonostante l elevato numero di possibili disposizioni di filamenti β (a costituire foglietti β antiparalleli) connessi da tratti di loop, i domini β più frequentemente osservati
I domini (II) Domini β Nonostante l elevato numero di possibili disposizioni di filamenti β (a costituire foglietti β antiparalleli) connessi da tratti di loop, i domini β più frequentemente osservati
Macromolecole Biologiche. I domini (I)
") I domini (I) I domini I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini. I domini sono definiti come una
I domini (I) I domini I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini. I domini sono definiti come una
amminico è legato all atomo di carbonio immediatamente adiacente al gruppo carbonilico e hanno la seguente
 Gli amminoacidi naturali sono α-amminoacidi : il gruppo amminico è legato all atomo di carbonio immediatamente adiacente al gruppo carbonilico e hanno la seguente formula generale: gruppo funzionale carbossilico
Gli amminoacidi naturali sono α-amminoacidi : il gruppo amminico è legato all atomo di carbonio immediatamente adiacente al gruppo carbonilico e hanno la seguente formula generale: gruppo funzionale carbossilico
Macromolecole Biologiche. I domini (III)
") I domini (III) Domini α/β La cross over connection è l unità costitutiva su cui si basa la topologia di 3 tipi di domini α/β osservati nelle proteine: - α/β barrel - motivi ricchi di Leu (fold a ferro
I domini (III) Domini α/β La cross over connection è l unità costitutiva su cui si basa la topologia di 3 tipi di domini α/β osservati nelle proteine: - α/β barrel - motivi ricchi di Leu (fold a ferro
I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini.
 I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini. I domini sono definiti come una catena polipeptidica
I motivi generalmente si combinano a formare strutture globulari compatte, chiamate domini. Una proteina può essere costituita da uno o più domini. I domini sono definiti come una catena polipeptidica
Le proteine. Polimeri composto da 20 diversi aminoacidi
 Le proteine Polimeri composto da 20 diversi aminoacidi (D. Voet, J.G. Voet, Biochemistry, 3 ed., John Wiley & Sons, 2004) PROTEINE come ATTUATORI nella cellula Trasporto elettronico Trasporto di ioni e
Le proteine Polimeri composto da 20 diversi aminoacidi (D. Voet, J.G. Voet, Biochemistry, 3 ed., John Wiley & Sons, 2004) PROTEINE come ATTUATORI nella cellula Trasporto elettronico Trasporto di ioni e
scaricato da www.sunhope.it Proteine semplici costituite dai soli amminoacidi
 Proteine semplici costituite dai soli amminoacidi Proteine coniugate costituite dagli amminoacidi + porzioni di natura non amminoacidica dette GRUPPI PROSTETICI Le Proteine coniugate prive del gruppo prostetico
Proteine semplici costituite dai soli amminoacidi Proteine coniugate costituite dagli amminoacidi + porzioni di natura non amminoacidica dette GRUPPI PROSTETICI Le Proteine coniugate prive del gruppo prostetico
Le Biomolecole I parte. Lezioni d'autore di Giorgio Benedetti
 Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
I gruppi R differenziano i 20 amminoacidi standard. Tratto da D. Voet, G. Voet e C.W. Pratt Fondamenti di biochimica
 Gli aminoacidi NOMENCLATURA Aminoacido Abbr. tre lettere Abbr. una lettera Aminoacido Abbr. tre lettere Abbr. una lettera Alanina ALA A Lisina LYS K Arginina ARG R Metionina MET M Asparagina ASN N Fenilalanina
Gli aminoacidi NOMENCLATURA Aminoacido Abbr. tre lettere Abbr. una lettera Aminoacido Abbr. tre lettere Abbr. una lettera Alanina ALA A Lisina LYS K Arginina ARG R Metionina MET M Asparagina ASN N Fenilalanina
AMINOACIDI - 1 AMINOACIDI - 2
 AMINOAIDI - 1 Proteine (gr. pròtos = primo) 50-80% peso secco cellulare GENE POTEINA EFFETTO proteine calore idrolisi acida o alcalina α-aminoacidi proteasi Struttura generale degli α-aminoacidi primari
AMINOAIDI - 1 Proteine (gr. pròtos = primo) 50-80% peso secco cellulare GENE POTEINA EFFETTO proteine calore idrolisi acida o alcalina α-aminoacidi proteasi Struttura generale degli α-aminoacidi primari
AMINOACIDI. GENE PROTEINA EFFETTO. calore. idrolisi acida (o alcalina) Struttura generale degli α-aminoacidi primari (standard, normali):
 Struttura generale degli α-aminoacidi primari (standard, normali):") AMINOAIDI. Proteine (gr. pròtos = primo) > 50% peso secco cellulare GENE POTEINA EFFETTO proteine calore idrolisi acida (o alcalina) α-aminoacidi Struttura generale degli α-aminoacidi primari (standard,
AMINOAIDI. Proteine (gr. pròtos = primo) > 50% peso secco cellulare GENE POTEINA EFFETTO proteine calore idrolisi acida (o alcalina) α-aminoacidi Struttura generale degli α-aminoacidi primari (standard,
CARBOIDRATI C H O ZUCCHERO SACCARIDE GLUCIDE CARBOIDRATO
 CARBOIDRATI ZUCCHERO SACCARIDE GLUCIDE CARBOIDRATO C H O carboidrati C n H 2n O n H C O C O Il glucosio è un monosaccaride con 6 atomi di carbonio GLUCOSIO Forma ciclica Forma lineare a ph 7 circa lo 0,0026%
CARBOIDRATI ZUCCHERO SACCARIDE GLUCIDE CARBOIDRATO C H O carboidrati C n H 2n O n H C O C O Il glucosio è un monosaccaride con 6 atomi di carbonio GLUCOSIO Forma ciclica Forma lineare a ph 7 circa lo 0,0026%
REPLICAZIONE DEL DNA
 REPLICAZIONE DEL DNA La replicazione (o anche duplicazione) è il meccanismo molecolare attraverso cui il DNA produce una copia di sé stesso. Ogni volta che una cellula si divide, infatti, l'intero genoma
REPLICAZIONE DEL DNA La replicazione (o anche duplicazione) è il meccanismo molecolare attraverso cui il DNA produce una copia di sé stesso. Ogni volta che una cellula si divide, infatti, l'intero genoma
AMMINOACIDI E PROTEINE
 AMMINOACIDI E PROTEINE Vengono chiamate amminoacidi quelle molecole organiche in cui sono contemporaneamente presenti sia un gruppo acido carbossilico -COO che un gruppo amminico -N2. Una molecola appartenente
AMMINOACIDI E PROTEINE Vengono chiamate amminoacidi quelle molecole organiche in cui sono contemporaneamente presenti sia un gruppo acido carbossilico -COO che un gruppo amminico -N2. Una molecola appartenente
Dipartimento Di Scienze Della Vita STRUTTURA DELLE PROTEINE
 Dipartimento Di Scienze Della Vita STRUTTURA DELLE PROTEINE LE PROTEINE possono assumere 4 LIVELLI DI ORGANIZZAZIONE STRUTTURA PRIMARIA SEQUENZA degli amminoacidi STRUTTURA SECONDARIA Ripiegamento locale
Dipartimento Di Scienze Della Vita STRUTTURA DELLE PROTEINE LE PROTEINE possono assumere 4 LIVELLI DI ORGANIZZAZIONE STRUTTURA PRIMARIA SEQUENZA degli amminoacidi STRUTTURA SECONDARIA Ripiegamento locale
Gli amminoacidi Il legame peptidico Motivi strutturali classificazione, architettura topologia delle strutture tridimensionali di proteine.
 Struttura di proteine Gli amminoacidi Il legame peptidico Motivi strutturali classificazione, architettura topologia delle strutture tridimensionali di proteine. Correlazioni struttura-funzione Gli amminoacidi
Struttura di proteine Gli amminoacidi Il legame peptidico Motivi strutturali classificazione, architettura topologia delle strutture tridimensionali di proteine. Correlazioni struttura-funzione Gli amminoacidi
2011 - G. Licini, Università di Padova. La riproduzione a fini commerciali è vietata
 Ammino acidi Composto che contiene una funziome acida e amminica. Usualmente però con amminoacidi si intendono gli alfa- amminoacidi. Tra questi composti ve ne sono 20 che vengono definiti geneticamente
Ammino acidi Composto che contiene una funziome acida e amminica. Usualmente però con amminoacidi si intendono gli alfa- amminoacidi. Tra questi composti ve ne sono 20 che vengono definiti geneticamente
Macromolecole Biologiche. Chimica Biologica A.A. 2010-2011. Struttura Terziaria
 Macromolecole Biologiche Chimica Biologica A.A. 2010-2011 Struttura Terziaria Domini e struttura terziaria Struttura terziaria L arrangiamento spaziale degli amminoacidi di una singola catena polipeptidica
Macromolecole Biologiche Chimica Biologica A.A. 2010-2011 Struttura Terziaria Domini e struttura terziaria Struttura terziaria L arrangiamento spaziale degli amminoacidi di una singola catena polipeptidica
STRUTTURA E FUNZIONE DELLE PROTEINE
 STRUTTURA E FUNZIONE DELLE PROTEINE Le PROTEINE sono i biopolimeri maggiormente presenti all interno delle cellule, dal momento che costituiscono dal 40 al 70% del peso secco. Svolgono funzioni biologiche
STRUTTURA E FUNZIONE DELLE PROTEINE Le PROTEINE sono i biopolimeri maggiormente presenti all interno delle cellule, dal momento che costituiscono dal 40 al 70% del peso secco. Svolgono funzioni biologiche
Aminoacidi e proteine
 Prof. Giorgio Sartor Aminoacidi e proteine Copyright 2001-2012 by Giorgio Sartor. Versione 1.7.1 mar 2012 All rights reserved. Le cellule V.1.7.1 gsartor 2001-2012 Aminoacidi e proteine -2- Aminoacidi
Prof. Giorgio Sartor Aminoacidi e proteine Copyright 2001-2012 by Giorgio Sartor. Versione 1.7.1 mar 2012 All rights reserved. Le cellule V.1.7.1 gsartor 2001-2012 Aminoacidi e proteine -2- Aminoacidi
BIOMOLECOLE STRUTTURA E FUNZIONE
 BIOMOLECOLE STRUTTURA E FUNZIONE Lo studio delle relazioni tra struttura e funzione nelle biomolecole è uno degli aspetti più importanti per la comprensione del funzionamento dei processi biologici La
BIOMOLECOLE STRUTTURA E FUNZIONE Lo studio delle relazioni tra struttura e funzione nelle biomolecole è uno degli aspetti più importanti per la comprensione del funzionamento dei processi biologici La
aa 2013-14 Proteine Struttura delle Proteine α Amminoacidi
 Proteine Biopolimeri degli α-amino acidi. Amino acidi sono uniti attraverso il legame peptidico. Alcune funzioni: Struttura (collagene, cheratina ecc.) Enzimi (maltasi, deidrogenasi ecc) Trasporto (albumine,
Proteine Biopolimeri degli α-amino acidi. Amino acidi sono uniti attraverso il legame peptidico. Alcune funzioni: Struttura (collagene, cheratina ecc.) Enzimi (maltasi, deidrogenasi ecc) Trasporto (albumine,
Metalli in Biologia. Metalli e leganti bio
 Metalli in Biologia Metalli e leganti bio La struttura della mioglobina La struttura della emoglobina John Cowdery Kendrew MRC, Cambridge, UK Nobel Prize in 1962 Max Ferdinand Perutz MRC, Cambridge, UK
Metalli in Biologia Metalli e leganti bio La struttura della mioglobina La struttura della emoglobina John Cowdery Kendrew MRC, Cambridge, UK Nobel Prize in 1962 Max Ferdinand Perutz MRC, Cambridge, UK
Struttura delle proteine
 Struttura delle proteine Nelle proteine vi sono quattro livelli di organizzazione strutturale Struttura Primaria: sequenza di aminoacidi legati tra loro da legami peptidici Tutte le proteine esistenti
Struttura delle proteine Nelle proteine vi sono quattro livelli di organizzazione strutturale Struttura Primaria: sequenza di aminoacidi legati tra loro da legami peptidici Tutte le proteine esistenti
INTRODUZIONE ALLA PROTEINE BCP 1-2
 INTRODUZIONE ALLA BIOCHIMICA DELLE PROTEINE BCP 1-2 Pietre miliari Lehninger 1973 Biochemistry Proteine ricombinanti Protein engineering Cristallografia, NMR, EM, MS, spettroscopie... Computing Bioinformaticai
INTRODUZIONE ALLA BIOCHIMICA DELLE PROTEINE BCP 1-2 Pietre miliari Lehninger 1973 Biochemistry Proteine ricombinanti Protein engineering Cristallografia, NMR, EM, MS, spettroscopie... Computing Bioinformaticai
Le proteine rappresentano gli elementi strutturali e funzionali più importanti nei sistemi viventi. Qualsiasi processo vitale dipende da questa
 Gli amminoacidi Le proteine rappresentano gli elementi strutturali e funzionali più importanti nei sistemi viventi. Qualsiasi processo vitale dipende da questa classe di molecole: p. es. la catalisi delle
Gli amminoacidi Le proteine rappresentano gli elementi strutturali e funzionali più importanti nei sistemi viventi. Qualsiasi processo vitale dipende da questa classe di molecole: p. es. la catalisi delle
Gli amminoacidi Gli amminoacidi sono dei composti polifunzionali che hanno formula generale:
 Gli amminoacidi Gli amminoacidi sono dei composti polifunzionali che hanno formula generale: N 2 Il nome ordinario degli amminoacidi prevale su quello della nomenclatura IUPA. Si possono avere α-amminoacidi,
Gli amminoacidi Gli amminoacidi sono dei composti polifunzionali che hanno formula generale: N 2 Il nome ordinario degli amminoacidi prevale su quello della nomenclatura IUPA. Si possono avere α-amminoacidi,
Prof. Maria Nicola GADALETA
 Prof. Maria Nicola GADALETA Email: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA Email: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
NH 2 CHCOOH AMINOACIDI PRESENTI NELLE PROTEINE. Abbreviazione internazionale. Nome. acido aspartico acido glutammico. CXasparagina.
 Le proteine sono composti organici quaternari; esse sono costituite infatti da C,H,N e O. Pertanto rappresentano l'unica fonte di azoto del nostro organismo. Le proteine si formano per polimerizzazione
Le proteine sono composti organici quaternari; esse sono costituite infatti da C,H,N e O. Pertanto rappresentano l'unica fonte di azoto del nostro organismo. Le proteine si formano per polimerizzazione
MEMBRANE. struttura: fosfolipidi e proteine
 MEMBRANE struttura: fosfolipidi e proteine I lipidi sono sostanze di origine biologica insolubili in acqua. Vi fanno parte: trigliceridi, fosfolipidi, colesterolo, sfingolipidi, alcoli alifatici, cere,
MEMBRANE struttura: fosfolipidi e proteine I lipidi sono sostanze di origine biologica insolubili in acqua. Vi fanno parte: trigliceridi, fosfolipidi, colesterolo, sfingolipidi, alcoli alifatici, cere,
LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI PROTEINE. sono ACIDI NUCLEICI. molecole complesse = POLIMERI. formate dall'unione di
 LE BIOMOLECOLE LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI LE BIOMOLECOLE sono LIPIDI PROTEINE ACIDI NUCLEICI molecole complesse = POLIMERI formate dall'unione di molecole semplici = MONOMERI
LE BIOMOLECOLE LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI LE BIOMOLECOLE sono LIPIDI PROTEINE ACIDI NUCLEICI molecole complesse = POLIMERI formate dall'unione di molecole semplici = MONOMERI
Continua. Peptidasi H 2 O
 Continua Peptidasi H 2 O Classificazione delle peptidasi 1. Meccanismo catalitico 2. Tipo di reazione catalizzata 3. Struttura molecolare e omologia 1. Meccanismo catalitico (mostrato per la chimotripsina)
Continua Peptidasi H 2 O Classificazione delle peptidasi 1. Meccanismo catalitico 2. Tipo di reazione catalizzata 3. Struttura molecolare e omologia 1. Meccanismo catalitico (mostrato per la chimotripsina)
Le proteine. Le proteine sono macromolecole che presentano differenze funzionali e strutturali
 Le proteine Le proteine sono macromolecole che presentano differenze funzionali e strutturali LE PROTEINE HANNO FUNZIONI BIOLOGICHE DIVERSE enzimi proteine di trasporto proteine strutturali proteine di
Le proteine Le proteine sono macromolecole che presentano differenze funzionali e strutturali LE PROTEINE HANNO FUNZIONI BIOLOGICHE DIVERSE enzimi proteine di trasporto proteine strutturali proteine di
Visualizzazioni 3D. Informatica. Matrice di voxel. Tipi di dato. Dati vettoriali. Tecniche di rappresentazione
 Informatica Lezione VIII Visualizzazione 3D di proteine Visualizzazioni 3D Rappresentazione di strutture/oggetti tridimensionali Risultato di un esperimento modello teorico dati fisici astrazione 1 Lezione
Informatica Lezione VIII Visualizzazione 3D di proteine Visualizzazioni 3D Rappresentazione di strutture/oggetti tridimensionali Risultato di un esperimento modello teorico dati fisici astrazione 1 Lezione
corta catena (meno di 20 ammino acidi) mancanza di una struttura spaziale organizzata lunga catena di ammino acidi struttura spaziale organizzata
 mancanza di una struttura spaziale organizzata lunga catena di ammino acidi struttura spaziale organizzata") STRUTTURA DELLE PROTEINE Peptide: corta catena (meno di 20 ammino acidi) mancanza di una struttura spaziale organizzata Polipeptide (proteina): lunga catena di ammino acidi struttura spaziale organizzata
STRUTTURA DELLE PROTEINE Peptide: corta catena (meno di 20 ammino acidi) mancanza di una struttura spaziale organizzata Polipeptide (proteina): lunga catena di ammino acidi struttura spaziale organizzata
4x4x4=4 3 =64 codoni. 20 aminoacidi
 4x4x4=4 3 =64 codoni 20 aminoacidi 1 Le 20 diverse catene laterali (gruppo R) che costituiscono gli aminoacidi si differenziano considerevolmente per dimensioni, volume e per le loro caratteristiche fisico-chimiche,
4x4x4=4 3 =64 codoni 20 aminoacidi 1 Le 20 diverse catene laterali (gruppo R) che costituiscono gli aminoacidi si differenziano considerevolmente per dimensioni, volume e per le loro caratteristiche fisico-chimiche,
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA. Angela Chambery Lezione 7
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 7 La struttura delle proteine Concetti chiave: La struttura terziaria di una proteina descrive il ripiegamentodei suoi elementi
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 7 La struttura delle proteine Concetti chiave: La struttura terziaria di una proteina descrive il ripiegamentodei suoi elementi
3.3. UMAMI NEGLI ALIMENTI
 3.3. UMAMI NEGLI ALIMENTI In questo capitolo saranno esaminati i vari cibi, ad esclusione di quelli ittici che verranno presi in considerazione successivamente, allo scopo di scoprire se in essi è presente,
3.3. UMAMI NEGLI ALIMENTI In questo capitolo saranno esaminati i vari cibi, ad esclusione di quelli ittici che verranno presi in considerazione successivamente, allo scopo di scoprire se in essi è presente,
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA. Angela Chambery Lezione 9
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 9 Funzioni delle proteine Concetti chiave: La varietà strutturale delle proteine consente loro di svolgere un enorme quantità
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 9 Funzioni delle proteine Concetti chiave: La varietà strutturale delle proteine consente loro di svolgere un enorme quantità
Lezione 2. Sommario. Bioinformatica. Lezione 2: AMMINOACIDI E POLIPEPTIDI Aminoacidi e proteine. Mauro Ceccanti e Alberto Paoluzzi
 Lezione 2 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 2: AMMINOACIDI E POLIPEPTIDI Aminoacidi e proteine Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università
Lezione 2 Bioinformatica Mauro Ceccanti e Alberto Paoluzzi Lezione 2: AMMINOACIDI E POLIPEPTIDI Aminoacidi e proteine Dip. Informatica e Automazione Università Roma Tre Dip. Medicina Clinica Università
PROTEINE. Amminoacidi
 PROTEINE Le proteine sono le macromolecole alla base delle attività cellulari. Sono oltre diecimila per cellula, dove svolgono differenti funzioni: Sono ad esempio: enzimi: aumentano la velocità delle
PROTEINE Le proteine sono le macromolecole alla base delle attività cellulari. Sono oltre diecimila per cellula, dove svolgono differenti funzioni: Sono ad esempio: enzimi: aumentano la velocità delle
Prof. Maria Nicola GADALETA
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Peptidi, proteine ed e nzim i i 1
 Peptidi, proteine ed enzimi 1 Gli amminoacidi possono formare catene Due amminoacidi possono unirsi tra loro attraverso il legame ammidico detto legame peptidico, tra il gruppo NH 2 di un amminoacido e
Peptidi, proteine ed enzimi 1 Gli amminoacidi possono formare catene Due amminoacidi possono unirsi tra loro attraverso il legame ammidico detto legame peptidico, tra il gruppo NH 2 di un amminoacido e
LE PROTEINE. Le proteine sono i biopolimeri piu' abbondanti negli organismi viventi e svolgono funzioni biologiche di fondamentale importanza:
 LE PRTEINE Le proteine sono i biopolimeri piu' abbondanti negli organismi viventi e svolgono funzioni biologiche di fondamentale importanza: PRTEINE CATALITICHE PRTEINE REGLATRICI PRTEINE DIFESA PRTEINE
LE PRTEINE Le proteine sono i biopolimeri piu' abbondanti negli organismi viventi e svolgono funzioni biologiche di fondamentale importanza: PRTEINE CATALITICHE PRTEINE REGLATRICI PRTEINE DIFESA PRTEINE
Seminario. Domini modulari delle proteine 1
 Seminario Proteine della matrice DOMINI E MODULI Domini modulari delle proteine 1 La maggior parte dei peptidi consiste in disposizioni lineari di regioni globulari, ripiegate in modo indipendente, dette
Seminario Proteine della matrice DOMINI E MODULI Domini modulari delle proteine 1 La maggior parte dei peptidi consiste in disposizioni lineari di regioni globulari, ripiegate in modo indipendente, dette
Capitolo 17. Risposte alle domande interne al capitolo. 17.1 (p. 501) a. glicina. b. prolina. c. treonina. d. aspartato
 a. glicina. b. prolina. c. treonina. d. aspartato") apitolo 17 Risposte alle domande interne al capitolo 17.1 (p. 501) a. glicina b. prolina c. treonina d. aspartato e. 17.2 (p. 501) a. La glicina è un aminoacido idrofobico. b. La prolina è un aminoacido
apitolo 17 Risposte alle domande interne al capitolo 17.1 (p. 501) a. glicina b. prolina c. treonina d. aspartato e. 17.2 (p. 501) a. La glicina è un aminoacido idrofobico. b. La prolina è un aminoacido
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
La funzione delle proteine dipende dalla loro struttura tridimensionale
 La funzione delle proteine dipende dalla loro struttura tridimensionale La struttura dipende dal ripiegamento di particolari sequenze aminoacidiche La sequenza aminoacidica della catena polipeptidica è
La funzione delle proteine dipende dalla loro struttura tridimensionale La struttura dipende dal ripiegamento di particolari sequenze aminoacidiche La sequenza aminoacidica della catena polipeptidica è
LIPIDI. I lipidi sono suddivisi in due gruppi principali.
 LIPIDI I lipidi sono una classe eterogenea di composti organici, raggruppati sulla base della loro solubilità. Sono insolubili in acqua, ma solubili nei solventi organici non polari, quali l etere dietilico,
LIPIDI I lipidi sono una classe eterogenea di composti organici, raggruppati sulla base della loro solubilità. Sono insolubili in acqua, ma solubili nei solventi organici non polari, quali l etere dietilico,
DOMINI E MODULI 02/04/2014. Domini modulari delle proteine 2
 Domini modulari delle proteine 1 Proteine della matrice DOMINI E MODULI La maggior parte dei peptidi consiste in disposizioni lineari di regioni globulari, ripiegate in modo indipendente, dette domini,
Domini modulari delle proteine 1 Proteine della matrice DOMINI E MODULI La maggior parte dei peptidi consiste in disposizioni lineari di regioni globulari, ripiegate in modo indipendente, dette domini,
Livello di organizzazione degli esseri viventi
 Livello di organizzazione degli esseri viventi _Organismo; _Apparato; _Organo; _Tessuti; _Cellule; _Organelli cellulari; _Molecole. Atomo, elemento, molecola, composto, formula, legame, elettronegativita.
Livello di organizzazione degli esseri viventi _Organismo; _Apparato; _Organo; _Tessuti; _Cellule; _Organelli cellulari; _Molecole. Atomo, elemento, molecola, composto, formula, legame, elettronegativita.
Relazione struttura-funzione
 Biotecnologie applicate alla progettazione e sviluppo di molecole biologicamente attive A.A. 2010-2011 Modulo di Biologia Strutturale Relazione struttura-funzione Marco Nardini Dipartimento di Scienze
Biotecnologie applicate alla progettazione e sviluppo di molecole biologicamente attive A.A. 2010-2011 Modulo di Biologia Strutturale Relazione struttura-funzione Marco Nardini Dipartimento di Scienze
dieta vengono convertiti in composti dei corpi chetonici.
 Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
La biochimica è anche definita la chimica del C :
 Tutte le cellule viventi sono composte da macromolecole simili, costituite dalle stesse piccole molecole di base. La grande diversità è data dalle diverse combinazioni di 4 principali elementi C H O N
Tutte le cellule viventi sono composte da macromolecole simili, costituite dalle stesse piccole molecole di base. La grande diversità è data dalle diverse combinazioni di 4 principali elementi C H O N
- lipidi strutturali, costituenti fondamentali delle membrane cellulari ed intracellulari (fosfolipidi, glicolipidi e colesterolo)
") I lipidi, o grassi, costituiscono un gruppo eterogeneo di sostanze accomunate dalla proprietà fisica della insolubilità nei solventi polari (es. acqua) (idrofobicità) e dalla solubilità nei soventi organici
I lipidi, o grassi, costituiscono un gruppo eterogeneo di sostanze accomunate dalla proprietà fisica della insolubilità nei solventi polari (es. acqua) (idrofobicità) e dalla solubilità nei soventi organici
delegate a conservare e trasmettere l'informazione genetica. Oggi é noto che sono gli acidi
 8. Le proteine Il termine "proteina" (dal greco προτειος: di primaria importanza) fu coniato nel 1838 dal chimico svedese J. Berzelius 1 quando ancora si riteneva che le proteine fossero le molecole delegate
8. Le proteine Il termine "proteina" (dal greco προτειος: di primaria importanza) fu coniato nel 1838 dal chimico svedese J. Berzelius 1 quando ancora si riteneva che le proteine fossero le molecole delegate
Analisi quantitative
 Analisi quantitative Diversi metodi per la quantificazione delle proteine totali (reazioni generali delle proteine): 1. Dosaggio spettrofotometrico diretto 2. Metodi colorimetrici 1. Dosaggio spettrofotometrico
Analisi quantitative Diversi metodi per la quantificazione delle proteine totali (reazioni generali delle proteine): 1. Dosaggio spettrofotometrico diretto 2. Metodi colorimetrici 1. Dosaggio spettrofotometrico
NUCLEOTIDI e ACIDI NUCLEICI
 NUCLEOTIDI e ACIDI NUCLEICI Struttura dei nucleotidi Il gruppo fosfato conferisce carica negativa e proprietà acide FUNZIONI DEI NUCLEOTIDI MOLECOLE DI RISERVA DI ENERGIA L idrolisi dei nucleosidi trifosfato
NUCLEOTIDI e ACIDI NUCLEICI Struttura dei nucleotidi Il gruppo fosfato conferisce carica negativa e proprietà acide FUNZIONI DEI NUCLEOTIDI MOLECOLE DI RISERVA DI ENERGIA L idrolisi dei nucleosidi trifosfato
Marinella Bosetto Irene Lozzi. Elementi di biochimica agraria
 A07 25 Marinella Bosetto Irene Lozzi Elementi di biochimica agraria Copyright MMVI ARACNE editrice S.r.l. www.aracneeditrice.it info@aracneeditrice.it via Raffaele Garofalo, 133 A/B 00173 Roma (06) 93781065
A07 25 Marinella Bosetto Irene Lozzi Elementi di biochimica agraria Copyright MMVI ARACNE editrice S.r.l. www.aracneeditrice.it info@aracneeditrice.it via Raffaele Garofalo, 133 A/B 00173 Roma (06) 93781065
Il DNA e la duplicazione cellulare. Acidi nucleici: DNA, materiale ereditario
 Il DN e la duplicazione cellulare Il DN, materiale ereditario Struttura del DN Replicazione del DN Dal DN alla proteina Il odice genetico iclo cellulare Mitosi Meiosi Da Figura 8-11 ampbell & Reece cidi
Il DN e la duplicazione cellulare Il DN, materiale ereditario Struttura del DN Replicazione del DN Dal DN alla proteina Il odice genetico iclo cellulare Mitosi Meiosi Da Figura 8-11 ampbell & Reece cidi
proteasi (distrugge le proteine) batteri virulenti del ceppo S e del ceppo R
 batteri virulenti del ceppo S e del ceppo R") unità 1. La funzione del DN negli organismi La funzione del DN L acido desossiribonucleico o DN (dall inglese deoxyribonucleic acid) è la molecola informazionale delle cellule. Essa contiene e trasmette
unità 1. La funzione del DN negli organismi La funzione del DN L acido desossiribonucleico o DN (dall inglese deoxyribonucleic acid) è la molecola informazionale delle cellule. Essa contiene e trasmette
Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA. Angela Chambery Lezione 4
 Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA Angela Chambery Lezione 4 Scoperta degli amminoacidi: gli amminoacidi essenziali Gli amminoacidi essenziali sono quegli amminoacidi che un
Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA Angela Chambery Lezione 4 Scoperta degli amminoacidi: gli amminoacidi essenziali Gli amminoacidi essenziali sono quegli amminoacidi che un
Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA. Angela Chambery Lezione 2
 Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA Angela Chambery Lezione 2 Versatilità del carbonio nel formare legami covalenti La chimica degli organismi viventi è organizzata intorno al
Corso di Laurea in Farmacia Insegnamento di CHIMICA BIOLOGICA Angela Chambery Lezione 2 Versatilità del carbonio nel formare legami covalenti La chimica degli organismi viventi è organizzata intorno al
Traduzione dell informazione genetica (1)
") Traduzione dell informazione genetica (1) 1 Traduzione dell informazione genetica (2) Il processo negli eucarioti richiede: 70 diverse proteine ribosomiali >20 enzimi che attivano i precursori degli amminoacidi
Traduzione dell informazione genetica (1) 1 Traduzione dell informazione genetica (2) Il processo negli eucarioti richiede: 70 diverse proteine ribosomiali >20 enzimi che attivano i precursori degli amminoacidi
Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica.
 Concanavalina A Emoglobina subunità Trioso fosfato isomerasi Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica. 1 La conformazione è
Concanavalina A Emoglobina subunità Trioso fosfato isomerasi Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica. 1 La conformazione è
N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi
 Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Lipidi: funzioni. Strutturale. Riserva energetica. Segnale.
 Lipidi: funzioni. Strutturale. Riserva energetica. Segnale. Lipidi:classificazione. Saponificabili Non saponificabili Semplici Complessi Prostaglandine Steroidi Cere Trigliceridi Fosfogliceridi Sfingolipidi
Lipidi: funzioni. Strutturale. Riserva energetica. Segnale. Lipidi:classificazione. Saponificabili Non saponificabili Semplici Complessi Prostaglandine Steroidi Cere Trigliceridi Fosfogliceridi Sfingolipidi
Rappresentazione dei Dati Biologici
 Rappresentazione dei Dati Biologici CORSO DI BIOINFORMATICA C.d.L. Ingegneria Informatica e Biomedica Outline Proteine ed Amminoacidi Rappresentazione di Amminoacidi Rappresentazione delle strutture Proteiche
Rappresentazione dei Dati Biologici CORSO DI BIOINFORMATICA C.d.L. Ingegneria Informatica e Biomedica Outline Proteine ed Amminoacidi Rappresentazione di Amminoacidi Rappresentazione delle strutture Proteiche
Amminoacidi e Proteine
 Amminoacidi e Proteine Struttura generale di un α-amminoacido R = catena laterale AMMINOACIDI (AA) CELLULARI Gli amminoacidi presenti nella cellula possono essere il prodotto di idrolisi delle proteine
Amminoacidi e Proteine Struttura generale di un α-amminoacido R = catena laterale AMMINOACIDI (AA) CELLULARI Gli amminoacidi presenti nella cellula possono essere il prodotto di idrolisi delle proteine
Digestione delle proteine: 6 fasi
 orletto a spazzola Digestione delle proteine: 6 fasi 1. Idrolisi gastrica del legame peptidico 2. Digestione a peptidi più piccoli da parte delle proteasi pancreatiche nel lume dell intestino tenue 3.
orletto a spazzola Digestione delle proteine: 6 fasi 1. Idrolisi gastrica del legame peptidico 2. Digestione a peptidi più piccoli da parte delle proteasi pancreatiche nel lume dell intestino tenue 3.
Gerarchia della struttura delle proteine
 Si indica con CONFORMAZIONE la disposizione tridimensionale degli atomi di una molecola, cioè la loro organizzazione spaziale. Gerarchia della struttura delle proteine struttura primaria: sequenza degli
Si indica con CONFORMAZIONE la disposizione tridimensionale degli atomi di una molecola, cioè la loro organizzazione spaziale. Gerarchia della struttura delle proteine struttura primaria: sequenza degli
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici
 I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica. Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA.
 Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA. In genere si ottengono trattando il DNA con agenti chimici (es.
Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA. In genere si ottengono trattando il DNA con agenti chimici (es.
MODIFICAZIONI POST-TRADUZIONALI DELLE PROTEINE
 MODIFICAZIONI POST-TRADUZIONALI DELLE PROTEINE Nell ultima fase della sintesi proteica la catena polipeptidica neosintetizzata assume spontaneamente la sua conformazione nativa (massimo numero di legami
MODIFICAZIONI POST-TRADUZIONALI DELLE PROTEINE Nell ultima fase della sintesi proteica la catena polipeptidica neosintetizzata assume spontaneamente la sua conformazione nativa (massimo numero di legami
Una proteina nella rete: Caccia al tesoro bioinformatica
 Una proteina nella rete: Caccia al tesoro bioinformatica Nel corso di questa attivita utilizzeremo alcune delle piu importanti banche dati disponibili in rete per cercare informazioni su una proteina.
Una proteina nella rete: Caccia al tesoro bioinformatica Nel corso di questa attivita utilizzeremo alcune delle piu importanti banche dati disponibili in rete per cercare informazioni su una proteina.
PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA
 PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA Svolgono la loro funzione legando reversibilmente l OSSIGENO. Aumentano la solubilità dell ossigeno nel plasma, da 3ml/L a 220 ml/l. La mioglobina
PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA Svolgono la loro funzione legando reversibilmente l OSSIGENO. Aumentano la solubilità dell ossigeno nel plasma, da 3ml/L a 220 ml/l. La mioglobina
Strutture molecolari della cellula: Bio-macromolecole. Prof. C. Guarino
 Strutture molecolari della cellula: Bio-macromolecole Prof. C. Guarino INTRO Ogni cellula vivente racchiude una pluralità di molecole diverse L acqua è l elemento dominante, nelle cellule vegetali e nei
Strutture molecolari della cellula: Bio-macromolecole Prof. C. Guarino INTRO Ogni cellula vivente racchiude una pluralità di molecole diverse L acqua è l elemento dominante, nelle cellule vegetali e nei
Elementi di Bioinformatica. Genomica. Introduzione
 Corso di Elementi di Bioinformatica Ingegneria Biomedica AA 2013-14 Elementi di Bioinformatica Genomica Introduzione Genomica Genomica (genomics) Riguarda lo studio del genoma degli organismi viventi e,
Corso di Elementi di Bioinformatica Ingegneria Biomedica AA 2013-14 Elementi di Bioinformatica Genomica Introduzione Genomica Genomica (genomics) Riguarda lo studio del genoma degli organismi viventi e,
LA MOLE LA MOLE 2.A PRE-REQUISITI 2.3 FORMULE E COMPOSIZIONE 2.B PRE-TEST
 LA MOLE 2.A PRE-REQUISITI 2.B PRE-TEST 2.C OBIETTIVI 2.1 QUANTO PESA UN ATOMO? 2.1.1 L IDEA DI MASSA RELATIVA 2.1.2 MASSA ATOMICA RELATIVA 2.2.4 ESERCIZI SVOLTI 2.3 FORMULE E COMPOSIZIONE 2.4 DETERMINAZIONE
LA MOLE 2.A PRE-REQUISITI 2.B PRE-TEST 2.C OBIETTIVI 2.1 QUANTO PESA UN ATOMO? 2.1.1 L IDEA DI MASSA RELATIVA 2.1.2 MASSA ATOMICA RELATIVA 2.2.4 ESERCIZI SVOLTI 2.3 FORMULE E COMPOSIZIONE 2.4 DETERMINAZIONE
Metabolismo degli aminoacidi
 Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Raffinamento Cristallografico e Validazione A.A. 2010-2011. Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano
 Raffinamento Cristallografico e Validazione A.A. 2010-2011 Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Cristalli Raffinamento Data collection ρ (x,y,z) Modello
Raffinamento Cristallografico e Validazione A.A. 2010-2011 Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Cristalli Raffinamento Data collection ρ (x,y,z) Modello
Capitolo 4: Le proteine: ossidazione degli amminoacidi, produzione dell urea e biosintesi degli amminoacidi
 Capitolo 4: Le proteine: ossidazione degli amminoacidi, produzione dell urea e biosintesi degli amminoacidi proteine della dieta nucleotidi, ammine biologiche amminoacidi proteine intracellulari ciclo
Capitolo 4: Le proteine: ossidazione degli amminoacidi, produzione dell urea e biosintesi degli amminoacidi proteine della dieta nucleotidi, ammine biologiche amminoacidi proteine intracellulari ciclo
Trasduzione del Segnale da Recettori di Superfice
 Trasduzione del Segnale da Recettori di Superfice MEMBRANA 1. Legame recettore-ligando 2. oligomerizzazione 3. Attivazione TYR-K -Dominio citosolico dei RTK -Reclutamento TYR-K (src, jak, fak, abl) 4.
Trasduzione del Segnale da Recettori di Superfice MEMBRANA 1. Legame recettore-ligando 2. oligomerizzazione 3. Attivazione TYR-K -Dominio citosolico dei RTK -Reclutamento TYR-K (src, jak, fak, abl) 4.
mediante proteasi industriali
 Valorizzazione delle farine disoleate mediante proteasi industriali Alessandra Stefan Unità operativa: CSGI (Consorzio per lo Sviluppo dei Sistemi a Grande Interfase, Firenze) Dipartimento di Farmacia
Valorizzazione delle farine disoleate mediante proteasi industriali Alessandra Stefan Unità operativa: CSGI (Consorzio per lo Sviluppo dei Sistemi a Grande Interfase, Firenze) Dipartimento di Farmacia
Laurea Magistrale in Biologia
 Laurea Magistrale in Biologia Laboratorio del corso di Chimica Fisica Biologica Anno accademico 2009/10 SCOPO Determinazione dei parametri termodinamici associati alla denaturazione termica di una piccola
Laurea Magistrale in Biologia Laboratorio del corso di Chimica Fisica Biologica Anno accademico 2009/10 SCOPO Determinazione dei parametri termodinamici associati alla denaturazione termica di una piccola
dominio strutturale dominio modulo
 Riepilogo 2^lezione DOMINI Si definisce dominio strutturale (o dominio o modulo) di una proteina: un'unità globulare o fibrosa formata da catene polipeptidiche ripiegate in più regioni compatte le quali
Riepilogo 2^lezione DOMINI Si definisce dominio strutturale (o dominio o modulo) di una proteina: un'unità globulare o fibrosa formata da catene polipeptidiche ripiegate in più regioni compatte le quali
SOLUZIONE ESERCIZIO 1.1
 SOLUZIONE ESERCIZIO 1.1 La temperatura di fusione ed il coefficiente di espansione termica di alcuni metalli sono riportati nella tabella e nel diagramma sottostante: Metallo Temperatura di fusione [ C]
SOLUZIONE ESERCIZIO 1.1 La temperatura di fusione ed il coefficiente di espansione termica di alcuni metalli sono riportati nella tabella e nel diagramma sottostante: Metallo Temperatura di fusione [ C]
FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI ALIMENTI PROTEINE ENDOGENE (TURNOVER) BIOSINTESI
 BIOSINTESI") FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI ALIMENTI PROTEINE ENDOGENE (TURNOVER) BIOSINTESI Berg et al., BIOCHIMICA 6/E, Zanichelli editore S.p.A. Copyright 2007 PROTEINE INGERITE CON GLI ALIMENTI
FONTI DI AMMINOACIDI: PROTEINE ASSUNTE CON GLI ALIMENTI PROTEINE ENDOGENE (TURNOVER) BIOSINTESI Berg et al., BIOCHIMICA 6/E, Zanichelli editore S.p.A. Copyright 2007 PROTEINE INGERITE CON GLI ALIMENTI
Il dogma centrale della biologia. molecolare
 Il dogma centrale della biologia Cell molecolare Transcription Translation Ribosome DNA mrna Polypeptide (protein) L informazione per la sintesi delle proteine è contenuta nel DNA. La trascrizione e la
Il dogma centrale della biologia Cell molecolare Transcription Translation Ribosome DNA mrna Polypeptide (protein) L informazione per la sintesi delle proteine è contenuta nel DNA. La trascrizione e la
PROTEINE. sono COMPOSTI ORGANICI QUATERNARI
 PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
DNA - RNA. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi.
 DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
Dipartimento Scientifico-Tecnologico
 ISTITUTO TECNICO STATALE LUIGI STURZO Castellammare di Stabia - NA Anno scolastico 2012-13 Dipartimento Scientifico-Tecnologico CHIMICA, FISICA, SCIENZE E TECNOLOGIE APPLICATE Settore Economico Indirizzi:
ISTITUTO TECNICO STATALE LUIGI STURZO Castellammare di Stabia - NA Anno scolastico 2012-13 Dipartimento Scientifico-Tecnologico CHIMICA, FISICA, SCIENZE E TECNOLOGIE APPLICATE Settore Economico Indirizzi:
FORMAGGIO - DETERMINAZIONE DEL CONTENUTO DI AMMINOACIDI LIBERI
 Decreto n. 6890 ALLEGATO : METODO DI ANALISI FORMAGGIO - DETERMINAZIONE DEL CONTENUTO DI AMMINOACIDI LIBERI 1. SCOPO E CAMPO DI APPLICAZIONE Questo standard descrive la determinazione del contenuto di
Decreto n. 6890 ALLEGATO : METODO DI ANALISI FORMAGGIO - DETERMINAZIONE DEL CONTENUTO DI AMMINOACIDI LIBERI 1. SCOPO E CAMPO DI APPLICAZIONE Questo standard descrive la determinazione del contenuto di
METABOLISMO AMINO ACIDI
 METABOLISMO AMINO ACIDI AMINO ACIDI ESOGENI (ALIMENTAZIONE) POOL AMINO ACIDI PROTEINE DEMOLIZIONE AMINE BIOGENE ORMONI PROTEICI BASI AZOTATE.. SCHELETRO CARBONIOSO NH 3 RESINTESI CICLO KREBS LIPIDI (AA
METABOLISMO AMINO ACIDI AMINO ACIDI ESOGENI (ALIMENTAZIONE) POOL AMINO ACIDI PROTEINE DEMOLIZIONE AMINE BIOGENE ORMONI PROTEICI BASI AZOTATE.. SCHELETRO CARBONIOSO NH 3 RESINTESI CICLO KREBS LIPIDI (AA
DEAMINAZIONE AA - 7. ADP + Pi ATP. glutamina sintetasi. glutaminasi COO CH 2 NH + 4. + _ COO L-glutamato (Glu) COO. L-glutamina (Gln) COO 2 H2O
 COO. L-glutamina (Gln) COO 2 H2O") C DEAMIAZIE AA - 7 ATP ADP Pi C C C C 3 4 glutamina sintetasi C C 3 L-glutamato (Glu) L-glutamina (Gln) C C 4 C C C 3 L-glutamina (Gln) glutaminasi C C 3 L-glutamato (Glu) DEAMIAZIE AA - 8 tessuti extraepatici,
C DEAMIAZIE AA - 7 ATP ADP Pi C C C C 3 4 glutamina sintetasi C C 3 L-glutamato (Glu) L-glutamina (Gln) C C 4 C C C 3 L-glutamina (Gln) glutaminasi C C 3 L-glutamato (Glu) DEAMIAZIE AA - 8 tessuti extraepatici,
ELASTOMERI ELASTOMERI
 ELASTOMERI Gli elastomeri o gomme, sono materiali polimerici le cui dimensioni possono variare enormemente so
ELASTOMERI Gli elastomeri o gomme, sono materiali polimerici le cui dimensioni possono variare enormemente so
Recenti Scoperte Scientifiche in Ottica
 Recenti Scoperte Scientifiche in Ottica Luca Salasnich Dipartimento di Fisica e Astronomia Galileo Galilei, Università di Padova, Italy Padova, 26 Maggio 2014 Secondo Meeting di Ottica e Optometria a Padova
Recenti Scoperte Scientifiche in Ottica Luca Salasnich Dipartimento di Fisica e Astronomia Galileo Galilei, Università di Padova, Italy Padova, 26 Maggio 2014 Secondo Meeting di Ottica e Optometria a Padova
TRASCRIZIONE DEL DNA, TRADUZIONE DELL RNA
 TRASCRIZIONE DEL DNA, TRADUZIONE DELL RNA TRADUZIONE La traduzione e il processo con cui viene sintetizzata un data proteina, attraverso reazioni chimiche di polimerizzazione di amminoacidi, in una
TRASCRIZIONE DEL DNA, TRADUZIONE DELL RNA TRADUZIONE La traduzione e il processo con cui viene sintetizzata un data proteina, attraverso reazioni chimiche di polimerizzazione di amminoacidi, in una