Il catabolismo dei lipidi e l ossidazione degli acidi grassi
|
|
|
- Fausta Ricci
- 6 anni fa
- Visualizzazioni
Transcript
1 Il catabolismo dei lipidi e l ossidazione degli acidi grassi Nella maggior parte degli organismi i lipidi si trovano principalmente nella forma di TRIACILGLICEROLI (trigliceridi) In molte piante una grossa quantità di trigliceridi sono conservati nei semi, nei frutti e nei granuli pollinici (eccezione jojoba che accumula acidi grassi e cere nei semi)
2 I lipidi di deposito vegetali sono anche un importante fonte di grassi alimentari per l uomo e gli altri animali Industria manifatturiera: produzione detergenti, rivestimenti, materie plastiche e lubrificanti.
3 I lipidi rappresentano una enorme riserva energetica. Questo in virtù del fatto che il carbonio nei triacilgliceroli presenta un grado di riduzione maggiore di quello presente nei carboidrati Non bisogna dimenticare la presenza del gruppo carbossilico, altamente ossidato, ma la maggior parte del carbonio è in forma metilenica, quindi altamente ossidata
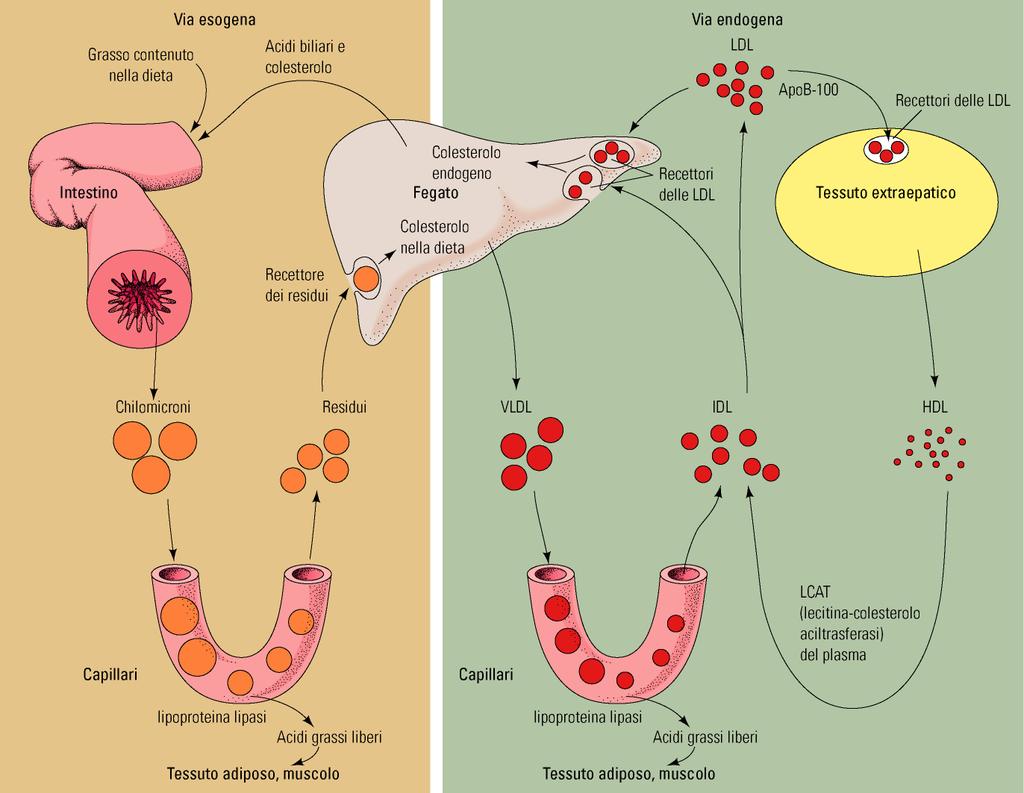
4 Da ciò ne deriva che l ossidazione dei grassi consuma più ossigeno, in rapporto al peso, dell ossidazione dei carboidrati e rilascia una quantità superiore di energia metabolica Negli animali, i grassi provengono da tre vie: - alimentazione - biosintesi ex-novo nel fegato - mobilizzazione del grasso accumulato negli adipociti La mobilizzazione dei lipidi negli animali è operata dalle proteine [(very low density lipoprotein (VLDL), low density lipoprotein (LDL) high density lipoprotein (HDL)]
5
6 Nei semi deidratati, prima della germinazione, vi è una bassissima attività enzimatica, mentre durante l imbibizione si ha un incremento di attività di un gran numero di enzimi tra i quali le lipasi dei trigliceridi I loro substrati sono contenuti in gocce di olio (oleosomi) dentro i semi e gli enzimi agiscono sulla superficie delle gocce, probabilmente con l aiuto di proteine leganti per facilitare il processo


7 Gli enzimi deputati alla scissione dei trigliceridi sono idrolasi più propriamente chiamate LIPASI Le lipasi catalizzano il rilascio di acidi grassi esterificati sulle posizioni 1 e 3 dei trigliceridi per ottenere un monoacilglicerolo Il rilascio di tutti e tre gli acidi grassi è ottenuto attraverso la migrazione dell acido grasso dalla posizione 2 alla posizione 1 e il suo susseguente rilascio


8 Il glicerolo rilasciato viene convertito ad un intermedio della Glicolisi, il DIIDROSSIACETONFOSFATO, in una reazione catalizzata dalla GLICEROLO CHINASI e dalla GLICEROLO FOSFATO DEIDROGENASI
9 La scissione dei fosfolipidi Gli enzimi coinvolti nella scissione e rimodellazione dei fosfolipidi sono chiamati FOSFOLIPASI Nelle piante e nei tessuti animali, un gran numero di fosfolipasi è stato identificato in funzione del sito di attacco sulla molecola del fosfolipide
10

11
12 Il composto risultante che contiene solo un acido grasso, nella posizione 1 o 2, è chiamato lisofosfolipide. Questi enzimi sono coinvolti nella modificazione della composizione in acidi grassi delle membrane fosfolipidiche esistenti La fosfolipasi B sembra essere presente solo nei microrganismi, e poiché agisce sia sulla posizione 1 che 2, è capace di agire sia sui fosfolipidi intatti che sui fosfolipidi liberi La fosfolipasi C ha un ruolo importante nel controllo dell attività enzimatica. Molti enzimi sono regolati da un meccanismo di fosforilazione/defosforilazione calcio dipendente
13 Anche se la fosfolipasi D è stata trovata nei tessuti animali, la fonte principale di quest enzima sono i tessuti vegetali. E un enzima molto attivo coinvolto nella reazione di scambio degli acidi fosfatidici; comunque il suo ruolo nel metabolismo dei fosfolipidi non è ancora del tutto chiaro
14 Catabolismo dei glicolipidi Le piante sono molto ricche di glicolipidi. Il loro catabolismo è particolarmente attivo durante la senescenza ed il susseguente danno ai tessuti La degradazione completa avviene in due stadi: 1) gli acidi grassi sono rimossi dalla acil-idrolasi 2) gli zuccheri residui sono rimossi per azione delle galattosidasi Nei ruminanti una grande quantità dei lipidi ingeriti sono degradati per azione dell acil-idrolasi e della glicosidasi batterica
15 L ossidazione dei grassi avviene mediante la betaossidazione Negli animali questa avviene nel mitocondrio o nel perossioma, mentre nelle piante e in molti funghi avviene solo nel perossisoma In realtà l ossidazione avviene nel perossisoma degli organismi animali solo quando la catena è troppo lunga per potere entrare nel mitocondrio anche se si blocca quando la catena è formata da 8 atomi di C e quindi prosegue nel mitocondrio Come vedremo nel caso della ossidazione perossisomale non si produce ATP ed invece perossido di idrogeno che è convertito ad acqua ed ossigeno dalla catalasi
16 Ossidazione degli acidi grassi Gli acidi grassi possono andare incontro ad una serie di modificazioni ossidative Quantitativamente il percorso biochimico più importante nell ossidazione degli acidi grassi è la OSSIDAZIONE Questo processo avviene in tutti gli organismi ed è la principale via attraverso cui gli acidi grassi sono utilizzati per produrre energia Nella ossidazione, gli acidi grassi sono degradati a composti più piccoli, solitamente acetil-coa, che può poi essere ossidato attraverso il ciclo TCA
17 La conversione delle lunghe catene degli acidi grassi in acetil-coa e la sua successiva ossidazione nel ciclo TCA porta alla produzione di una grossa quantità di coenzimi ridotti quali NADH e FADH 2, che verranno utilizzati per la produzione di ATP Nelle piante e nei funghi, inoltre, la ossidazione può essere utilizzata per ottenere acetil-coa per la sintesi dei carboidrati tramite la via del gliossilato
18 Due altri processi ossidativi, conosciuti come - ossidazione e -ossidazione, sono coinvolti nella modificazione degli acidi grassi -ossidazione: provoca l ossidazione degli acidi grassi sul carbonio in posizione 2 ( rispetto al carbonile) -ossidazione: determina l ossidazione degli acidi grassi sul gruppo metile terminale cioè sul carbonio
19 ossidazione Quasi tutti i tessuti sono in grado di ossidare gli acidi grassi tramite la ossidazione Nella cellula sono presenti due principali siti dove avviene la ossidazione Negli animali è la matrice mitocondriale il luogo in cui gli acidi grassi sono completamente ossidati da acetil-coa. Nel fegato e nei reni riveste invece maggior importanza l ossidazione che avviene nei perossisomi
20 La ossidazione nei perossisomi sembra essere un meccanismo utilizzato principalmente per ridurre la lunghezza delle catene degli acidi grassi, che possono essere successivamente completamente ossidati nella matrice mitocondriale Nei tessuti fogliari delle piante, sembrano i perossisomi più che i mitocondri, gli organelli deputati all ossidazione degli acidi grassi, mentre nei semi i gliossisomi sono i siti principali dell ossidazione degli acidi grassi, in quanto il prodotto dell ossidazione, l acetil-coa, può entrare direttamente nel ciclo del gliossilato
21 ossidazione mitocondriale Gli acidi grassi metabolizzati dalla ossidazione possono derivare sia da fonti esogene (alimenti) che endogene (esempio tessuto adiposo e fegato) Il trasporto degli acidi grassi nel sangue avviene nella forma non esterificata (NEFAs) con la proteina trasportatrice, che normalmente è l ALBUMINA, prodotta dal fegato. Altrimenti vengono trasportati come fosfolipidi e trigliceridi incorporati nelle lipoproteine, principalmente lipoproteine a bassissima densità (VLDL)
22 Gli acidi grassi legati alle lipoproteine sono rilasciati nel sangue attraverso l azione della lipoproteina lipasi, enzima localizzato sulla superficie del lumen delle cellule capillari dell endotelio dei capillari I NEFAs passano dal sangue alle cellule adiacenti sia per diffusione sia per un processo mediato della membrana
23 ATTIVAZIONE DELL ACIDO GRASSO Nel citoplasma, gli acidi grassi devono essere attivati come esteri del CoA prima di essere successivamente metabolizzati La reazione richiede ATP ed è catalizzata da un enzima chiamato ACETIL-CoA SINTASI
24 L acido grasso così attivato sul lato citosolico della membrana mitocondriale esterna può essere trasportato nel mitocondrio oppure essere utilizzati nel citosol per la sintesi dei lipidi di membrana
25 Una serie di enzimi catalizzano questa reazione, ma hanno differente specificità per la lunghezza della catena. I due più importanti nell ossidazione degli acidi grassi sono: acetil-coa sintasi a media catena (C4-C12) acetil-coa sintasi a lunga catena (C10 e più)
26 L acetil-coa sintasi a lunga catena è un enzima legato alla membrana dei perossisomi, nel reticolo endoplasmatico e nella membrana mitocondriale esterna Questa distribuzione permette agli acidi grassi di essere metabolizzati attraverso diversi percorsi metabolici in differenti siti sub-cellulari Ad esempio la riduzione della lunghezza della catena che avviene nei perossisomi, la sintesi dei fosfolipidi e trigliceridi nel reticolo endoplasmatico e la ossidazione nei mitocondri

27 La membrana mitocondriale interna costituisce una barriera agli acetil-coa destinati all ossidazione. Né il CoA né gli esteri del CoA possono attraversarla. Questo è dovuto principalmente alle grandi dimensioni ed alla natura delle cariche presenti nella molecola del CoA Per facilitare il movimento degli acidi grassi attraverso la membrana interna mitocondriale verso la matrice, gli acetil-coa sono convertiti in acil-carnitina ad opera dell enzima carnitina-aciltransferasi I, localizzata nello spazio intermembrane mitocondriale
28 Il complesso Acido grasso-carnitina si forma attraverso la membrana interna tramite una proteina di trasporto, la Acil-carnitina Translocasi, che attua un meccanismo di antiporto (entrata della Carnitina e uscita della Acido grasso-carnitina). Nella matrice mitocondriale l acido grasso viene trasferito dalla Carnitina di nuovo al CoA grazie alla Carnitina aciltransferasi II. Una volta entrati nella matrice mitocondriale come estere del CoA gli acidi grassi subiscono la ossidazione.
29
30 Le reazioni della ossidazione La ossidazione è costituita da una sequenza di quattro reazioni necessarie per rimuovere un unità bicarboniosa della terminazione carbossilica dell acido grasso Questa sequenza di reazioni è ripetuta fino a quando l acido grasso è completamente ossidato
31
32 1) Reazione iniziale di deidrogenazione Il primo passo della ossidazione di un acil-coa è la rimozione di un atomo di idrogeno dai carboni 2 ( ) e 3 ( ) con formazione del 2 TRANS-ENOIL-CoA Si tratta di un ossidazione operata da tre diverse ACETL- CoA DEIDROGENASI che agiscono su acidi grassi a catena di diversa lunghezza (corta, media e lunga) Ciascuna di esse ha tuttavia come gruppo prostetico il FAD. Gli atomi di idrogeno sono accettati dal FAD che viene ridotto a FADH 2 Dal FADH 2 gli elettroni sono traslocati ad una flavoproteina che trasferisce elettroni (ETF) che, a Sua volta li trasferisce alla catena di trasporto mitocondriale
33 2) Idratazione e deidrogenazione Il passo successivo della reazione è l idratazione della molecola. L enzima responsabile di questa reazione è l ENOIL-CoA IDRATASI Questo enzima catalizza l addizione stereospecifica dell acqua sul doppio legame con formazione dell L-3-IDROSSIALCIL-CoA
34 Successivamente si ha una deidrogenazione NAD + - dipendente. L enzima specifico è la 3-IDROSSIACIL-CoA DEIDROGENASI. Questo enzima è relativamente non specifico per la lunghezza della catena, ma è estremamente specifico riguardo allo stereoisomero L del 3-idrossiacil- CoA L enzima catalizza la rimozione di due atomi di idrogeno dal carbonio 1 ed il loro trasferimento al NAD +. Il prodotto della reazione è il 3-CHETACIL-CoA
35 3) Rottura del legame tioestere Il passo finale nel ciclo di reazioni della ossidazione è la rottura tiolitica del legame del 3- acetoacil-coa tra i carboni 2 e 3 ad opera del CoA L enzima che opera la rottura è chiamato CHETOTIOLASI. Un gruppo tiolico essenziale dell enzima (E-SH) attacca il substrato portando alla formazione dell enzima acilato e di acetil-coa come intermedi
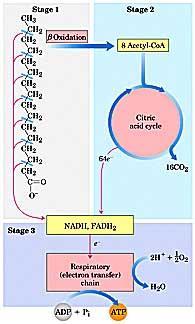
36 Resa energetica della ossidazione Le due riserve più usate dalle cellule per la produzione di Acetil-CoA sono il glucosio e gli acidi grassi. Gli acidi grassi a lunga catena sono composti altamente ridotti e dunque la loro completa ossidazione può condurre alla produzione di un grande quantitativo di ATP La completa ossidazione di una mole di acido palmitico produce 8 moli di acetil-coa che può essere successivamente ossidato attraverso il ciclo TCA Una molecola di ATP è idrolizzata ad AMP e PPi per l attivazione degli acidi grassi prima del trasporto della matrice mitocondriale
37
38 TABELLA 14.1 Resa in ATP dell ossidazione dell acido palmitico (C16) Percorso Conversione netta Cofattori prodotti ossidazione Da acido palmitico a 8 acetil-coa Ciclo TCA 8 Acetil-CoA a CO 2 e acqua Resa in ATP 7 NADH 21 ATP 7 FADH 2 14 ATP 24 NADH 72 ATP 8 FADH 2 16 ATP 8 GTP 8 ATP Totale 131 ATP
39 Una molecola di ATP è idrolizzata ad AMP e PP i per l attivazione degli acidi grassi prima del trasporto nella matrice mitocondriale Questo equivale all utilizzazione di 2 ATP, quindi si ha una sintesi netta di 129 moli di ATP dalla beta ossidazione di una mole di acido palmitico
40 ossidazione degli acidi grassi a catena dispari La ossidazione degli acidi grassi a catena dispari è in parte simile a quella degli acidi grassi a catena pari Il processo procede attraverso la rimozione sequenziale di acetil-coa fino a quando non rimane l intermedio a cinque atomi di cabonio. Questo è quindi convertito in una molecola di acetil-coa e una molecola di propionil-coa
41 Poiché il propionil-coa è un composto a tre atomi di carbonio questo può essere utilizzato per la sintesi del glucosio Comunque, la quantità di acidi grassi a catena dispari trovata nei tessuti animali è solitamente piccola (1-2%) e dunque il contributo alla produzione di glucosio è trascurabile in confronto con altri composti gluconeogenetici Questa è negli animali una delle poche possibilità di conversione di un prodotto dell ossidazione degli acidi grassi in glucosio
42 In questo caso il succinil-coa entra nel ciclo TCA e viene demolito per la produzione di energia
43 La ossidazione degli acidi grassi insaturi Una ossidazione leggermente modificata rispetto a quella tipica è necessaria per l ossidazione degli acidi grassi insaturi L intermedio insaturo prodotto dall Acil-CoA deidrogenasi ha la configurazione 2 Trans, mentre negli acidi grassi insaturi si riscontra un doppio legame in configurazione cis Poiché gli acidi grassi insaturi sono ossidati tramite la rimozione di una unità bicarboniosa dalla coda carbossilica, la posizione dei doppi legami si sposta mano a mano che la catena si accorcia sul carbonio del gruppo carbossilico che è sempre il carbonio 1 Tale situazione ha due importanti conseguenze nella ossidazione degli acidi grassi insaturi
44 Per un acido grasso come l acido oleico che ha un doppio legame cis tra il carbonio 9 e 10, è necessaria una reazione addizionale per fare avvenire la ossidazione Il primo passo è la rimozione dei sei atomi di carbonio (tre molecole di Acetil-CoA) attraverso il solito percorso della ossidazione Il risultato è la produzione di un intermedio enoil- CoA di 12 atomi di carbonio ( 3 cis-dodecanoil- CoA) in cui il doppio legame è nella posizione sbagliata 3 invece che 2, e nella configurazione errata cis anziché trans
45 La posizione e la configurazione del doppio legame vengono modificate grazie all enzima enoil-coa isomerasi Questo enzima converte il doppio legame 3 cis in un doppio legame 2 trans che permette alla ossidazione di continuare regolarmente Attraverso l utilizzo di questi due enzimi addizionali, l enoil-coa isomerasi e la 2,4-dienoil- CoA riduttasi, quasi tutti gli acidi grassi presenti in natura possono essere ossidati tramite la ossidazione
46
47 CONTROLLO DELLA Β- OSSIDAZIONE - Trasporto degli acil-coa nei mitocondri (CAT è inibita da malonil-coa) - Rapporto NAD/NADH - AcetilCoA inibisce la tiolasi
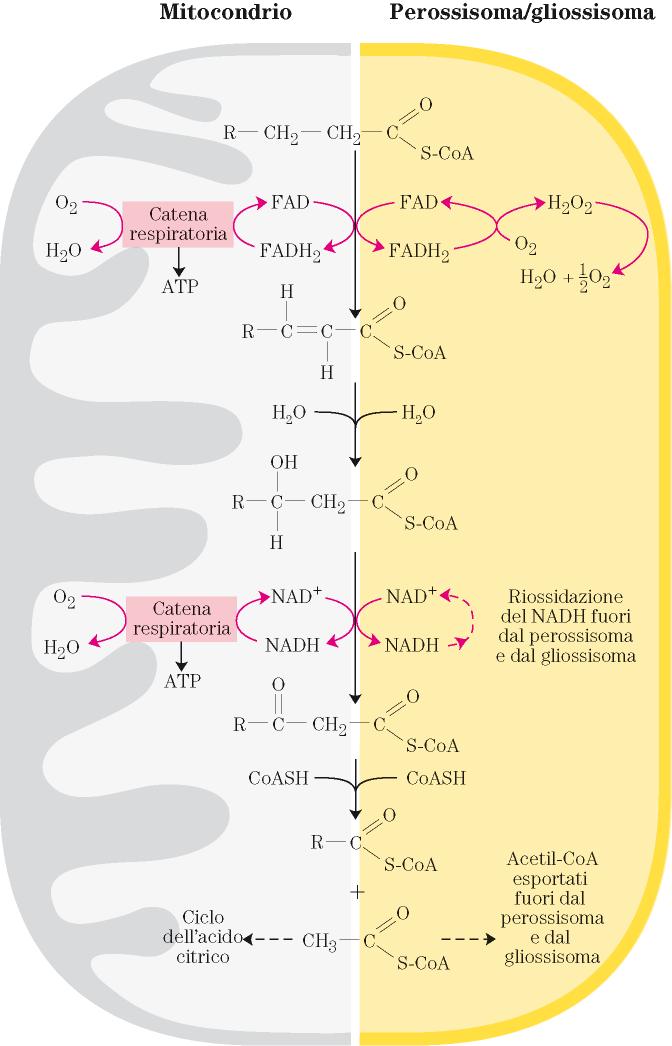
48 La ossidazione degli acidi grassi nei perossisomi e negli gliossisomi differisce da quella dei mitocondri La prima reazione è catalizzata dall ACIL-CoA OSSIDASI e si ha produzione di perossido di idrogeno, che è poi scisso dall azione della catalasi
49
50 Le rimanenti reazioni del ciclo di ossidazione sembrano essere le stesse di quelle dei mitocondri, anche se potrebbero esserci differenze strutturali e funzionali degli enzimi nei diversi organelli subcellulari La membrana dei perossisomi e gliossisomi non costituisce, al contrario della membrana interna mitocondriale, una barriera di permeabilità ai derivati degli acidi grassi collegati al CoA Quindi non è necessario che gli acidi grassi vengano trasportati dalla carnitina prima di essere ossidati
51 Differentemente dai mitocondri, i perossisomi e gliossisomi non contengono una catena di trasporto elettronico capace di utilizzare il NADH prodotto Per assicurare l apporto di NAD + (ossidato), per l ossidazione continua degli acidi grassi, il NADH (ridotto) è trasportato nel citosol in cambio di NAD + La situazione è diversa tra cellule animali e vegetali a)negli ANIMALI l ossidazione perossisomica dà un significativo contributo al processo totale di ossidazione degli acidi grassi: nel fegato si può raggiungere il 50% del totale degli acidi grassi ossidati. Comunque, nei perossisomi, l ossidazione degli acidi grassi è parziale
52 Questi organelli difatti sembrano essere particolarmente importanti nella conversione degli acidi grassi a lunga catena in acidi grassi a media catena che poi vengono trasportati, tramite lo shuttle della carnitina, nei mitocondri, dove il processo di ossidazione si completa L acetil-coa prodotto nei perossisomi viene anche trasportato nella matrice mitocondriale dove può essere ossidato dal ciclo TCA o convertito in corpi chetonici. Il movimento di acetato tra gli organelli subcellulari si basa sempre su meccanismi mediati dalla carnitina
53 Nelle piante, i gliossisomi nei semi ed i perossisomi nelle foglie sono i principali siti di ossidazione. Infatti, si pensa che la ossidazione mitocondriale contribuisca marginalmente all ossidazione degli acidi grassi nelle piante, mentre è sicuro che sia i gliossisomi che i perossisomi sono capaci di ossidare completamente gli acidi grassi In alcuni semi germinanti, la quantità di acidi grassi mobilitata dai grassi di riserva e ossidata attraverso i gliossisomi è enorme, specialmente nelle oleaginose. Questi speciali organelli contengono sia enzimi necessari per ossidare gli acidi in acetil-coa, sia gli enzimi del ciclo del gliossilato, l isocitrato liasi e la malato sintasi che permettono alle piante di convertire l acetil-coa in glucosio
54 -ossidazione Come suggerisce lo stesso nome, questo processo determina l ossidazione degli acidi grassi a livello del carbonio (carbonio 2). Si può avere la rimozione di un singolo atomo di carbonio dal gruppo carbossilico o la produzione di acidi grassi -idrossilici
55 (a) Rimozione del gruppo COOH (b) Produzione di un -idrossi acido grasso
56 Il significato dell -ossidazione nei tessuti animali è ancora sconosciuto. Ci sono tre campi in cui potrebbe giocare un ruolo: Primo, potrebbe essere un meccanismo attraverso il quale sono sintetizzati gli acidi grassi a catena dispari, ad esempio la rimozione di un atomo di carbonio dal gruppo carbossilico di un acido grasso a catena pari Secondo, potrebbe essere coinvolta nella sintesi degli acidi grassi -idrossilici trovati in certi tessuti, in particolare nei tessuti cerebrali dove gli sfingolipidi contengono una grande quantità di questo tipo di acidi grassi Terzo, potrebbe agire di concerto con la -ossidazione per facilitare l ossidazione di acidi grassi che hanno caratteristiche strutturali che ne impedirebbero l ossidazione tramite la sola -ossidazione
57 Anche nelle piante l importanza della - ossidazione non è chiara. Si è tuttavia dimostrato che, con l eccezione dei semi germinanti dove la -ossidazione è estremamente attiva, l ossidazione potrebbe essere il percorso più importante per l ossidazione degli acidi grassi Oltre al suo ruolo nella degradazione degli acidi grassi, l -ossidazione è certamente coinvolta nella produzione degli alcooli a lunga catena e degli idrocarburi trovati nella cutina e suberina che compongono la cuticola
58 -ossidazione In questo processo, gli acidi grassi vanno incontro all ossidazione del carbonio o metilico trasformando acidi dicarbossilici e acidi grassi - idrossilici Negli animali, l enzima responsabile sembra essere un ossidasi con funzioni miste, associata al reticolo endoplasmatico e probabilmente coinvolgente un citocromo specializzato (P 450 ). Nelle piante il coinvolgimento del citocromo è dubbio
59 La -ossidazione può essere un passo essenziale nell ossidazione degli acidi grassi là dove il gruppo carbossilico sia indisponibile per la - ossidazione; infatti, la produzione di un gruppo carbossile dal carbonio metilico di un acido grasso permette al processo della -ossidazione di partire dalla parte opposta della molecola La produzione di un acido grasso -idrossilico può avere un ruolo anche nella formazione della cutina e della suberina
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di
 IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia.
 La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
Il catabolismo degli acidi grassi
 Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico 2014-15
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Capitolo 6 La respirazione cellulare
 Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico
 Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Attenzione : lunedì 29 aprile NON ci sarà lezione
 Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Idratazione (introduciamo un gruppo OH sul Cβ) per mezzo della enoil-coa idratasi
 per mezzo della enoil-coa idratasi") β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
LIPIDI COMPLESSI E LIPOPROTEINE
 LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
Seminario. Ciclo di Krebs 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A
 Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine
 Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
Cap.19. Ciclo di Krebs. o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico
 Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
INSULINA RECERE REC INS T U T L O INA RE OR
 INSULINA RECERE REC INSULINA TTORE OR Lipidi Sono un gruppo eterogeneo di composti con una caratteristica comune: l isolubilità in acqua Lipidi di riserva (triacilgliceroli o trigliceridi e cere) Lipidi
INSULINA RECERE REC INSULINA TTORE OR Lipidi Sono un gruppo eterogeneo di composti con una caratteristica comune: l isolubilità in acqua Lipidi di riserva (triacilgliceroli o trigliceridi e cere) Lipidi
Formazione dei chilomicroni
 Formazione dei chilomicroni I triacilgliceroli forniscono il combustibile per i lunghi voli migratori del Piviere minore. I triacilgliceroli sono riserve di energia molto concentrate essendo ridotti ed
Formazione dei chilomicroni I triacilgliceroli forniscono il combustibile per i lunghi voli migratori del Piviere minore. I triacilgliceroli sono riserve di energia molto concentrate essendo ridotti ed
Biosintesi dei carboidrati
 Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
INTRODUZIONE AL METABOLISMO. dal gr. metabolè = trasformazione
 INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
Come le cellule traggono energia dal cibo: produzione di ATP
 Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la
 Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
METABOLISMO DEI LIPIDI
 METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
Energia e metabolismi energetici
 Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
BIOCHIMICA e BIOTECNOLOGIE degli ALIMENTI
 Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
L ossidazione completa del glucosio da parte dell O 2. può essere suddivisa in due semi-reazioni
 L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
Indice generale 1. Introduzione alla biochimica 2. Acqua, ph, acidi, basi, sali, tamponi
 Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
3. Citologia i. Strutture cellulari comuni tra cellule animali e vegetali
 Strutture cellulari comuni tra cellule animali e vegetali: CITOPLASMA CITOSCHELETRO RIBOSOMI RETICOLO ENDOPLASMATICO APPARATO DEL GOLGI MITOCONDRI NUCLEO PEROSSISOMI CITOPLASMA materiale gelatinoso incolore
Strutture cellulari comuni tra cellule animali e vegetali: CITOPLASMA CITOSCHELETRO RIBOSOMI RETICOLO ENDOPLASMATICO APPARATO DEL GOLGI MITOCONDRI NUCLEO PEROSSISOMI CITOPLASMA materiale gelatinoso incolore
6 H 2. con G=-686 kcal/mole di H 12 O 6 O + 6 CO O 2. glucosio La respirazione avviene in tre stadi principali; ognuno di questi
 La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
DESTINI DEL PIRUVATO
 DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore)
") Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale
 Stechiometria della reazione finale") Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
 1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
Connessioni. Le cellule cardiache sono unite tra loro da parti più ispessite del sarcolemma dette: Gap - junction
 Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
2 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA
 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
Fegato e metabolismo lipidico
 Fegato e metabolismo lipidico LIPIDI : Acidi grassi liberi o esterificati ad Acil gliceroli ( mono, di, tri) Fosfolipidi e colesterolo FONTI : Grassi della dieta Depositati sotto forma di goccioline (tessuto
Fegato e metabolismo lipidico LIPIDI : Acidi grassi liberi o esterificati ad Acil gliceroli ( mono, di, tri) Fosfolipidi e colesterolo FONTI : Grassi della dieta Depositati sotto forma di goccioline (tessuto
Ossidazione del glucosio
 Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
INTRODUZIONE AL METABOLISMO
 INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE
 BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
Catabolismo dei lipidi
 Catabolismo dei lipidi Il 90% dei lipidi è costituito da trigliceridi o triacilgliceroli o grassi neutri! 5 25 % peso corporeo dei mammiferi! E la fonte primaria di riserva di energia degli animali! Nel
Catabolismo dei lipidi Il 90% dei lipidi è costituito da trigliceridi o triacilgliceroli o grassi neutri! 5 25 % peso corporeo dei mammiferi! E la fonte primaria di riserva di energia degli animali! Nel
 http://digilander.libero.it/glampis64 Idrogeno, ossigeno, carbonio e azoto costituiscono il 99% delle cellule. I composti del carbonio sono chiamati composti organici o molecole organiche. I composti organici
http://digilander.libero.it/glampis64 Idrogeno, ossigeno, carbonio e azoto costituiscono il 99% delle cellule. I composti del carbonio sono chiamati composti organici o molecole organiche. I composti organici
prodotto finale della glicolisi
 Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Anatomia biochimica di un mitocondrio
 Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Ruolo del fegato nella nutrizione
 Ruolo del fegato nella nutrizione Centro di distribuzione dei nutrienti per tutti gli altri organi Annulla le fluttuazioni del metabolismo determinate dall assunzione intermittente del cibo Funge da deposito
Ruolo del fegato nella nutrizione Centro di distribuzione dei nutrienti per tutti gli altri organi Annulla le fluttuazioni del metabolismo determinate dall assunzione intermittente del cibo Funge da deposito
CLASSIFICAZIONE ENZIMI
 CLASSIFICAZIONE ENZIMI nome raccomandato nome sistematico reazione catalizzata Classe di enzima Reazione catalizzata 1. Ossidoreduttasi Ossido-riduzione 2. Transferasi Trasferimento di gruppi funzionali
CLASSIFICAZIONE ENZIMI nome raccomandato nome sistematico reazione catalizzata Classe di enzima Reazione catalizzata 1. Ossidoreduttasi Ossido-riduzione 2. Transferasi Trasferimento di gruppi funzionali
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO)
") RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
Prof. Maria Nicola GADALETA DISPENSA N. 23
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI
 LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
scaricato da www.sunhope.it
 CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
Laboratorio Fitness & Wellness
 Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
scaricato da
 LIPIDI LIPIDI A differenza delle altre biomolecole, presentano: piccole dimensioni grande varietà strutturale I lipidi sono eccellenti isolanti elettrici: lipidi di membrana termici: lipidi di riserva
LIPIDI LIPIDI A differenza delle altre biomolecole, presentano: piccole dimensioni grande varietà strutturale I lipidi sono eccellenti isolanti elettrici: lipidi di membrana termici: lipidi di riserva
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
I LIPIDI (GRASSI) Cosa sono
 Cosa sono") I LIPIDI (GRASSI) Cosa sono Comunemente chiamati "grassi", i lipidi comprendono una grande varietà di molecole, accomunate dalla caratteristica di essere insolubili in acqua. I lipidi più importanti dal
I LIPIDI (GRASSI) Cosa sono Comunemente chiamati "grassi", i lipidi comprendono una grande varietà di molecole, accomunate dalla caratteristica di essere insolubili in acqua. I lipidi più importanti dal
ENZIMI. Un enzima è un catalizzatore (acceleratore) di reazioni biologiche.
 di reazioni biologiche.") ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
Il metabolismo cellulare
 Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Prof. Maria Nicola GADALETA FOSFORILAZIONE OSSIDATIVA 4
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
Il FEGATO Cardarera, cap.3
 Il FEGATO Cardarera, cap.3 2.5% della massa corporea 1,6-1.8 Kg nel maschio 1.3-1.4 Kg nella femmina Tramite la vena porta, è il primo organo a ricevere il sangue arricchito di nutrienti assorbiti dall
Il FEGATO Cardarera, cap.3 2.5% della massa corporea 1,6-1.8 Kg nel maschio 1.3-1.4 Kg nella femmina Tramite la vena porta, è il primo organo a ricevere il sangue arricchito di nutrienti assorbiti dall
MODULO 2 UNITÀ 2.3 I LIPIDI
 MODULO 2 UNITÀ 2.3 I LIPIDI Pag. 116 Aspetti generali I lipidi, anche detti grassi, sono composti ternari C, H, O (possono contenere anche P, N) Caratteristiche: sono untuosi al tatto sono insolubili in
MODULO 2 UNITÀ 2.3 I LIPIDI Pag. 116 Aspetti generali I lipidi, anche detti grassi, sono composti ternari C, H, O (possono contenere anche P, N) Caratteristiche: sono untuosi al tatto sono insolubili in
G i l i A ut u otro r fi G i l i E t E ero r tro r fi I F ototro r fi I C he h mo m tro r fi i us u a s no C e h moete t ro r tr t o r fi
 Organismi differenti utilizzano differenti modalità per ottenere carbonio ed energia: carbonio: Gli Autotrofi usano CO 2 Gli Eterotrofi usano carbonio organico energia: I Fototrofi usano la luce I Chemotrofi
Organismi differenti utilizzano differenti modalità per ottenere carbonio ed energia: carbonio: Gli Autotrofi usano CO 2 Gli Eterotrofi usano carbonio organico energia: I Fototrofi usano la luce I Chemotrofi
Il metabolismo microbico
 Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
Utilizzo del glucosio: la glicolisi
 Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
DESTINI METABOLICI DEL PIRUVATO
 DETINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DETINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa
 e Fosforilazione ossidativa") Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
scaricato da 1
 scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
Βeta ossidazione degli acidi grassi BETA OSSIDAZIONE DEGLI ACIDI GRASSI 13/12/2013 SEMINARIO. Acidi grassi (2)
") Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI
 METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO
 IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO ( Introduzione Metabolismo energetico Funzione Il metabolismo energetico è un attività coordinata cui partecipano
IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO ( Introduzione Metabolismo energetico Funzione Il metabolismo energetico è un attività coordinata cui partecipano
FOSFORILAZIONE OSSIDATIVA
 FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
Lipidi: funzioni. Strutturale. Riserva energetica. Segnale.
 Lipidi: funzioni. Strutturale. Riserva energetica. Segnale. Lipidi:classificazione. Saponificabili Non saponificabili Semplici Complessi Prostaglandine Steroidi Cere Trigliceridi Fosfogliceridi Sfingolipidi
Lipidi: funzioni. Strutturale. Riserva energetica. Segnale. Lipidi:classificazione. Saponificabili Non saponificabili Semplici Complessi Prostaglandine Steroidi Cere Trigliceridi Fosfogliceridi Sfingolipidi
FADH ADP + Pi ATP...29 AG ' = -30,5 kj/mol...29 Resa di ATP per l ossidazione completa del glucosio...31
 FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA FOTOSINTESI INDICE Organismi autotrofi ed eterotrofi Confronto tra fotosintesi e respirazione Reazioni endoergoniche ed esoergoniche Reazioni accoppiate Fase luce-dipendente
Helena Curtis N. Sue Barnes LA FOTOSINTESI INDICE Organismi autotrofi ed eterotrofi Confronto tra fotosintesi e respirazione Reazioni endoergoniche ed esoergoniche Reazioni accoppiate Fase luce-dipendente
Immagini e concetti della biologia
 Sylvia S. Mader Immagini e concetti della biologia 2 A3 Le molecole biologiche 3 Il carbonio è l elemento di base delle biomolecole Una cellula batterica può contenere fino a 5000 tipi diversi di composti
Sylvia S. Mader Immagini e concetti della biologia 2 A3 Le molecole biologiche 3 Il carbonio è l elemento di base delle biomolecole Una cellula batterica può contenere fino a 5000 tipi diversi di composti
Regolazione ormonale del metabolismo energetico. Lezione del 11 marzo 2014
 Regolazione ormonale del metabolismo energetico Lezione del 11 marzo 2014 Effetti fisiologici e metabolici dell adrenalina: preparazione all azione EFFETTO IMMEDIATO EFFETTO COMPLESSIVO Effetto fisiologico
Regolazione ormonale del metabolismo energetico Lezione del 11 marzo 2014 Effetti fisiologici e metabolici dell adrenalina: preparazione all azione EFFETTO IMMEDIATO EFFETTO COMPLESSIVO Effetto fisiologico
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Metabolismo glucidico
 Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
NADH FADH 2 (trasportatori ridotti di elettroni) Catena respiratoria (trasferimento degli
 Catena respiratoria (trasferimento degli") NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
LE MOLECOLE BIOLOGICHE
 LE MOLECOLE BIOLOGICHE Le cellule contengono quattro famiglie principali di molecole organiche Zuccheri (monosaccaridi) - forniscono una fonte di energia - subunità dei polisaccaridi Amminoacidi - subunità
LE MOLECOLE BIOLOGICHE Le cellule contengono quattro famiglie principali di molecole organiche Zuccheri (monosaccaridi) - forniscono una fonte di energia - subunità dei polisaccaridi Amminoacidi - subunità
Il ciclo di Krebs e la fosforilazione ossidativa
 Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
MEMBRANE. struttura: fosfolipidi e proteine
 MEMBRANE struttura: fosfolipidi e proteine I lipidi sono sostanze di origine biologica insolubili in acqua. Vi fanno parte: trigliceridi, fosfolipidi, colesterolo, sfingolipidi, alcoli alifatici, cere,
MEMBRANE struttura: fosfolipidi e proteine I lipidi sono sostanze di origine biologica insolubili in acqua. Vi fanno parte: trigliceridi, fosfolipidi, colesterolo, sfingolipidi, alcoli alifatici, cere,
Fonte diretta di Energia è l ATP.
 Una cellula compie tre tipi di lavoro: -Lavoro meccanico: movimenti muscolari.. -Lavoro di trasporto: trasporto contro gradiente -Lavoro chimico: assemblaggio di polimeri da monomeri Fonte diretta di Energia
Una cellula compie tre tipi di lavoro: -Lavoro meccanico: movimenti muscolari.. -Lavoro di trasporto: trasporto contro gradiente -Lavoro chimico: assemblaggio di polimeri da monomeri Fonte diretta di Energia
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Lipidi. 1.Trigliceridi 2.Fosfogliceridi 3.Sfingolipidi 4.Prostaglandine 5.Terpeni 6.Steroidi
 Lipidi Lipidi Sono sostanze solubili in composti organici non polari, e possono essere suddivise in 6 distinte tipologie sulla base della loro struttura e funzione: 1.Trigliceridi 2.Fosfogliceridi 3.Sfingolipidi
Lipidi Lipidi Sono sostanze solubili in composti organici non polari, e possono essere suddivise in 6 distinte tipologie sulla base della loro struttura e funzione: 1.Trigliceridi 2.Fosfogliceridi 3.Sfingolipidi
IL METABOLISMO ENERGETICO
 IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
La quantità di glu presente nei liquidi extracellulari è di circa 20g, 80kcal.
 I principali nutrienti metabolici sono il glucosio e gli acidi grassi. In circostanze normali il glucosio è l unico nutriente che possa essere utilizzato dal cervello. E anche utilizzato preferenzialmente
I principali nutrienti metabolici sono il glucosio e gli acidi grassi. In circostanze normali il glucosio è l unico nutriente che possa essere utilizzato dal cervello. E anche utilizzato preferenzialmente
Apparato digerente. Quantità Secrezione. Composizione. giornaliera (L) Saliva Amilasi, bicarbonato. Succo gastrico Succo pancreatico
 Saliva Amilasi, bicarbonato. Succo gastrico Succo pancreatico") Apparato digerente Quantità Secrezione giornaliera (L) ph Composizione Saliva 1 6.5 Amilasi, bicarbonato Succo gastrico Succo pancreatico 1-3 1.5 Pepsinogeno, HCl, fattore intrinseco 1 7-8 Tripsinogeno,
Apparato digerente Quantità Secrezione giornaliera (L) ph Composizione Saliva 1 6.5 Amilasi, bicarbonato Succo gastrico Succo pancreatico 1-3 1.5 Pepsinogeno, HCl, fattore intrinseco 1 7-8 Tripsinogeno,
I lipidi. Semplici Complessi
 Lipidi I lipidi I lipidi sono sostanze non polari (idrofobiche) solubili in solven8 organici. Mol8 dei lipidi che compongono le membrane biologiche sono anfipa8ci, hanno una parte polare ed una apolare
Lipidi I lipidi I lipidi sono sostanze non polari (idrofobiche) solubili in solven8 organici. Mol8 dei lipidi che compongono le membrane biologiche sono anfipa8ci, hanno una parte polare ed una apolare
Metabolismo glucidico
 Metabolismo glucidico Glicogeno glicogenolisi Riboso-5P (NADPH) Colesterolo glicogenosintesi shunt pentoso fosfati Glucoso Glucoso-6P glicolisi Piruvato gluconeogenesi CO 2 -ossidazione Trigliceridi Acidi
Metabolismo glucidico Glicogeno glicogenolisi Riboso-5P (NADPH) Colesterolo glicogenosintesi shunt pentoso fosfati Glucoso Glucoso-6P glicolisi Piruvato gluconeogenesi CO 2 -ossidazione Trigliceridi Acidi
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula