LA GENETICA QUANTITATIVA
|
|
|
- Cesare Romagnoli
- 8 anni fa
- Visualizzazioni
Transcript
1 Genetica 5

2 LA GENETICA QUANTITATIVA

3 Caratteri quantitativi vs. Caratteri qualitativi

4 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005


5 controversia Mendel/cararatteri continui Mendel aveva lavorato su caratteri discontinui, per cui molti dubitavano che le sue leggi valessero anche per i caratteri quantitativi, quali l altezza, che si sapevano essere ereditari ma non se ne conoscevano i meccanismi di trasmissione. Nel 1918 Fisher provò che l ereditarietà delle caratteristiche quantitative poteva realmente venire spiegata attraverso gli effetti cumulativi di molti geni, ciascuno dei quali seguiva le leggi di Mendel


6 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

7 Ereditarietà e variabilità dei caratteri quantitativi > caratteri che variano in modo continuo nella popolazione e che per tale motivo possono essere misurati es. lunghezza della spiga, peso del seme, dimensione corporea di un animale, altezza di un individuo, ecc. > caratteri poligenici poichè sono controllati da molti geni con effetto limitato ma aventi azione uguale e cumulativa sul valore fenotipico; > caratteri la cui manifestazione fenotipica può essere influenzata da fattori ambientali.

8 Caratteri multifattoriali: poligeni e ambiente a) Carattere Poligenico: per l espressione di un determinato carattere continuo (varietà di fenotipi) concorrono soltanto più geni Ma non solo genotipi multipli. b) Carattere Multifattoriale: per l espressione di un determinato carattere continuo concorrono più geni e fattori ambientali
 Carattere Multifattoriale: per l espressione di un determinato carattere")
9 Fattori multigenici + norma di reazione (NdR: gamma di fenotipi prodotti da un genotipo in vari ambienti) Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

10 Analisi dei caratteri quantitativi a) Dividere le variazioni osservate nel carattere nelle due componenti genetica ambientale b) Utilizzare la componente genetica per la previsione dei fenotipi della progenie

11 Campioni e popolazioni Affinchè il campione descriva la popolazione, esso deve essere: Rappresentativo Sufficientemente ampio Rappresentativo: una squadra di pallacanestro non rappresenta tutte le altezze di una popolazione Ampio: se misurassimo altezza di soli 3 studenti dell ateneo di VR e questi causalmente fossero tutti e 3 bassi, sottostimeremmo l altezza media della popolazione studentesca di VR

12 Metodi statistici per l analisi delle caratteristiche quantitative All interno di un gruppo, la variabilità fenotipica può essere rappresentata per mezzo di una distribuzione di frequenza: grafico di frequenze relative ai diversi fenotipi Asse X riporta le classi fenotipiche. L asse Y il numero o la percentuale. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

13 Distribuzione normale, asimmetrica e bimodale Quando la curva è simmetrica, la distribuzione è normale. B) asimmetrica, C) bimodale Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
 asimmetrica, C) bimodale Pierce, GENETICA,")
14 La media La media fornisce informazioni sul centro di una distribuzione Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 x = n x i Misuriamo altezza ragazzi di 10 e di 18 anni. Entrambe le distribuzioni sono normali, ma centrate su valori di statura differenti. FORMULA: la media si calcola sommando le singole misurazioni e dividendo per il numero di misurazioni


15 La varianza (s 2 ) La varianza è definita come la deviazione quadratica media dalla media. Essa indica il grado di variabilità di un gruppo di fenotipi (misurazioni). Quanto maggiore è la varianza, tanto più dispersi sono i valori di una distribuzione intorno alla media Per calcolare la varianza si sottrae la media da ogni misurazione, portando al quadrato il valore ottenuto. Si sommano poi i valori e il risultato lo si divide per il numero di misurazioni di partenza meno 1 S 2 = (x i x) 2 n - 1


16 La deviazione standard (s) La deviazione standard è la radice quadrata della varianza. Essa misura la dispersione dei dati intorno al valore atteso Si esprime nelle stesse unità della misura originale, pertanto descrive la variabilità di una misura s = s 2 La media più o meno una deviazione standard comprende il 66% circa delle misurazioni relative ad una distribuzione normale. Di conseguenza, solo l 1% di una popolazione con distribuzione normale rimane fuori dall intervallo media +- 3s Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

17 Coefficiente di correlazione (r) Il coefficiente di correlazione stabilisce la forza dell associazione fra due caratteristiche. Es. altezza (x) e peso di una persona (y) Accade sovente che due o più caratteri varino contemporaneamente. Per esempio, negli allevamenti di polli sono importanti sia il numero sia la dimensione delle uova prodotte. Queste due caratteristiche non sono tra di loro indipendenti, bensi legate da una correlazione inversa

18 Coefficiente di correlazione (r) r = cov xy s x s y Il coeff. di correlazione si ottiene dividendo la covarianza di x e y per il prodotto delle deviazioni standard di x e y. Esso spazia da +1 a -1. Un valore positivo indica l esistenza di correlazione positiva fra le due variabili cov xy = (x i x) (y i y) n - 1 La covarianza è un indice che misura la "contemporaneità" della variazione (in termini lineari) di due variabili casuali. In pratica, la covarianza di due variabili aleatorie X e Y è il valore atteso dei prodotti delle loro distanze dalla media Essa può assumere sia valori positivi che negativi. Nel caso di valori positivi indica che al crescere di una caratteristica statisticamente cresce anche l'altra, nel caso di valori negativi accade il contrario
 di due variabili casuali.")

19 altezza e peso numero uova e peso uova altezza e intelligenza Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

20 r puo essere calcolato anche per una sola variabile misurata per coppie di individui Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

21 Coefficiente di regressione (b) Indica l entità dell aumento di y all aumentare di x. Pertanto, b consente di prevedere il valore di una variabile, dato il valore dell altra b = cov xy s x 2 la curva di regressione è ( y = a + bx ) a = intercetta di y, ovvero il valore di y quando x=0, b = pendenza della curva, ovvero l aumento medio di y all aumentare di x Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

22 Il coefficiente di regressione consente di prevedere determinate caratteristiche della progenie generata da una data unione anche senza conoscere i genotipi che codificano per tale carattere Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
23 Influenza dell ambiente sulla variabilità dei caratteri quantitativi Wilhelm Johannsen ( ): primo studioso a mettere in evidenza l azione congiunta dei fattori genetici e dei fattori ambientali nell eredità dei caratteri quantitativi egli realizzò una serie di esperimenti utilizzando semi di fagiolo della varietà commerciale Princess, sapendo che la specie scelta è strettamente autogama e che ciascun seme era da ritenersi omozigote a tutti i loci
24 Influenza dell ambiente sulla variabilità dei caratteri quantitativi la variabilità fenotipica di una carattere quantitativo può avere due componenti: una genetica ed un altra ambientale; la selezione è efficace solo in presenza di variabilità genetica; la variabilità che si osserva entro linee pure è dovuta unicamente all ambiente; la selezione entro linea pura è del tutto inefficace.
25 Esperimenti di W. Johannsen in fagiolo: Influenza dell ambiente sull espressione dei caratteri quantitativi Johannsen valutando il peso dei semi di fagiolo è stato il primo a distinguere la variabilità genetica dalla variabilità ambientale e a dimostrare che la selezione può operare solo sulla variabilità genetica. Le 19 linee estratte dalla varietà commerciale erano infatti differenti: le differenze tra le medie delle linee in termini di peso del seme dovevano essere di natura genetica. La selezione entro linee risultava invece inefficace dimostrando che la variabilita del peso del seme entro linea dipendeva soltanto da fattori ambientali.
26 Influenza dei fattori genetici sulla variabilità dei caratteri quantitativi Herman Nilsson-Ehle (1908): è stato il primo ricercatore a trovare un modello naturale in grado di spiegare l eredità dei caratteri quantitativi. egli studiò il colore della cariosside in frumento e formulò l ipotesi che più coppie alleliche segreganti in maniera indipendente, ereditate in assenza di dominanza ed aventi azione uguale e cumulativa (additiva) potessero spiegare i risultati relativi al grado di espressione del carattere nella generazione F 2.
27 Eredità del colore della cariosside in frumento P Colore delle cariossidi A A B B x A 1 A 1 B 1 B1 Bianco Rosso molto scuro Grado di colore 0 4 F 1 AB (¼) A 1 B (¼) AB 1 (¼) A 1 B 1 (¼) A A 1 B B1 Rosso intermedio 2 AB (¼) A 1 B (¼) AB 1 (¼) A 1 B 1 (¼) AABB AA 1 BB AABB 1 AA 1 BB 1 AA 1 BB A 1 A 1 BB AA 1 BB 1 A 1 A 1 BB 1 AABB 1 AA 1 BB 1 AAB 1 B 1 AA 1 B 1 B 1 AA 1 BB 1 A 1 A 1 BB 1 AA 1 B 1 B 1 A 1 A 1 B 1 B 1 F 2 Rapporto 15:1 piante con cariossidi colorate (ma di intensità variabile: chiaro, medio, scuro e molto scuro) e piante con cariossidi bianche
28
29 Influenza dei fattori genetici sulla variabilità dei caratteri quantitativi Se due coppie alleliche vengano ereditate in maniera indipendente, in assenza di dominanza e gli alleli plus hanno azione uguale e cumulativa nel dare la colorazione: l intensità del colore è data dal numero di alleli A 1 e B 1 che nel genotipo delle diverse piante F 2 agiscono nello stesso senso in maniera cumulativa AB 1 A 1 B AB A 1 B 1 AB A 1 B AB 1 A 1 B 1 Azione di fattori A 1 B 1 per il colore e di alleli A e B per il non colorato geni plus geni minus Alleli minus Alleli che esercitano un effetto minimo o che sono ininfluenti sulla manifestazione fenotipica di un carattere quantitativo. Alleli plus Alleli in grado di contribuire alla manifestazione di un carattere quantitativo agendo nello stesso senso in maniera cumulativa e con effetto sul fenotipo superiore alla media.
30 Assunzioni dell ipotesi multigenica di Nilsson-Ehle: in nessuno dei loci uno degli alleli presenta dominanza sull altro, piuttosto risulta coinvolta una serie di alleli con effetto additivo; ogni allele plus agisce nello stesso senso in maniera cumulativa ed ha uguale effetto sul fenotipo; gli alleli minus non contribuiscono (o contribuiscono in maniera minima) al fenotipo; non esiste interazione genica (epistasi) tra loci differenti di una serie; i loci non sono associati, cioè gli alleli segregano in maniera indipendente. Questo lavoro ha dimostrato che la differenza fra eredità di caratteri continui e discontinui risiede nel numero di loci che determinano quel carattere
31 Determinazione numero di geni per caratteristica poligenica Vengono incrociati due individui omozigoti per alleli differenti (A1 e A) di un singolo locus (A) A 1 A 1 x AA A 1 A Gli individui della risultante F1 sono incrociati fra di loro A 1 A x A 1 A ¼ della F2 risulterà omozigote come i parentali. Se ci sono 2 loci coinvolti, ¼ x ¼ = 1/16 A 1 A 1 A 1 A AA 1 AA ¼ 2/4 ¼ Pertanto, in F2 il numero di individui simili a ciascuno dei parentali (n indica numero di loci) e : 1 4 n se trovo: 1/4 individui, significa che c e un locus 1/16 individui, significa che ci sono 2 loci 1/256 individui, significa che ci sono 4 loci Questo metodo assume che tutti i geni che influenzano la caratteristica siano additivi, presentino effetti equivalenti e non siano associati.
32 Determinazione numero di geni per caratteristica poligenica Edward East accoppiò due varietà di tabacco omozigoti per la lunghezza del fiore: 40,5 mm e 93,3 mm, rispettivamente. Egli trovò che nella F 1 la lunghezza del fiore era intermedia, con una varianza simile a quella riscontrata nei parentali. East accoppiò poi fra di loro gli individui F 1. Nella F 2 la lunghezza media del fiore era simile a quella riscontrata nella F 1, ma la varianza era molto superiore, ad indicare la presenza di differenti genotipi. Dato che nessuna delle 444 piante F 2 mostrava lunghezza del fiore analoga ai parentali, il carattere doveva essere codificato da più di 4 loci Successivamente, East selezionò alcune piante F 2 e le incrociò: nella F 3 l estensione del fiore dipendeva dalla lunghezza del fiore nei parentali. Questo indicava l esistenza di differenze sul piano genetico nell ambito della F 2. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

33 Localizzazione dei geni che influiscono sulle caratteristiche quantitative QTL (quantitative trait loci) sono i geni che controllano le caratteristiche poligeniche Si identificano marcatori molecolari che segregano nella popolazione assieme ad una porzione del carattere quantitativo Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005


34 Quantitative Trait Locus (QTL) Un QTL è una regione di DNA associata ad un particolare carattere quantitativo. Il QTL è strettamente associato ai geni che determinano il carattere fenotipico in questione o partecipa nella sua determinazione. Normalmente infatti un carattere quantitativo è determinato dalla somma dell'azione di più geni (un fenomeno detto additività). Di conseguenza più QTL, che possono trovarsi anche su diversi cromosomi, sono associati ad un singolo carattere. Il numero di QTL coinvolti in un carattere fornisce informazioni sull'architettura genetica del carattere; per esempio indica se l'altezza di una pianta è determinata da molti geni, l'effetto di ognuno dei quali è di portata limitata, oppure invece da pochi geni con un effetto più marcato.
35 Ereditabilità di un carattere (H 2 ) Frazione della variazione fenotipica causata dalle differenze genetiche Osservabile a livello di popolazione, non di singolo individuo! Oltre ad essere poligeniche, le caratteristiche quantitative sono spesso influenzate da fattori ambientali. E quindi utile sapere quanto la variabilità di un carattere quantitativo sia dovuta a differenze genetiche e quanto a differenze ambientali. La quota di variazione fenotipica complessiva derivante da fattori genetici è nota come ereditabilità
36 Ereditabilità di un carattere (H 2 ) Esprime la proporzione della variabilità fenotipica (V P ) imputabile a differenze genetiche (V G ) Il contributo relativo dei fattori genetici e dei fattori ambientali nella determinazione della variabilità di un carattere quantitativo è misurato da un parametro che prende il nome di ereditabilità. Essa consente di prevedere il fenotipo della progenie in base al fenotipo parentale EREDITABILITA : H 2 = V G V P frazione ereditabile della variazione di un carattere quantitativo H 2 = 0-1 Per determinare l ereditabilità di un carattere quantitativo occorre per prima cosa misurarne la variabilità - varianza (misura la fluttuazione dei dati individuali intorno alla media del campione ed equivale allo scarto quadratico medio) - e quindi suddividere questa nelle sue componenti.
37 COMPONENTI DELLA VARIANZA FENOTIPICA (V P ) Alcune differenze fenotipiche sono dovute a fattori genetici e a fattori ambientali. VARIANZA GENETICA = V G (differenze genetiche tra gli individui) VARIANZA AMBIENTALE = V E (influenza delle condizioni ambientali) V P =V G +V E

38 Esempio di interazione genotipo ambiente. Il genotipo influisce sul peso della pianta, ma sono le condizioni climatiche a determinare quale genotipo produca l esemplare piu pesante! Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
39 Una certa quota di variazione genetica è ereditabile poiché passa dai genitori ai figli. Un altra quota di variazione è genetica ma non strettamente ereditabile a causa di fenomeni quali la dominanza e le interazioni epistatiche che non passano direttamente dai genitori ai figli. Per esempio, se un allele è dominante su un altro, il fenotipo di un genitore eterozigote è determinato in parte dalla interazione di dominanza fra i due alleli. Un genitore che si riproduce sessualmente passerà alla sua prole soltanto un allele. La prole, infatti, eredita il genotipo da entrambi i genitori e quindi non erediterà l interazione di dominanza, ma soltanto l effetto di un singolo allele

40 COMPONENTI DELLA VARIANZA GENETICA (V G ) La variazione genetica può quindi essere ulteriormente suddivisa in tre componenti: 1. variazione genetica additiva: alcuni alleli possono contribuire con un valore fisso al valore metrico di un carattere quantitativo. Tali geni si definiscono additivi e contribuiscono alla varianza genetica additiva (V A ). 2. variazione genetica dominante: alcuni alleli sono dominanti su altri e mascherano il contributo degli alleli recessivi in quel locus. Questa fonte di variabilità contribuisce alla varianza genetica da dominanza (V D ). 3. variazione genetica causata dalle interazioni fra geni diversi, dovuta fondamentalmente a fenomeni di epistasi. Tale variazione contribuisce alla varianza genetica da interazione (V I ) V G = V A + V D + V I
41 COMPONENTI DELLA VARIANZA GENETICA (V G ) VARIANZA ADDITIVA = V A VARIANZA dovuta alla DOMINANZA = V D VARIANZA EPISTATICA = V I V G = V A + V D + V I e la varianza fenotipica totale può essere riscritta come: V P = V E + V A + V D + V I
42 EREDITABILITA Il termine generale che descrive la proporzione della varianza genetica alla varianza totale è l ereditabilità. Si possono stimare due tipi specifici di ereditabilità. In senso lato l ereditabilità è il rapporto fra la varianza genetica totale (incluse dominanza ed epistasi) e la varianza fenotipica totale, cioè: H 2 = V G /V P In senso stretto l ereditabilità è il rapporto fra la varianza genetica additiva e la varianza fenotipica totale, cioè: h 2 varia tra 0 (V A =0) e 1(V A =V P ) h 2 = V A /V P Per predire il fenotipo della prole proveniente da un particolare accoppiamento è importante conoscere la porzione di varianza genetica additiva, poiché in questo modo sarà possibile conoscere il contributo relativo dei genitori alla costituzione della F1.
43 EREDITABILITA IN SENSO LATO E la frazione di varianza totale che rimane dopo esclusione della frazione dovuta agli effetti ambientali. Una misura del grado di determinazione genetica di un carattere. H 2 = V G V P = V G V G + V E
44 EREDITABILITA IN SENSO STRETTO E il rapporto fra varianza genetica additiva e varianza totale. Una stima dell'efficienza della selezione. h 2 = V A V P = V A V G + V E Solo le differenze genetiche attribuibili alle azioni geniche additive possono essere fissate con la selezione: Essendo legate all effetto medio dei geni, le azioni additive rimangono inalterate nelle generazioni successive; quando invece le differenze tra i materiali selezionati sono dovute a specifiche interazioni geniche, come dominanza ed epistasi, queste non possono essere fissate con la selezione, poichè per effetto della segregazione e della ricombinazione possono ottenersi combinazioni di geni diverse da quelle della generazione precedente.
45 Calcolo dell ereditabilità in senso lato mediante eliminazione di componenti della varianza V P =V G +V E possiamo annullare V E facendo sviluppare gli individui nello stesso ambiente -> impossibile possiamo annullare V G facendo sviluppare individui geneticamente identici se V E = 0, V P = V G se V G = 0, V P = V E V G (degli individui variabili geneticamente) = V p (variabili) V E (identici) H 2 = V G V p Facciamo un esperimento in cui alleviamo individui geneticamente identici in un determinato ambiente e calcoliamo la V P, che in questo caso corrisponde alla V E. Poi prendiamo individui geneticamente diversi e calcoliamo la V P. La V G ora la posso calcolare perche ho la p e la e
46 Calcolo dell ereditabilità in senso lato mediante eliminazione di componenti della varianza: esempio Ereditabilità della chiazzatura bianca nelle cavie nella popolazione variabile geneticamente V P =573 nella popolazione dopo numerosi incroci (omozigote) V P =340 Poichè in quest ultimo gruppo V G =0, V P = V E. Questo valore di V E puo essere assegnato anche alla popolazione di partenza (geneticamente variabile) V P V E = V G = 233 H 2 = V G V p 233 H 2 = 0, % della variazione nella chiazzatura delle cavie era dovuto a differenze riguardanti il genotipo
47 Problemi nel calcolo dell ereditabilità mediante eliminazione di componenti della varianza Si assume che la varianza ambientale degli individui geneticamente identici sia equivalente a quella degli individui variabili geneticamente, e ciò può non essere vero Approccio applicabile solo quando è possibile ottenere individui geneticamente identici
48 Ereditabilità mediante regressione tra individui parentali e prole Si confrontano fenotipi di individui che presentano gradi di parentela diversi. Se la varianza fenotipica è determinata da differenze genetiche: i gemelli monozigotici devono essere simili per un determinato carattere più dei gemelli dizigotici la progenie deve risultare simile ai genitori in misura maggiore a quanto avviene negli individui non imparentati In una regressione del fenotipo medio della prole rispetto ai genitori, l ereditabilità in senso stretto (h 2 ) è uguale al coefficiente di regressione (b). Pertanto, il coefficiente di regressione fornisce informazioni sul grado dell ereditablità
49 Ereditabilità mediante regressione tra individui parentali e prole Il coefficiente di regressione fornisce informazioni sul grado dell ereditablità in (a) non c e alcuna relazione fra fenotipo parentale e della prole (raggruppati in famiglie) in (b) il fenotipo della prole è equivalente a quelli parentali in (c) sia i geni sia l ambiente influiscono sul fenotipo Ogni punto rappresenta un nucleo famigliare, dove sull asse delle X c è il valore fenotipico medio dei parentali di quella famiglia, e sull asse delle y il valore fenotipico medio della prole di quella famiglia. Se per un dato tratto non c è ereditabilità in senso stretto (h2=0) i punti si disperderanno in maniera casuale e il conseguente coefficiente di regressione sarà = 0 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
50 Limitazioni dell ereditabilita Non dice quanto l espressione di un carattere dipenda da fattori genetici, ma esprime solo la proporzione della varianza fenotipica tra gli individui di una popolazione che è dovuta a differenze genetiche. L ereditabilità pertanto indica il grado a cui i geni stabiliscono la variazione di un tratto; Non si riferisce ad un individuo. E una caratteristica di una popolazione (un individuo non presenta ereditabilità!); Non esiste ereditabilità universale per un dato carattere. L ereditabilità è specifica per una data popolazione in uno specifico ambiente; Non può venire usata per trarre conclusioni riguardo la natura di differenze genetiche tra popolazioni, appunto perché è specifica per un data popolazione in un dato ambiente
51 Riepilogo La varianza fenotipica è uguale alla somma della varianza dovuta a differenze genetiche e ambientali Ereditabilità in senso lato è la proporzione di varianza fenotipica dovuta alle differenze genetiche nella popolazione Ereditabilità in senso stretto è la proporzione di varianza fenotipica dovuta agli effetti additivi degli alleli nella popolazione
52 LA GENETICA EVOLUZIONISTICA
53 Evoluzione Alla fine del 1700 la teoria predominante era quella di Linneo, detta della fissità, che definiva le varie specie come entità create una volta per tutte e incapaci di modificarsi o capaci entro ben determinati limiti. All'inizio del XIX secolo iniziarono a sorgere, negli studiosi di Scienze Naturali i primi dubbi concreti: negli strati rocciosi più antichi infatti mancano totalmente tracce (fossili) degli esseri attualmente viventi e se ne rinvengono altre appartenenti ad organismi attualmente non esistenti. Nel 1809, il naturalista Lamarck presentò per primo una teoria evoluzionista (detta lamarckismo) secondo cui gli organismi viventi si modificherebbero gradualmente nel tempo adattandosi all'ambiente: l'uso o il non uso di determinati organi porterebbe con il tempo ad un loro potenziamento o ad un'atrofia. Tale ipotesi implica quello che oggi viene considerato l'errore di fondo: l'ereditabilità dei caratteri acquisiti (esempio: un culturista non avrà necessariamente figli muscolosi; la muscolosità del culturista è infatti una manifestazione fenotipica, cioè morfologica, derivante dall'interazione dello sportivo con l'ambiente, il continuo sollevare pesi; ma il particolare sviluppo muscolare non è dettato dal suo patrimonio genetico, il genotipo).
54 Lamarck trovò opposizione in Georges L. Chretien Cuvier, il quale aveva elaborato la 'teoria delle catastrofi naturali' secondo la quale la maggior parte degli organismi viventi nel passato sarebbero stati spazzati via da numerosi cataclismi e il mondo infatti sarebbe stato ripopolato dalle specie sopravvissute. Dopo cinquant'anni Darwin formulò una nuova teoria evoluzionista; il noto naturalista, durante il suo viaggio giovanile sul brigantino Beagle, fu colpito dalla variabilità delle forme viventi che aveva avuto modo di osservare nei loro ambienti naturali intorno al mondo. Riflettendo sugli appunti di viaggio e traendo spunto dagli scritti dell'economista Thomas Malthus, Darwin si convinse che la lotta per la vita fosse uno dei motori principali dell'evoluzione intuendo il ruolo selettivo dell'ambiente sulle specie viventi. L'ambiente, infatti, non può essere la causa primaria nel processo di evoluzione (come invece sostenuto nella teoria di Lamarck) in quanto tale ruolo è giocato dalle mutazioni genetiche, in gran parte casuali. L'ambiente entra in azione in un secondo momento, nella determinazione del vantaggio o svantaggio riproduttivo che quelle mutazioni danno alla specie mutata, in poche parole, al loro migliore o peggiore adattamento (fitness in inglese). I principali meccanismi che partecipano in queste situazioni sono: * meccanismi genetici * meccanismi ecologici

55 Specie Un insieme di popolazioni formate da individui interfecondi, la cui prole a sua volta è interfeconda: pool genico chiuso
56 Evoluzione E il cambiamento della variabilità in una popolazione valutato nel tempo Le forze dell Evoluzione: Mutazione Selezione Deriva genetica: Isolamento geografico e migrazioni L evoluzione di una nuova specie da una specie ancestrale avviene sulla base della selezione naturale. Le variazioni casuali favorevoli tendono ad essere conservate, quelle non favorevoli tendono ad essere eliminate dalla pressione ambientale: adattamento all ambiente che cambia (modificazione evolutiva che aumenta la probabilità di sopravivenza di un organismo e il suo successo evolutivo viene selezionata).
57 Filogenesi E il percorso evolutivo che ha portato alla differenziazione delle specie a partire da un origine comune. La sua ricostruzione è fondamentale per la sistematica che si occupa di ricostruire le relazioni di parentela evolutiva, di gruppi tassonomici di organismi a qualunque livello sistematico La filogenetica studia l'origine e l'evoluzione di un insieme di organismi, solitamente di una specie. Un compito essenziale della sistematica è di determinare le relazioni ancestrali fra specie note (vive ed estinte). Rapporti filogenetici: i rapporti di discendenza comune tra specie o gruppi di ordine superiore
58 La selezione naturale Insieme dei fattori che determinano la sopravvivenza di alcuni organismi rispetto ad altri, all interno di una data popolazione. Essa determina l evoluzione della specie, favorendo gli organismi dotati dei caratteri più adatti in un determinato ambiente, ossia quelli che hanno la fitness maggiore In una popolazione compaiono continuamente nuovi caratteri, e nuove combinazioni di geni, sia per effetto dei meccanismi di mutazione spontanea, sia per la variabilità genetica assicurata dal crossing-over e dalla ricombinazione dei cromosomi nei gameti durante il processo di meiosi; questi fenomeni avvengono in modo casuale, e hanno quindi la stessa probabilità di verificarsi in qualsiasi popolazione. A seconda delle caratteristiche ambientali, però, la possibilità di sopravvivere e di riprodursi di alcuni individui, caratterizzati da un particolare patrimonio genetico, sarà maggiore rispetto a quella di altri, che risultano sfavoriti. Il cambiamento delle condizioni esterne determina inevitabilmente anche un cambiamento della fitness degli individui: quelli che nelle condizioni precedenti erano favoriti, possono in seguito risultare sfavoriti.
59 La selezione naturale Ad ogni generazione viene prodotto un numero sempre maggiore di individui in grado di sopravvivere fino alla riproduzione Il grado di variabilità fenotipica è elevato Una certa quota di questa variabilità fenotipica è ereditabile pertanto Gli individui con determinati caratteri (detti adattativi) sopravvivono e si riproducono in modo più efficiente rispetto agli altri. Questi caratteri sono ereditabili, per cui nella generazione successiva aumenterà in numero di individui che evidenzieranno tali caratteri
60 La selezione artificiale Selezione intenzionale e programmata delle caratteristiche biologiche di una specie o popolazione che non risponde ai criteri di autoorganizzazione propri dell ambiente naturale. La selezione artificiale si attua senza competizione per la sopravvivenza tra individui che mutino in modo casuale. In essa, infatti, i mutanti dotati di una qualsiasi proprietà desiderata possono essere favoriti dall intervento umano mirato alla produzione di particolari razze animali o vegetali mediante incroci selettivi, per scopi zootecnici o agronomici. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
61 La risposta alla selezione (R) Entità di variazione di un carattere sottoposto a selezione, nel corso di una generazione La risposta alla selezione si calcola sottraendo il fenotipo medio della popolazione originale da quello medio della prole. Es. mandria produce 80 l latte a settimana, la progenie ne produce 100. Risposta alla selezione= 20 l a settimana
62 La risposta alla selezione (R) Viene determinata prevalentemente da due fattori: ereditabilità in senso stretto grado di selezione degli individui da incrociare: differenza fenotipica di questi individui, che si misura con il differenziale di selezione (S). R = h 2 x S Il differenziale di selezione (S) è definito come la differenza fra il fenotipo medio degli individui parentali scelti, e quello medio della popolazione originale. Es. mandria produce 80 l latte a settimana, i parentali scelti per l incrocio ne producono 120. Differenziale di selezione = 40 l

63 Risposta alla selezione in mais Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
64 Prevedere la risposta alla selezione (R) R = h 2 x S h 2 = V A V P Numero setole addominali in Drosophila Ereditabilità in senso stretto = 0,52 Numero medio setole in popolazione di partenza = 35,3 Numero medio setole in moscerini selezionati per l incrocio = 40,6 Pertanto, differenziale di selezione = 5,3 R = 0,52 x 5,3 = 2,8 R rappresenta l aumento atteso nella prole rispetto alla popolazione originale
65 L ereditabilità realizzata La risposta alla selezione può essere utilizzata per stimare l ereditabilità Quando G. Clayton osservò il numero medio di setole (37.9), scoprì che questo differiva dal numero stimato: 38.1 ( ) L ereditabilità in senso stretto puo pertanto essere calcolata rielaborando l equazione R = h 2 x S Ereditabilità realizzata h 2 = R S S: differenziale di selezione R: risposta alla selezione L ereditabilità realizzata è definita per mezzo di un esperimento di risposta alla selezione. Calcolare S: media parentali scelti media popolazione di partenza Calcolare R: media progenie media popolazione di partenza

66 Limiti della risposta alla selezione Quando un carattere è selezionato per molte generazioni non risponde più alla selezione, si stabilizza. Questo avviene perché si esaurisce la variabilità genetica della popolazione o perché la selezione naturale si oppone a uteriori cambiamenti: se cerchi di rimpicciolire i topi questi ad un certo punto diventano sterili per cui non posso trasmettere alla progenie i loro geni responsabili della ridotta dimensione Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
67 LA GENETICA DI POPOLAZIONE La genetica di popolazione riguarda la costituzione genetica di un gruppo di individui, e il modo in cui essa muta nel tempo

68 La genetica di popolazione Gli esseri viventi presentano un certo grado di variabilità genetica Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
69 Genetica di popolazione Nasce all inizio del 900 come logica conseguenza dello sviluppo della genetica mendeliana Studia geni e alleli non più nella singola famiglia, o nel singolo incrocio, ma in popolazioni di individui interfecondi Ricerca strumenti matematici per descrivere la variabilità genetica entro una popolazione. La struttura genetica di una popolazione considera i tipi e le frequenze dei genotipi e degli alleli di quella popolazione (frequenze geniche e genotipiche) Sviluppa modelli matematici che spieghino come tale variabilità si mantenga di generazione in generazione ovvero come e perché si verificano cambiamenti nelle frequenze geniche La genetica di popolazione riguarda cioè la costituzione genetica di un gruppo di individui e il modo in cui essa muta nel tempo
70 Popolazioni mendeliane e pool genico Si definisce popolazione mendeliana un gruppo di individui interfecondi che risiede all interno di confini geografici definiti. Se tutti i gameti prodotti da una popolazione mendeliana sono considerati come un ipotetico miscuglio di unità genetiche dalle quali nascerà la generazione successiva, ne emerge il concetto di pool genico. Il pool genico (o pool genetico) di una specie o di una popolazione è il set completo di alleli unici che verrebbero trovati esaminando il materiale genetico di ogni membro vivente di quella specie o popolazione, cioè l'insieme di tutti gli alleli di tutti i geni di tutti gli individui di una popolazione
71 Genetica di popolazione Parametri: Frequenza fenotipica: la proporzione di individui in una popolazione con un dato fenotipo Frequenza genotipica: la proporzione di uno specifico genotipo ad un dato locus, considerando che sono possibili molti genotipi diversi Frequenza allelica: la proporzione di uno specifico allele in un dato locus, considerando che la popolazione può avere da uno a più alleli a quel locus
72 Le frequenze genotipiche Frequenza genotipica: proporzione di uno specifico genotipo ad un dato locus, considerando che sono possibili molti genotipi diversi Si calcola sommando gli individui che possiedono lo specifico genotipo e poi dividendo per l ammontare complessivo di individui del campione frequenza (AA) = # individui AA N (# individui campione) frequenza (Aa) = # individui Aa N (# individui campione) frequenza (aa) = # individui aa N (# individui campione) La somma delle frequenze genotipiche è sempre uguale a 1
73 Le frequenze alleliche Il pool genetico di una popolazione può essere descritto anche in forma di frequenze alleliche: la proporzione di uno specifico allele in un dato locus. Il numero di alleli è sempre inferiore al numero genotipi! Se f(aa), f(aa) e f(aa) sono le frequenze di tre genotipi in un locus con due alleli, allora la frequenza p dell' allele A e la frequenza q dell'allele a si ottengono contando gli alleli. Poiché ogni omozigote AA consiste solo di alleli A, e dato che metà degli alleli di ogni eterozigote Aa sono alleli A, la frequenza totale p di alleli A nella popolazione si calcola come: frequenza di A In modo analogo, la frequenza q dell'allele a è data da frequenza di a Ci si aspetta che la somma di p e q sia 1, dato che essi sono le frequenze degli unici due alleli presenti. Infatti otteniamo: e da quest'ultima: q = 1 p e p = 1 q Se ci sono più di due diverse forme alleliche, la frequenza per ogni allele è semplicemente la frequenza del suo omozigote più la metà della somma delle frequenze per tutti gli altri eterozigoti in cui appare quel determinato allele.
74 Calcolo delle frequenze alleliche Somma delle copie di un particolare allele presente in un campione diviso l ammontare complessivo degli alleli del campione Si consideri una popolazione di dieci individui e un dato locus con due possibili alleli, A e a. Si supponga che i genotipi degli individui siano i seguenti: AA, Aa, AA, aa, Aa, AA, AA, Aa, Aa, e AA p = frequenza (A) = 2n AA + n Aa 2N (ogni individuo diploide possiede due alleli in un locus) q = frequenza (a) = 2n aa + n Aa 2N Si noti che p + q = 1, cioè la percentuale dei gameti A e a deve dare il 100%, se tiene conto di tutti i gameti nel pool genico.
75 Esempio di calcolo delle frequenze alleliche Si consideri una popolazione di dieci individui e un dato locus con due possibili alleli, A e a. Si supponga che i genotipi degli individui siano i seguenti: AA, Aa, AA, aa, Aa, AA, AA, Aa, Aa, e AA le frequenze alleliche dell'allele A e dell'allele a sono: ovvero c'è un 70% di probabilità che la popolazione riceva questo allele, e c'è un 30% di probabilità che la popolazione ottenga questo.
76 LEGGE DI HARDY-WEINBERG (1908) Il fine primario della genetica di popolazione consiste nel comprendere i processi che modellano il pool genico di un gruppo di individui. Bisogna pertanto comprendere gli effetti della riproduzione e delle leggi di Mendel sulle frequenze genotipiche e alleliche. Cioè: in che modo la segregazione degli alleli durante la formazione dei gameti e il loro combinarsi nel corso della fecondazione influenzano il pool di geni? La risposta a questa domanda risiede nella legge formulata indipendentemente da Hardy e da Weinberg nel 1908: modello matematico che valuta l effetto della riproduzione sulle frequenze genotipiche e alleliche di una popolazione
77 LEGGE DI HARDY-WEINBERG (1908) Modello matematico che valuta l effetto della riproduzione sulle frequenze genotipiche e alleliche di una popolazione In una popolazione, le frequenze alleliche e genotipiche si mantengono costanti di generazione in generazione Le frequenze genotipiche sono correlate alle frequenze alleliche da una semplice formula: A con frequenza p a con frequenza q Le frequenze dei tre possibili genotipi sono date da: p 2 + 2pq + q 2 = (p + q) 2 = 1 AA Aa aa p + q = 1
78 Equazione di HARDY-WEINBERG Dal momento che ciascun soggetto ha due alleli per un gene, la distribuzione dei genotipi alla generazione successiva può essere riassunta come segue: (p + q) 2 p 2 + 2pq + q 2 = 1 AA Aa aa Ogni soggetto della popolazione deriva infatti dall unione di due gameti, nei quali i due alleli A ed a possono presentarsi con probabilità p e q rispettivamente. Questa legge indica che la sola riproduzione non altera le frequenze alleliche o genotipiche, e che le frequenze alleliche determinano le frequenze genotipiche Quando i genotipi sono nelle proporzioni attese di p 2, 2pq e q 2, si dice che la popolazione è in equilibrio di Hardy-Weinberg
79 Condizioni necessarie per soddisfare la legge di Hardy e Weinberg 1. La popolazione è infinitamente estesa e si incrocia a caso (PANMISSIA) 2. Non c è selezione in atto a favore o contro un determinato fenotipo, tutti gli individui portatori dell uno o dell altro allele ad un dato locus sono ugualmente vitali e fertili 3. La popolazione è chiusa, cioè non vi sono movimenti migratori in atto 4. Non c è mutazione da uno stato allelico ad un altro, oppure il tasso di mutazione A -> a è uguale ad a -> A Questa legge si applica AD UN SINGOLO LOCUS. Quindi non è proprio necessario l accoppiamento casuale e la mancanza di selezione, migrazione e mutazione per tutte le caratteristiche, bensì solo per quel locus
80 Sommario condizioni necessarie per soddisfare la legge di Hardy e Weinberg 1. La popolazione è sufficientemente ampia 2. Gli individui della popolazione si accoppiano in maniera casuale 3. Le frequenze alleliche della popolazione non sono influenzate da selezione naturale, migrazioni, mutazioni
81 Implicazioni della legge di Hardy e Weinberg Se le condizioni elencate sopra fossero sempre rispettate, cioè se la popolazione fosse in equilibrio: L accoppiamento casuale genererebbe le frequenze di equilibrio p 2, 2pq, q 2 le frequenze genotipiche nelle popolazioni sarebbero sempre in equilibrio e IMMUTABILI nel tempo, cioè non ci sarebbe alcuna EVOLUZIONE Le frequenze genotipiche sarebbero stabilite dalle frequenze alleliche Questa legge afferma che la riproduzione, da sola, non potrà determinare l evoluzione
82 Quando una popolazione è in equilibrio di Hardy-Weinberg, le proporzioni dei genotipi sono stabilite dalle frequenze degli alleli Per un locus con 2 alleli, la frequenza dell eterozigote è più alta delle frequenze degli omozigoti quando le frequenze dei due alleli sono comprese fra 0,33 e 0,66. E massima (0,5) quando entrambe le frequenze sono = 0,5 Frequenza allele bassa: omozigoti rari Frequenza allele alta: omozigoti numerosi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
83 Condizioni necessarie per soddisfare la legge di Hardy e Weinberg Le condizioni elencate in precedenza sono teoriche, e non vengono mai rispettate in natura in modo assoluto Di fatto la legge di H.W. è applicabile a molti studi di popolazione ed è molto utile Bisogna comunque verificare con test statistici (es: test χ2) la sua applicabilità al campione in esame
84 Calcolo delle frequenze alleliche 1. E semplice per un locus con due alleli codominanti (p.es. locus gruppo sanguigno MN) Fenotipi: M MN N Genotipi: MM MN NN In un campione di 100 individui si contano 52 MM 36 MN 12 NN Frequenza dell allele M? n copie M presenti / n totale degli alleli ((52x2) + 36) / 200 = 0,7
85 Calcolo delle frequenze alleliche 2. Frequenza dell allele N? n copie N presenti/ n totale degli alleli ((12x2) + 36) / 200 = 0,3 Se la popolazione campionata rispetta equilibrio di H.W. le distribuzioni genotipiche attese sono: p 2 (MM)= 0,7x0,7= pq (MN)= 2x0,7x0,3=0,42 q 2 (NN)= 0,3x0,3=0,09 Il test del χ2 [Σ (oss.-att.) 2 /att.] mi dice che gli scostamenti fra osservati e attesi NON sono statisticamente significativi, quindi H.W. è rispettato: la popolazione è in equilibrio
86 Utilizzo pratico della legge di Hardy-Weinberg La fibrosi cistica è una malattia recessiva, rara, che colpisce in Italia 1/2000 neonati. Qual e la frequenza dei portatori sani? q 2 = 1/2000 (frequenza di aa) = q = = p = 1-q = ƒ Aa = 2pq = cioè 1/23 La probabilità che due Aa si incrocino è 1/23 x 1/23 = 1/529 Se entrambi i genitori sono Aa, ¼ dei figli sarà aa (malato). Quindi, il rischio che due italiani non imparentati e senza casi di malattia nelle rispettive famiglie abbiano un figlio malato è: 1/4 x 1/23 x 1/23 = 1/2116 cioè molto simile all 1/2000 osservato

87

88
89 L analisi dei profili del DNA si basa sulle assunzioni di Hardy-Weinberg Si studiano quelle parti del genoma che non influiscono sul fenotipo e che quindi non sono soggette a selezione naturale/artificiale, quali le sequenze corte ripetute che ricadono nelle regioni intergeniche
90 Ogni individuo può essere classificato come omozigote o eterozigote in base al numero di copie di una particolare ripetizione presenti nello stesso locus dei due cromosomi omologhi Individuo 1: eterozigote Individuo 2: omozigote Individuo 3: eterozigote
91

92 A ciascun numero di copie è assegnata una frequenza in base alla frequenza osservata in una particolare popolazione Se si considerano ripetizioni su cromosomi diversi, è possibile usare la regola del prodotto per calcolare la probabilità che una particolare combinazione di ripetizioni venga trovata in una popolazione. Si applica poi la legge di Hardy- Weinberg: la disposizione dei frammenti indica se un individuo è omozigote (p 2 o q 2 ) o eterozigote (2pq) per ogni ripetizione. Queste frequenze vengono poi moltiplicate.

93
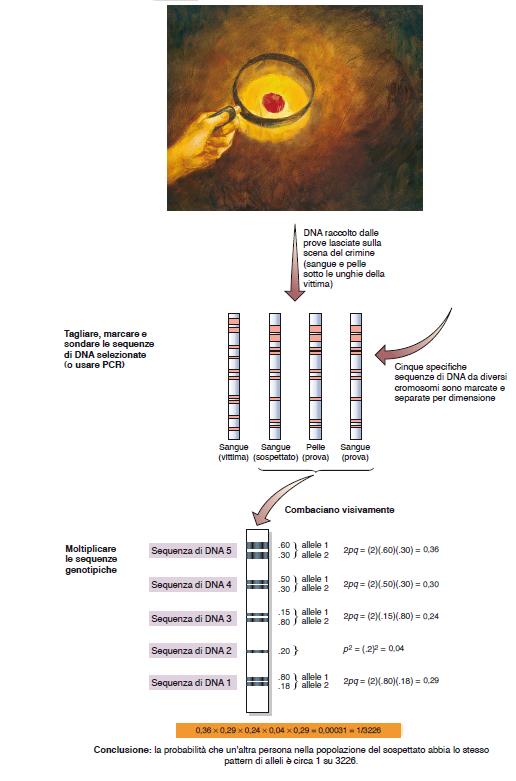

94
 2 = 0,49 = 49% Dd = 2pq = 0,7 x 0,3 =")
95 Dito medio più corto del secondo e del quarto: Carattere autosomico recessivo Se 9 individui su 100 hanno il dito medio più corto (dd), la frequenza è 9/100 o 0,09 Dato che dd corrisponde a q 2, q = 0,3 Dato che p + q = 1, p= 0,7 Quindi la proporzione dei tre genotipi generati quando i gameti si combinano casualmente, nella generazione 1 è: DD = p 2 = (0,7) 2 = 0,49 = 49% Dd = 2pq = 0,7 x 0,3 = 0,42 = 42% dd = q 2 = (0,3) 2 = 0,09 = 9% Quindi 9 individui su 100 avranno il dito medio più corto (dd), mantenendo la frequenza di 9/100 o 0,09 Pertanto la proporzione dei tre genotipi generati quando i gameti si combinano casualmente, nella generazione 2 sarà: DD = p 2 = (0,7) 2 = 0,49 = 49% Dd = 2pq = 0,7 x 0,3 = 0,42 = 42% dd = q 2 = (0,3) 2 = 0,09 = 9%
96 Quando i test statistici mostrano discrepanze significative fra le frequenze genotipiche osservate e le frequenze attese in base alla formula p2, 2pq, q2.. dobbiamo pensare che una o piu delle condizioni di validità della legge siano state violate Per esempio, nel caso del dito medio più corto del secondo e del quarto possiamo ipotizzare che sia errata l ipotesi che le persone scelgano il proprio compagno senza considerare la lunghezza delle dita (accoppiamento quindi non casuale)
97 Fattori che possono disturbare l equilibrio di Hardy-Weinberg Vari fattori possono disturbare l equilibrio di H. W.: 1. Accoppiamento non casuale 2. Mutazione 3. Migrazione 4. Inincrocio 5. Selezione 6. Dimensioni ridotte della popolazione Uno o piu fattori che violino le condizioni di validità della legge di H.W. possono promuovere cambiamenti nelle frequenze alleliche e genotipiche -> EVOLUZIONE
98 Fattori che cambiano frequenze geniche Mutazione Selezione Migrazione Introducono variabilità nel pool genico Dimensioni limitate di una popolazione (<1000 ii) Unioni non casuali (p.es. inincrocio) Riducono la variabilità del pool genico L'inincrocio è l'incrocio fra individui strettamente imparentali o consanguinei

99 Fattori che cambiano frequenze geniche. 1 Accoppiamento non casuale
100 Fattori che cambiano frequenze geniche. 2 Mutazione/Selezione La mutazione - evento raro e casuale - crea nuove varianti alleliche La selezione agisce sugli individui mutanti: se la mutazione è vantaggiosa verrà selezionata positivamente -> aumenta la frequenza della nuova variante allelica
101 Fattori che cambiano frequenze geniche. 3 Migrazione o flusso genico Influsso di geni provenienti da altre popolazioni Previene la differenza genetica fra popolazioni Aumenta la variabilità genetica entro popolazioni Effetti della migrazione sulle frequenze geniche: Δq = variazione frequenza genica i = frazione di immigrati qn = frequenza nei nativi qi = frequenza negli immigrati Δq= i(qn-qi)

102 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
103 Fattori che cambiano frequenze geniche. 4 L inincrocio (inbreeding) Accoppiamento fra individui che condividono una certa quota di alleli identici per discendenza Può essere la conseguenza di matrimoni fra membri di comunità ristrette e geograficamente isolate Può essere la conseguenza di matrimoni combinati fra consanguinei (p.es. primi cugini) per ragioni religiose, culturali, patrimoniali etc. Aumenta la probabilità di omozigosi nella prole per alleli recessivi rari Comparsa di malattie AR rare

104 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

105 Per la maggior parte delle specie a fecondazione incrociata, l inincrocio è dannoso in quanto aumenta la proporzione degli omozigoti favorendo quindi la comparsa di omozigoti recessivi deleteri o addirittura letali Depressione da inincrocio Riduzione della fitness conseguente alla pratica sistematica dell inbreeding Dovuta all omozigosi per alleli recessivi con effetto deleterio Si osservano ad es. riduzione di fertilità, ridotto tasso di sopravvivenza, comparsa di malformazioni, aumentata suscettibilità alle infezioni

106 Eteròsi (o vigore) dell ibrido L'eterosi si riferisce all'incrocio tra individui non imparentati. Il termine è l'opposto di inbreeding che si riferisce all'incrocio tra consanguinei. La popolazione prodotto di eterosi risulta con un genotipo in cui aumenta la frequenza di eterozigosi. Al contrario nell'inbreeding aumenta la omozigosi cioè la presenza di alleli identici nello stesso locus. All'eterosi si associa il fenomeno osservato detto vigore dell'ibrido, in cui l'individuo prodotto dell'accoppiamento risulta con caratteristiche di fenotipo particolarmente vigorose
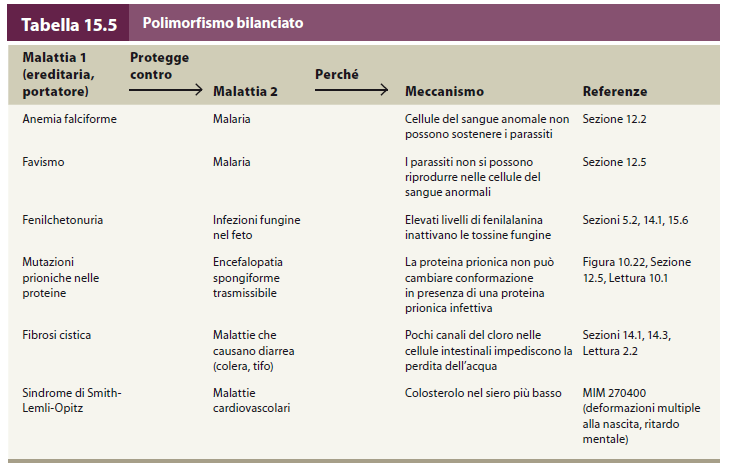
107 Vantaggio dell eterozigote si è verificato in africa occ. in ambiente malarico. gli eterozigoti β a β s hanno fitness maggiore degli omozigoti β a β a, perché risultano resistenti alla malaria (i loro globuli rossi sono inospitali al protozoo plasmodium) e anche degli omozigoti β s β s, gravemente colpiti dalla anemia falciforme. Quindi l allele dannoso viene mantenuto nella popolazione perché porta un vantaggio: polimorfismo bilanciato
108 Polimorfismo bilanciato In una popolazione, mantenimento di più di un allele in conseguenza della superiorità adattativa degli eterozigoti rispetto agli omozigoti. Rappresenta il risultato della pressione selettiva su una mutazione sfavorevole, che ne permette la persistenza in una popolazione. Per esempio, la persistenza del gene per l'anemia falciforme nelle popolazioni africane grazie alla protezione contro la malaria manifestata dagli eterozigoti. La diffusione della malaria rende la situazione eterozigote molto favorevole dal punto di vista evolutivo. In questo modo le frequenze dei fenotipi omozigoti ed eterozigoti si mantengono relativamente stabili.

109
110

111 Fattori che cambiano le frequenze geniche. 5 La selezione (naturale e artificiale)
112 Fattori che cambiano le frequenze geniche. 6 La deriva genetica La deriva genetica (Drift) è la variazione, dovuta al caso, delle frequenze alleliche in una piccola popolazione La deriva genetica è principalmente causata dall effetto del fondatore e dal collo di bottiglia

113 La deriva genetica fa variare le frequenze alleliche all interno delle popolazioni, portando a una riduzione di variabilità genetica attraverso la fissazione e la divergenza tra le popolazioni Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

114 In questo esperimento sono prese in esame le frequenze di due alleli (bw75 e bw) che influiscono sul colore dell occhio in drosofila, in 107 popolazioni replicate, costituite da 8 maschi e 8 femmine, in cui la frequenza iniziale di bw75 era 0,5 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
115 Cause della deriva genetica EFFETTO DEL FONDATORE Si ha quando una nuova popolazione viene stabilita da un numero limitato di individui. Es. Situazioni di isolamento ecologico

116
117

118 Cause della deriva genetica EFFETTO COLLO DI BOTTIGLIA (Bottleneck) Si ha quando una popolazione si trova in condizioni sfavorevoli che riducono drasticamente il numero di individui Determina variazioni casuali nelle frequenze alleliche simili a quelle dovute all effetto del fondatore
Genetica. Mendel e la genetica
 Genetica Le leggi dell ereditarietà di Mendel Ereditarietà e cromosomi Estensioni della genetica mendeliana Applicazioni della genetica Genoma umano Mendel e la genetica Mendel 81822-1884), un monaco di
Genetica Le leggi dell ereditarietà di Mendel Ereditarietà e cromosomi Estensioni della genetica mendeliana Applicazioni della genetica Genoma umano Mendel e la genetica Mendel 81822-1884), un monaco di
GENETICA MENDELIANA NELL UOMO
 GENETICA MENDELIANA NELL UOMO GENETICA FORMALE o GENETICA CLASSICA basata unicamente su risultati visibili di atti riproduttivi. È la parte più antica della genetica, risalendo agli esperimenti di Mendel
GENETICA MENDELIANA NELL UOMO GENETICA FORMALE o GENETICA CLASSICA basata unicamente su risultati visibili di atti riproduttivi. È la parte più antica della genetica, risalendo agli esperimenti di Mendel
La trasmissione dei caratteri ereditari. Le leggi di Mendel (1882-1884)
") La trasmissione dei caratteri ereditari Le leggi di Mendel (1882-1884) Le leggi di Mendel studiano la trasmissione di caratteri qualitativi prodotti da un singolo gene Procedimento sperimentale di Mendel
La trasmissione dei caratteri ereditari Le leggi di Mendel (1882-1884) Le leggi di Mendel studiano la trasmissione di caratteri qualitativi prodotti da un singolo gene Procedimento sperimentale di Mendel
TEORIA CROMOSOMICA : ALLEGATI
 TEORIA CROMOSOMICA : ALLEGATI FIG. 2 a pag. 1 FIG. 5 a pag. 3 FIG. 7 a pag. 5 FIG. 9 a pag. 7 FIG. 3 e 4 a pag. 2 FIG. 6 a pag. 4 FIG. 8 a pag. 6 FIG. 10 e 11 a pag. 8 1 FIGURA 2 Perché sono tutti maschi
TEORIA CROMOSOMICA : ALLEGATI FIG. 2 a pag. 1 FIG. 5 a pag. 3 FIG. 7 a pag. 5 FIG. 9 a pag. 7 FIG. 3 e 4 a pag. 2 FIG. 6 a pag. 4 FIG. 8 a pag. 6 FIG. 10 e 11 a pag. 8 1 FIGURA 2 Perché sono tutti maschi
Prima Legge di Mendel LEGGE DELLA SEGREGAZIONE IN PROPORZIONI UGUALI:
 Prima Legge di Mendel LEGGE DELLA SEGREGAZIONE IN PROPORZIONI UGUALI: Durante la meiosi, i membri di una coppia allelica si separano in modo simmetrico nelle uova e negli spermatozoi. Questa separazione
Prima Legge di Mendel LEGGE DELLA SEGREGAZIONE IN PROPORZIONI UGUALI: Durante la meiosi, i membri di una coppia allelica si separano in modo simmetrico nelle uova e negli spermatozoi. Questa separazione
La Genetica. Le leggi di Mendel
 La Genetica Le leggi di Mendel La Genetica Il monaco Gregor Mendel (1822-1884) fu il primo a studiare in modo rigoroso il fenomeno della trasmissione dei caratteri ereditari. Per questo, pur non avendo
La Genetica Le leggi di Mendel La Genetica Il monaco Gregor Mendel (1822-1884) fu il primo a studiare in modo rigoroso il fenomeno della trasmissione dei caratteri ereditari. Per questo, pur non avendo
CORSO INTEGRATO DI GENETICA
 CORSO INTEGRATO DI GENETICA a.a.2011-2012 11.10.2011 Lezioni N. 7 e 8 Ereditarietà Mendeliana Segregazione alleli, indipendenza geni, associazione, ricombinazione Dott.ssa Elisabetta Trabetti UN GENE =
CORSO INTEGRATO DI GENETICA a.a.2011-2012 11.10.2011 Lezioni N. 7 e 8 Ereditarietà Mendeliana Segregazione alleli, indipendenza geni, associazione, ricombinazione Dott.ssa Elisabetta Trabetti UN GENE =
www.fisiokinesiterapia.biz LE LEGGI DI MENDEL
 www.fisiokinesiterapia.biz LE LEGGI DI MENDEL Gregor Johann Mendel (1822-1884) Comprese i principi che regolano la trasmissione dei caratteri ereditari alla progenie senza conoscere - l esistenza dei geni
www.fisiokinesiterapia.biz LE LEGGI DI MENDEL Gregor Johann Mendel (1822-1884) Comprese i principi che regolano la trasmissione dei caratteri ereditari alla progenie senza conoscere - l esistenza dei geni
= femmina. = maschio. = fenotipo banda bianca. = fenotipo pezzato. =fenotipo colore uniforme
 Test n.8 Dalle Olimpiadi delle Scienze Naturali 2002 PARTE TERZA Le 5 domande di questa parte riguardano il medesimo argomento e sono introdotte da un breve testo e da uno schema. In una razza bovina il
Test n.8 Dalle Olimpiadi delle Scienze Naturali 2002 PARTE TERZA Le 5 domande di questa parte riguardano il medesimo argomento e sono introdotte da un breve testo e da uno schema. In una razza bovina il
GRUPPI SANGUIGNI. Supponendo che la popolazione italiana sia H-W, calcola la probabilità di ogni singolo allele e di ogni genotipo
 GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
Le leggi di Mendel esposte in modo ragionato e critico di Luciano Porta
 Le leggi di Mendel esposte in modo ragionato e critico di Luciano Porta Le tre leggi di Mendel, che descrivono la trasmissione dei caratteri ereditari da una generazione all altra, segnano l inizio della
Le leggi di Mendel esposte in modo ragionato e critico di Luciano Porta Le tre leggi di Mendel, che descrivono la trasmissione dei caratteri ereditari da una generazione all altra, segnano l inizio della
SOMIGLIANZA TRA INDIVIDUI
 SOMIGLIANZA TRA INDIVIDUI Si riferisce alla possibilità di riconoscere delle caratteristiche fenotipiche comuni tra animali diversi, dovute alla loro parentela. PARENTELA Due individui si dicono parenti
SOMIGLIANZA TRA INDIVIDUI Si riferisce alla possibilità di riconoscere delle caratteristiche fenotipiche comuni tra animali diversi, dovute alla loro parentela. PARENTELA Due individui si dicono parenti
Page 1. Evoluzione. Intelligenza Artificiale. Algoritmi Genetici. Evoluzione. Evoluzione: nomenclatura. Corrispondenze natura-calcolo
 Evoluzione In ogni popolazione si verificano delle mutazioni. Intelligenza Artificiale In un ambiente che varia, le mutazioni possono generare individui che meglio si adattano alle nuove condizioni. Questi
Evoluzione In ogni popolazione si verificano delle mutazioni. Intelligenza Artificiale In un ambiente che varia, le mutazioni possono generare individui che meglio si adattano alle nuove condizioni. Questi
ESTENSIONI DELLE LEGGI DI MENDEL
 ESTENSIONI DELLE LEGGI DI MENDEL Mendel rinuncia alle sue ricerche Mendel proseguì le sue ricerche su altre piante per ottenere conferme alle sue leggi, ma trovò tante e tali contraddizioni che (si dice)
ESTENSIONI DELLE LEGGI DI MENDEL Mendel rinuncia alle sue ricerche Mendel proseguì le sue ricerche su altre piante per ottenere conferme alle sue leggi, ma trovò tante e tali contraddizioni che (si dice)
Il concetto di valore medio in generale
 Il concetto di valore medio in generale Nella statistica descrittiva si distinguono solitamente due tipi di medie: - le medie analitiche, che soddisfano ad una condizione di invarianza e si calcolano tenendo
Il concetto di valore medio in generale Nella statistica descrittiva si distinguono solitamente due tipi di medie: - le medie analitiche, che soddisfano ad una condizione di invarianza e si calcolano tenendo
GENETICA... lessico. Genetica: studio dei geni e dell'ereditarietà
 GENETICA... lessico Genetica: studio dei geni e dell'ereditarietà Geni: porzioni di DNA contenenti un'informazione che permette di decodificare una certa proteina. Es: gene che determina il colore dei
GENETICA... lessico Genetica: studio dei geni e dell'ereditarietà Geni: porzioni di DNA contenenti un'informazione che permette di decodificare una certa proteina. Es: gene che determina il colore dei
OMOZIGOTE Dominante. OMOZIGOTE Recessivo ETEROZIGOTE
 GENI E CARATTERI EREDITARI I caratteri ereditari corrispondono a precisi tratti di DNA, i geni, che contengono le informazioni per la sintesi delle proteine. Ciascun gene occupa nel cromosoma una determinata
GENI E CARATTERI EREDITARI I caratteri ereditari corrispondono a precisi tratti di DNA, i geni, che contengono le informazioni per la sintesi delle proteine. Ciascun gene occupa nel cromosoma una determinata
Statistica inferenziale
 Statistica inferenziale Popolazione e campione Molto spesso siamo interessati a trarre delle conclusioni su persone che hanno determinate caratteristiche (pazienti, atleti, bambini, gestanti, ) Osserveremo
Statistica inferenziale Popolazione e campione Molto spesso siamo interessati a trarre delle conclusioni su persone che hanno determinate caratteristiche (pazienti, atleti, bambini, gestanti, ) Osserveremo
Relazioni statistiche: regressione e correlazione
 Relazioni statistiche: regressione e correlazione È detto studio della connessione lo studio si occupa della ricerca di relazioni fra due variabili statistiche o fra una mutabile e una variabile statistica
Relazioni statistiche: regressione e correlazione È detto studio della connessione lo studio si occupa della ricerca di relazioni fra due variabili statistiche o fra una mutabile e una variabile statistica
GENETICA MENDELIANA. Per i suoi studi, Mendel utilizzò piante di pisello odoroso (Pisum sativum) Facilità di coltivazione. Disponibilità di varietà
 Facilità di coltivazione. Disponibilità di varietà") GENETICA: è la scienza che studia i caratteri ereditari degli organismi viventi, i meccanismi attraverso i quali si trasmettono ai discendenti e le modalità con cui si manifestano. La genetica moderna
GENETICA: è la scienza che studia i caratteri ereditari degli organismi viventi, i meccanismi attraverso i quali si trasmettono ai discendenti e le modalità con cui si manifestano. La genetica moderna
LA CRESCITA DELLE POPOLAZIONI ANIMALI
 LA CRESCITA DELLE POPOLAZIONI ANIMALI Riccardo Scipioni Generalmente, con il termine crescita di una popolazione si intende l aumento, nel tempo, del numero di individui appartenenti ad una stessa popolazione.
LA CRESCITA DELLE POPOLAZIONI ANIMALI Riccardo Scipioni Generalmente, con il termine crescita di una popolazione si intende l aumento, nel tempo, del numero di individui appartenenti ad una stessa popolazione.
Probabilità condizionata: p(a/b) che avvenga A, una volta accaduto B. Evento prodotto: Evento in cui si verifica sia A che B ; p(a&b) = p(a) x p(b/a)
 che avvenga A, una volta accaduto B. Evento prodotto: Evento in cui si verifica sia A che B ; p(a&b) = p(a) x p(b/a)") Probabilità condizionata: p(a/b) che avvenga A, una volta accaduto B Eventi indipendenti: un evento non influenza l altro Eventi disgiunti: il verificarsi di un evento esclude l altro Evento prodotto:
Probabilità condizionata: p(a/b) che avvenga A, una volta accaduto B Eventi indipendenti: un evento non influenza l altro Eventi disgiunti: il verificarsi di un evento esclude l altro Evento prodotto:
EREDITA MENDELIANA IL CARATTERE E TRASMESSO CON GLI AUTOSOMI O E ASSOCIATO AI CROMOSOMI SESSUALI?
 EREDITA MENDELIANA IL CARATTERE E TRASMESSO CON GLI AUTOSOMI O E ASSOCIATO AI CROMOSOMI SESSUALI? CARATTERE AUTOSOMICO -codificato da geni su cromosomi non sessuali -non ci sono differenze di trasmissione
EREDITA MENDELIANA IL CARATTERE E TRASMESSO CON GLI AUTOSOMI O E ASSOCIATO AI CROMOSOMI SESSUALI? CARATTERE AUTOSOMICO -codificato da geni su cromosomi non sessuali -non ci sono differenze di trasmissione
LA GENETICA. Dott.ssa Valentina Terio
 LA GENETICA Dott.ssa Valentina Terio LLA GENETCA SCIENZA NATA CIRCA 150 ANNI FA GRAZIE AD UN STUDIOSO AUSTRIACO DI NOME MENDEL Pisello da giardino per la facilità di crescita e la possibilità di una impollinazione
LA GENETICA Dott.ssa Valentina Terio LLA GENETCA SCIENZA NATA CIRCA 150 ANNI FA GRAZIE AD UN STUDIOSO AUSTRIACO DI NOME MENDEL Pisello da giardino per la facilità di crescita e la possibilità di una impollinazione
1. Distribuzioni campionarie
 Università degli Studi di Basilicata Facoltà di Economia Corso di Laurea in Economia Aziendale - a.a. 2012/2013 lezioni di statistica del 3 e 6 giugno 2013 - di Massimo Cristallo - 1. Distribuzioni campionarie
Università degli Studi di Basilicata Facoltà di Economia Corso di Laurea in Economia Aziendale - a.a. 2012/2013 lezioni di statistica del 3 e 6 giugno 2013 - di Massimo Cristallo - 1. Distribuzioni campionarie
RELAZIONE TRA VARIABILI QUANTITATIVE. Lezione 7 a. Accade spesso nella ricerca in campo biomedico, così come in altri campi della
 RELAZIONE TRA VARIABILI QUANTITATIVE Lezione 7 a Accade spesso nella ricerca in campo biomedico, così come in altri campi della scienza, di voler studiare come il variare di una o più variabili (variabili
RELAZIONE TRA VARIABILI QUANTITATIVE Lezione 7 a Accade spesso nella ricerca in campo biomedico, così come in altri campi della scienza, di voler studiare come il variare di una o più variabili (variabili
Calcolo delle probabilità
 Calcolo delle probabilità Laboratorio di Bioinformatica Corso A aa 2005-2006 Statistica Dai risultati di un esperimento si determinano alcune caratteristiche della popolazione Calcolo delle probabilità
Calcolo delle probabilità Laboratorio di Bioinformatica Corso A aa 2005-2006 Statistica Dai risultati di un esperimento si determinano alcune caratteristiche della popolazione Calcolo delle probabilità
I.7.1 Malattie genetiche legate al sesso
 verificare tutti i possibili risultati della fecondazione tra cellula uovo e spermatozoi e constatare come le probabilità che nasca una femmina o un maschio sono entrambe pari al 50%. Figura 7 - Ad ogni
verificare tutti i possibili risultati della fecondazione tra cellula uovo e spermatozoi e constatare come le probabilità che nasca una femmina o un maschio sono entrambe pari al 50%. Figura 7 - Ad ogni
CORSO INTEGRATO DI GENETICA ESERCIZI
 CORSO INTEGRATO DI GENETICA A.A. 2007-2008 15 novembre 2007 dott.ssa E. Trabetti Lezioni 37-38 ESERCIZI (1) Una coppia sarda di rivolge a voi per una consulenza genetica poiché, nel corso di uno screening
CORSO INTEGRATO DI GENETICA A.A. 2007-2008 15 novembre 2007 dott.ssa E. Trabetti Lezioni 37-38 ESERCIZI (1) Una coppia sarda di rivolge a voi per una consulenza genetica poiché, nel corso di uno screening
Corso di. Dott.ssa Donatella Cocca
 Corso di Statistica medica e applicata Dott.ssa Donatella Cocca 1 a Lezione Cos'è la statistica? Come in tutta la ricerca scientifica sperimentale, anche nelle scienze mediche e biologiche è indispensabile
Corso di Statistica medica e applicata Dott.ssa Donatella Cocca 1 a Lezione Cos'è la statistica? Come in tutta la ricerca scientifica sperimentale, anche nelle scienze mediche e biologiche è indispensabile
4 modulo didattico - Modalità di trasmissione delle malattie
 4 modulo didattico - Modalità di trasmissione delle malattie monogeniche. L analisi dell albero genealogico: uno strumento indispensabile della genetica medica I SIMBOLI DELL ALBERO GENEALOGICO L ANEMIA
4 modulo didattico - Modalità di trasmissione delle malattie monogeniche. L analisi dell albero genealogico: uno strumento indispensabile della genetica medica I SIMBOLI DELL ALBERO GENEALOGICO L ANEMIA
Mendeliana Autosomica Dominante (AD) Autosomica Recessiva (AR) X-linked Recessiva (X-linked R) X-linked Dominante (X-linked D) Y-linked
 Autosomica Recessiva (AR) X-linked Recessiva (X-linked R) X-linked Dominante (X-linked D) Y-linked") Trasmissione ereditaria di un singolo gene (eredità monofattoriale) Mendeliana Autosomica Dominante (AD) Autosomica Recessiva (AR) X-linked Recessiva (X-linked R) X-linked Dominante (X-linked D) Y-linked
Trasmissione ereditaria di un singolo gene (eredità monofattoriale) Mendeliana Autosomica Dominante (AD) Autosomica Recessiva (AR) X-linked Recessiva (X-linked R) X-linked Dominante (X-linked D) Y-linked
Test statistici di verifica di ipotesi
 Test e verifica di ipotesi Test e verifica di ipotesi Il test delle ipotesi consente di verificare se, e quanto, una determinata ipotesi (di carattere biologico, medico, economico,...) è supportata dall
Test e verifica di ipotesi Test e verifica di ipotesi Il test delle ipotesi consente di verificare se, e quanto, una determinata ipotesi (di carattere biologico, medico, economico,...) è supportata dall
STATISTICA IX lezione
 Anno Accademico 013-014 STATISTICA IX lezione 1 Il problema della verifica di un ipotesi statistica In termini generali, si studia la distribuzione T(X) di un opportuna grandezza X legata ai parametri
Anno Accademico 013-014 STATISTICA IX lezione 1 Il problema della verifica di un ipotesi statistica In termini generali, si studia la distribuzione T(X) di un opportuna grandezza X legata ai parametri
Metodi statistici per le ricerche di mercato
 Metodi statistici per le ricerche di mercato Prof.ssa Isabella Mingo A.A. 2014-2015 Facoltà di Scienze Politiche, Sociologia, Comunicazione Corso di laurea Magistrale in «Organizzazione e marketing per
Metodi statistici per le ricerche di mercato Prof.ssa Isabella Mingo A.A. 2014-2015 Facoltà di Scienze Politiche, Sociologia, Comunicazione Corso di laurea Magistrale in «Organizzazione e marketing per
1) Si consideri un esperimento che consiste nel lancio di 5 dadi. Lo spazio campionario:
 Si consideri un esperimento che consiste nel lancio di 5 dadi. Lo spazio campionario:") Esempi di domande risposta multipla (Modulo II) 1) Si consideri un esperimento che consiste nel lancio di 5 dadi. Lo spazio campionario: 1) ha un numero di elementi pari a 5; 2) ha un numero di elementi
Esempi di domande risposta multipla (Modulo II) 1) Si consideri un esperimento che consiste nel lancio di 5 dadi. Lo spazio campionario: 1) ha un numero di elementi pari a 5; 2) ha un numero di elementi
CAPITOLO 9 EREDITA ED EREDITABILITA DEI CARATTERI QUANTITATIVI LIGUORI EDITORE
 CAPITOLO 9 EREDITA ED EREDITABILITA DEI CARATTERI QUANTITATIVI LIGUORI EDITORE 1 Gli studi genetici presentati hanno riguardato soprattutto studi di sostituzione alleliche che causano differenze qualitative
CAPITOLO 9 EREDITA ED EREDITABILITA DEI CARATTERI QUANTITATIVI LIGUORI EDITORE 1 Gli studi genetici presentati hanno riguardato soprattutto studi di sostituzione alleliche che causano differenze qualitative
Slide Cerbara parte1 5. Le distribuzioni teoriche
 Slide Cerbara parte1 5 Le distribuzioni teoriche I fenomeni biologici, demografici, sociali ed economici, che sono il principale oggetto della statistica, non sono retti da leggi matematiche. Però dalle
Slide Cerbara parte1 5 Le distribuzioni teoriche I fenomeni biologici, demografici, sociali ed economici, che sono il principale oggetto della statistica, non sono retti da leggi matematiche. Però dalle
La trasmissione delle malattie genetiche. Anna Onofri
 La trasmissione delle malattie genetiche Gli alberi genealogici Anna Onofri I simboli maggiormente utilizzati Le malattie genetiche Molte malattie genetiche sono legate ad un singolo gene e possono verificarsi
La trasmissione delle malattie genetiche Gli alberi genealogici Anna Onofri I simboli maggiormente utilizzati Le malattie genetiche Molte malattie genetiche sono legate ad un singolo gene e possono verificarsi
Trasmissione del materiale ereditario
 Trasmissione del materiale ereditario Confronto tra mitosi e meiosi: La mitosi consiste in una duplicazione dei cromosomi seguita da una regolare separazione Ciascun cromosoma si comporta indipendentemente
Trasmissione del materiale ereditario Confronto tra mitosi e meiosi: La mitosi consiste in una duplicazione dei cromosomi seguita da una regolare separazione Ciascun cromosoma si comporta indipendentemente
La distribuzione Normale. La distribuzione Normale
 La Distribuzione Normale o Gaussiana è la distribuzione più importante ed utilizzata in tutta la statistica La curva delle frequenze della distribuzione Normale ha una forma caratteristica, simile ad una
La Distribuzione Normale o Gaussiana è la distribuzione più importante ed utilizzata in tutta la statistica La curva delle frequenze della distribuzione Normale ha una forma caratteristica, simile ad una
VERIFICA DELLE IPOTESI
 VERIFICA DELLE IPOTESI Nella verifica delle ipotesi è necessario fissare alcune fasi prima di iniziare ad analizzare i dati. a) Si deve stabilire quale deve essere l'ipotesi nulla (H0) e quale l'ipotesi
VERIFICA DELLE IPOTESI Nella verifica delle ipotesi è necessario fissare alcune fasi prima di iniziare ad analizzare i dati. a) Si deve stabilire quale deve essere l'ipotesi nulla (H0) e quale l'ipotesi
CAPITOLO III CONFRONTI TRA DISTRIBUZIONI
 CAPITOLO III CONFRONTI TRA DISTRIBUZIONI 3.1 CONFRONTI TRA DISTRIBUZIONI OSSERVATE E DISTRIBUZIONI TEORICHE OD ATTESE. Nella teoria statistica e nella pratica sperimentale, è frequente la necessità di
CAPITOLO III CONFRONTI TRA DISTRIBUZIONI 3.1 CONFRONTI TRA DISTRIBUZIONI OSSERVATE E DISTRIBUZIONI TEORICHE OD ATTESE. Nella teoria statistica e nella pratica sperimentale, è frequente la necessità di
Un po di statistica. Christian Ferrari. Laboratorio di Matematica
 Un po di statistica Christian Ferrari Laboratorio di Matematica 1 Introduzione La statistica è una parte della matematica applicata che si occupa della raccolta, dell analisi e dell interpretazione di
Un po di statistica Christian Ferrari Laboratorio di Matematica 1 Introduzione La statistica è una parte della matematica applicata che si occupa della raccolta, dell analisi e dell interpretazione di
Metodologia per l analisi dei dati sperimentali L analisi di studi con variabili di risposta multiple: Regressione multipla
 Il metodo della regressione può essere esteso dal caso in cui si considera la variabilità della risposta della y in relazione ad una sola variabile indipendente X ad una situazione più generale in cui
Il metodo della regressione può essere esteso dal caso in cui si considera la variabilità della risposta della y in relazione ad una sola variabile indipendente X ad una situazione più generale in cui
LE FUNZIONI A DUE VARIABILI
 Capitolo I LE FUNZIONI A DUE VARIABILI In questo primo capitolo introduciamo alcune definizioni di base delle funzioni reali a due variabili reali. Nel seguito R denoterà l insieme dei numeri reali mentre
Capitolo I LE FUNZIONI A DUE VARIABILI In questo primo capitolo introduciamo alcune definizioni di base delle funzioni reali a due variabili reali. Nel seguito R denoterà l insieme dei numeri reali mentre
TEORIA DI LAMARCK O DELLA EREDITARIETA DEI CARATTERI ACQUISITI
 L EVOLUZIONE IN PASSATO SI PENSAVA CHE LE SPECIE ANIMALI E VEGETALI CHE OGGI POPOLANO LA TERRA AVESSERO AVUTO SEMPRE LA FORMA ATTUALE. SI CREDEVA CIOE NELLA TEORIA DELCREAZIONISMO (NELLA CREAZIONE ORIGINALE)
L EVOLUZIONE IN PASSATO SI PENSAVA CHE LE SPECIE ANIMALI E VEGETALI CHE OGGI POPOLANO LA TERRA AVESSERO AVUTO SEMPRE LA FORMA ATTUALE. SI CREDEVA CIOE NELLA TEORIA DELCREAZIONISMO (NELLA CREAZIONE ORIGINALE)
MUTAZIONI ED EVOLUZIONE
 MUTAZIONI ED EVOLUZIONE Durante la duplicazione del DNA possono verificarsi errori di copiatura se ad esempio al posto di una base azotata ne viene inserita un altra. In questo caso può succedere che cambi
MUTAZIONI ED EVOLUZIONE Durante la duplicazione del DNA possono verificarsi errori di copiatura se ad esempio al posto di una base azotata ne viene inserita un altra. In questo caso può succedere che cambi
OSSERVAZIONI TEORICHE Lezione n. 4
 OSSERVAZIONI TEORICHE Lezione n. 4 Finalità: Sistematizzare concetti e definizioni. Verificare l apprendimento. Metodo: Lettura delle OSSERVAZIONI e risoluzione della scheda di verifica delle conoscenze
OSSERVAZIONI TEORICHE Lezione n. 4 Finalità: Sistematizzare concetti e definizioni. Verificare l apprendimento. Metodo: Lettura delle OSSERVAZIONI e risoluzione della scheda di verifica delle conoscenze
IL RUOLO DEL CASO NELL'EVOLUZIONE
 IL RUOLO DEL CASO NELL'EVOLUZIONE Docenti: Francesca Del Santo Maria Elena Rossi Discipline: Scienze Sociali Matematica Scuola: Liceo G. Mazzini La Spezia Classe: II A Liceo delle Scienze Sociali Finalità
IL RUOLO DEL CASO NELL'EVOLUZIONE Docenti: Francesca Del Santo Maria Elena Rossi Discipline: Scienze Sociali Matematica Scuola: Liceo G. Mazzini La Spezia Classe: II A Liceo delle Scienze Sociali Finalità
Fondamenti e didattica di Matematica Finanziaria
 Fondamenti e didattica di Matematica Finanziaria Silvana Stefani Piazza dell Ateneo Nuovo 1-20126 MILANO U6-368 silvana.stefani@unimib.it 1 Unità 9 Contenuti della lezione Operazioni finanziarie, criterio
Fondamenti e didattica di Matematica Finanziaria Silvana Stefani Piazza dell Ateneo Nuovo 1-20126 MILANO U6-368 silvana.stefani@unimib.it 1 Unità 9 Contenuti della lezione Operazioni finanziarie, criterio
ELEMENTI DI DEMOGRAFIA
 ELEMENTI DI DEMOGRAFIA 2. Caratteristiche strutturali della popolazione Posa Donato k posa@economia.unisalento.it Maggio Sabrina k s.maggio@economia.unisalento.it UNIVERSITÀ DEL SALENTO DIP.TO DI SCIENZE
ELEMENTI DI DEMOGRAFIA 2. Caratteristiche strutturali della popolazione Posa Donato k posa@economia.unisalento.it Maggio Sabrina k s.maggio@economia.unisalento.it UNIVERSITÀ DEL SALENTO DIP.TO DI SCIENZE
Statistica. Lezione 6
 Università degli Studi del Piemonte Orientale Corso di Laurea in Infermieristica Corso integrato in Scienze della Prevenzione e dei Servizi sanitari Statistica Lezione 6 a.a 011-01 Dott.ssa Daniela Ferrante
Università degli Studi del Piemonte Orientale Corso di Laurea in Infermieristica Corso integrato in Scienze della Prevenzione e dei Servizi sanitari Statistica Lezione 6 a.a 011-01 Dott.ssa Daniela Ferrante
Corrispondenze e funzioni
 Corrispondenze e funzioni L attività fondamentale della mente umana consiste nello stabilire corrispondenze e relazioni tra oggetti; è anche per questo motivo che il concetto di corrispondenza è uno dei
Corrispondenze e funzioni L attività fondamentale della mente umana consiste nello stabilire corrispondenze e relazioni tra oggetti; è anche per questo motivo che il concetto di corrispondenza è uno dei
Laboratorio di Pedagogia Sperimentale. Indice
 INSEGNAMENTO DI LABORATORIO DI PEDAGOGIA SPERIMENTALE LEZIONE III INTRODUZIONE ALLA RICERCA SPERIMENTALE (PARTE III) PROF. VINCENZO BONAZZA Indice 1 L ipotesi -----------------------------------------------------------
INSEGNAMENTO DI LABORATORIO DI PEDAGOGIA SPERIMENTALE LEZIONE III INTRODUZIONE ALLA RICERCA SPERIMENTALE (PARTE III) PROF. VINCENZO BONAZZA Indice 1 L ipotesi -----------------------------------------------------------
Capitolo 3 Riproduzione e trasmissione dei cromosomi
 Capitolo 3 Riproduzione e trasmissione dei cromosomi 3.1 Nelle cellule somatiche del topolino domestico vi sono 40 cromosomi. a. Quanti cromosomi riceve dal padre? b. Quanti autosomi sono presenti nel
Capitolo 3 Riproduzione e trasmissione dei cromosomi 3.1 Nelle cellule somatiche del topolino domestico vi sono 40 cromosomi. a. Quanti cromosomi riceve dal padre? b. Quanti autosomi sono presenti nel
Corso di Laurea in Scienze e Tecnologie Biomolecolari. NOME COGNOME N. Matr.
 Corso di Laurea in Scienze e Tecnologie Biomolecolari Matematica e Statistica II Prova di esame del 18/7/2013 NOME COGNOME N. Matr. Rispondere ai punti degli esercizi nel modo più completo possibile, cercando
Corso di Laurea in Scienze e Tecnologie Biomolecolari Matematica e Statistica II Prova di esame del 18/7/2013 NOME COGNOME N. Matr. Rispondere ai punti degli esercizi nel modo più completo possibile, cercando
LEZIONE 3. Ing. Andrea Ghedi AA 2009/2010. Ing. Andrea Ghedi AA 2009/2010
 LEZIONE 3 "Educare significa aiutare l'animo dell'uomo ad entrare nella totalità della realtà. Non si può però educare se non rivolgendosi alla libertà, la quale definisce il singolo, l'io. Quando uno
LEZIONE 3 "Educare significa aiutare l'animo dell'uomo ad entrare nella totalità della realtà. Non si può però educare se non rivolgendosi alla libertà, la quale definisce il singolo, l'io. Quando uno
generazione filiale ibridi
 La Genetica Mendel, una volta che si fu assicurato di aver piante pure per un determinato carattere iniziò la fecondazione incrociata: ad esempio, prelevò del polline da una varietà dal fiore bianco e
La Genetica Mendel, una volta che si fu assicurato di aver piante pure per un determinato carattere iniziò la fecondazione incrociata: ad esempio, prelevò del polline da una varietà dal fiore bianco e
Rapporto dal Questionari Insegnanti
 Rapporto dal Questionari Insegnanti SCUOLA CHIC81400N N. Docenti che hanno compilato il questionario: 60 Anno Scolastico 2014/15 Le Aree Indagate Il Questionario Insegnanti ha l obiettivo di rilevare la
Rapporto dal Questionari Insegnanti SCUOLA CHIC81400N N. Docenti che hanno compilato il questionario: 60 Anno Scolastico 2014/15 Le Aree Indagate Il Questionario Insegnanti ha l obiettivo di rilevare la
Parte I. Prima Parte
 Parte I Prima Parte Capitolo 1 Introduzione generale 1.1 Il problema dell assegnazione Corsi-Borsisti Il problema dell assegnazione delle borse dei corsi ai vari studenti può essere riassunto nei punti
Parte I Prima Parte Capitolo 1 Introduzione generale 1.1 Il problema dell assegnazione Corsi-Borsisti Il problema dell assegnazione delle borse dei corsi ai vari studenti può essere riassunto nei punti
Alberto Viale I CROMOSOMI
 Alberto Viale I CROMOSOMI DA MENDEL ALLA GENETICA AL DNA ALLE MUTAZIONI I cromosomi sono dei particolari bastoncelli colorati situati nel nucleo delle cellule. Sono presenti nelle cellule di ogni organismo
Alberto Viale I CROMOSOMI DA MENDEL ALLA GENETICA AL DNA ALLE MUTAZIONI I cromosomi sono dei particolari bastoncelli colorati situati nel nucleo delle cellule. Sono presenti nelle cellule di ogni organismo
Siamo così arrivati all aritmetica modulare, ma anche a individuare alcuni aspetti di come funziona l aritmetica del calcolatore come vedremo.
 DALLE PESATE ALL ARITMETICA FINITA IN BASE 2 Si è trovato, partendo da un problema concreto, che con la base 2, utilizzando alcune potenze della base, operando con solo addizioni, posso ottenere tutti
DALLE PESATE ALL ARITMETICA FINITA IN BASE 2 Si è trovato, partendo da un problema concreto, che con la base 2, utilizzando alcune potenze della base, operando con solo addizioni, posso ottenere tutti
PROBABILITA MISURARE L INCERTEZZA Lanciamo due dadi, facciamo la somma dei punteggi ottenuti. Su quale numero mi conviene scommettere?
 Lanciamo due dadi, facciamo la somma dei punteggi ottenuti. Su quale numero mi conviene scommettere? Abbiamo visto nella lezione precedente che lo spazio degli eventi più idoneo a rappresentare l esperimento
Lanciamo due dadi, facciamo la somma dei punteggi ottenuti. Su quale numero mi conviene scommettere? Abbiamo visto nella lezione precedente che lo spazio degli eventi più idoneo a rappresentare l esperimento
L intelligenza numerica
 L intelligenza numerica Consiste nel pensare il mondo in termini di quantità. Ha una forte base biologica, sia gli animali che i bambini molto piccoli sanno distinguere poco e molto. È potentissima e può
L intelligenza numerica Consiste nel pensare il mondo in termini di quantità. Ha una forte base biologica, sia gli animali che i bambini molto piccoli sanno distinguere poco e molto. È potentissima e può
Capitolo 2 - Teoria della manutenzione: classificazione ABC e analisi di Pareto
 Capitolo 2 - Teoria della manutenzione: classificazione ABC e analisi di Pareto Il presente capitolo continua nell esposizione di alcune basi teoriche della manutenzione. In particolare si tratteranno
Capitolo 2 - Teoria della manutenzione: classificazione ABC e analisi di Pareto Il presente capitolo continua nell esposizione di alcune basi teoriche della manutenzione. In particolare si tratteranno
Prova di autovalutazione Prof. Roberta Siciliano
 Prova di autovalutazione Prof. Roberta Siciliano Esercizio 1 Nella seguente tabella è riportata la distribuzione di frequenza dei prezzi per camera di alcuni agriturismi, situati nella regione Basilicata.
Prova di autovalutazione Prof. Roberta Siciliano Esercizio 1 Nella seguente tabella è riportata la distribuzione di frequenza dei prezzi per camera di alcuni agriturismi, situati nella regione Basilicata.
Matematica generale CTF
 Successioni numeriche 19 agosto 2015 Definizione di successione Monotonìa e limitatezza Forme indeterminate Successioni infinitesime Comportamento asintotico Criterio del rapporto per le successioni Definizione
Successioni numeriche 19 agosto 2015 Definizione di successione Monotonìa e limitatezza Forme indeterminate Successioni infinitesime Comportamento asintotico Criterio del rapporto per le successioni Definizione
Verifica di ipotesi e intervalli di confidenza nella regressione multipla
 Verifica di ipotesi e intervalli di confidenza nella regressione multipla Eduardo Rossi 2 2 Università di Pavia (Italy) Maggio 2014 Rossi MRLM Econometria - 2014 1 / 23 Sommario Variabili di controllo
Verifica di ipotesi e intervalli di confidenza nella regressione multipla Eduardo Rossi 2 2 Università di Pavia (Italy) Maggio 2014 Rossi MRLM Econometria - 2014 1 / 23 Sommario Variabili di controllo
( x) ( x) 0. Equazioni irrazionali
 ( x) 0. Equazioni irrazionali") Equazioni irrazionali Definizione: si definisce equazione irrazionale un equazione in cui compaiono uno o più radicali contenenti l incognita. Esempio 7 Ricordiamo quanto visto sulle condizioni di esistenza
Equazioni irrazionali Definizione: si definisce equazione irrazionale un equazione in cui compaiono uno o più radicali contenenti l incognita. Esempio 7 Ricordiamo quanto visto sulle condizioni di esistenza
come nasce una ricerca
 PSICOLOGIA SOCIALE lez. 2 RICERCA SCIENTIFICA O SENSO COMUNE? Paola Magnano paola.magnano@unikore.it ricevimento: martedì ore 10-11 c/o Studio 16, piano -1 PSICOLOGIA SOCIALE COME SCIENZA EMPIRICA le sue
PSICOLOGIA SOCIALE lez. 2 RICERCA SCIENTIFICA O SENSO COMUNE? Paola Magnano paola.magnano@unikore.it ricevimento: martedì ore 10-11 c/o Studio 16, piano -1 PSICOLOGIA SOCIALE COME SCIENZA EMPIRICA le sue
SISTEMI DI NUMERAZIONE E CODICI
 SISTEMI DI NUMERAZIONE E CODICI Il Sistema di Numerazione Decimale Il sistema decimale o sistema di numerazione a base dieci usa dieci cifre, dette cifre decimali, da O a 9. Il sistema decimale è un sistema
SISTEMI DI NUMERAZIONE E CODICI Il Sistema di Numerazione Decimale Il sistema decimale o sistema di numerazione a base dieci usa dieci cifre, dette cifre decimali, da O a 9. Il sistema decimale è un sistema
Esercitazione 1 del corso di Statistica 2 Prof. Domenico Vistocco
 Esercitazione 1 del corso di Statistica 2 Prof. Domenico Vistocco Alfonso Iodice D Enza April 26, 2007 1...prima di cominciare Contare, operazione solitamente semplice, può diventare complicata se lo scopo
Esercitazione 1 del corso di Statistica 2 Prof. Domenico Vistocco Alfonso Iodice D Enza April 26, 2007 1...prima di cominciare Contare, operazione solitamente semplice, può diventare complicata se lo scopo
LA CORRELAZIONE LINEARE
 LA CORRELAZIONE LINEARE La correlazione indica la tendenza che hanno due variabili (X e Y) a variare insieme, ovvero, a covariare. Ad esempio, si può supporre che vi sia una relazione tra l insoddisfazione
LA CORRELAZIONE LINEARE La correlazione indica la tendenza che hanno due variabili (X e Y) a variare insieme, ovvero, a covariare. Ad esempio, si può supporre che vi sia una relazione tra l insoddisfazione
IL RISCHIO D IMPRESA ED IL RISCHIO FINANZIARIO. LA RELAZIONE RISCHIO-RENDIMENTO ED IL COSTO DEL CAPITALE.
 IL RISCHIO D IMPRESA ED IL RISCHIO FINANZIARIO. LA RELAZIONE RISCHIO-RENDIMENTO ED IL COSTO DEL CAPITALE. Lezione 5 Castellanza, 17 Ottobre 2007 2 Summary Il costo del capitale La relazione rischio/rendimento
IL RISCHIO D IMPRESA ED IL RISCHIO FINANZIARIO. LA RELAZIONE RISCHIO-RENDIMENTO ED IL COSTO DEL CAPITALE. Lezione 5 Castellanza, 17 Ottobre 2007 2 Summary Il costo del capitale La relazione rischio/rendimento
Statistica e biometria. D. Bertacchi. Variabili aleatorie. V.a. discrete e continue. La densità di una v.a. discreta. Esempi.
 Iniziamo con definizione (capiremo fra poco la sua utilità): DEFINIZIONE DI VARIABILE ALEATORIA Una variabile aleatoria (in breve v.a.) X è funzione che ha come dominio Ω e come codominio R. In formule:
Iniziamo con definizione (capiremo fra poco la sua utilità): DEFINIZIONE DI VARIABILE ALEATORIA Una variabile aleatoria (in breve v.a.) X è funzione che ha come dominio Ω e come codominio R. In formule:
Esercizio 1 Dato il gioco ({1, 2, 3}, v) con v funzione caratteristica tale che:
 con v funzione caratteristica tale che:") Teoria dei Giochi, Trento, 2004/05 c Fioravante Patrone 1 Teoria dei Giochi Corso di laurea specialistica: Decisioni economiche, impresa e responsabilità sociale, A.A. 2004/05 Soluzioni degli esercizi
Teoria dei Giochi, Trento, 2004/05 c Fioravante Patrone 1 Teoria dei Giochi Corso di laurea specialistica: Decisioni economiche, impresa e responsabilità sociale, A.A. 2004/05 Soluzioni degli esercizi
RAPPRESENTAZIONE DEI DATI
 Rappresentazione dei Dati RAPPRESENTAZIONE DEI DATI Quando si dispone di un alto numero di misure della stessa grandezza fisica è opportuno organizzarle in modo da rendere evidente Quandoil si loro dispone
Rappresentazione dei Dati RAPPRESENTAZIONE DEI DATI Quando si dispone di un alto numero di misure della stessa grandezza fisica è opportuno organizzarle in modo da rendere evidente Quandoil si loro dispone
Facciamo qualche precisazione
 Abbiamo introdotto alcuni indici statistici (di posizione, di variabilità e di forma) ottenibili da Excel con la funzione Riepilogo Statistiche Facciamo qualche precisazione Al fine della partecipazione
Abbiamo introdotto alcuni indici statistici (di posizione, di variabilità e di forma) ottenibili da Excel con la funzione Riepilogo Statistiche Facciamo qualche precisazione Al fine della partecipazione
Ing. Simone Giovannetti
 Università degli Studi di Firenze Dipartimento di Elettronica e Telecomunicazioni Ing. Simone Giovannetti Firenze, 29 Maggio 2012 1 Incertezza di Misura (1/3) La necessità di misurare nasce dall esigenza
Università degli Studi di Firenze Dipartimento di Elettronica e Telecomunicazioni Ing. Simone Giovannetti Firenze, 29 Maggio 2012 1 Incertezza di Misura (1/3) La necessità di misurare nasce dall esigenza
matematica probabilmente
 IS science centre immaginario scientifico Laboratorio dell'immaginario Scientifico - Trieste tel. 040224424 - fax 040224439 - e-mail: lis@lis.trieste.it - www.immaginarioscientifico.it indice Altezze e
IS science centre immaginario scientifico Laboratorio dell'immaginario Scientifico - Trieste tel. 040224424 - fax 040224439 - e-mail: lis@lis.trieste.it - www.immaginarioscientifico.it indice Altezze e
Capitolo 25: Lo scambio nel mercato delle assicurazioni
 Capitolo 25: Lo scambio nel mercato delle assicurazioni 25.1: Introduzione In questo capitolo la teoria economica discussa nei capitoli 23 e 24 viene applicata all analisi dello scambio del rischio nel
Capitolo 25: Lo scambio nel mercato delle assicurazioni 25.1: Introduzione In questo capitolo la teoria economica discussa nei capitoli 23 e 24 viene applicata all analisi dello scambio del rischio nel
Elementi di Psicometria con Laboratorio di SPSS 1
 Elementi di Psicometria con Laboratorio di SPSS 1 12-Il t-test per campioni appaiati vers. 1.2 (7 novembre 2014) Germano Rossi 1 germano.rossi@unimib.it 1 Dipartimento di Psicologia, Università di Milano-Bicocca
Elementi di Psicometria con Laboratorio di SPSS 1 12-Il t-test per campioni appaiati vers. 1.2 (7 novembre 2014) Germano Rossi 1 germano.rossi@unimib.it 1 Dipartimento di Psicologia, Università di Milano-Bicocca
Il mercato assicurativo: selezione avversa, fallimenti del mercato, menù di contratti, assicurazione obbligatoria
 Il mercato assicurativo: selezione avversa, fallimenti del mercato, menù di contratti, assicurazione obbligatoria Esercizio 1 Ci sono 2000 individui ciascuno con funzione di utilità Von Neumann-Morgestern
Il mercato assicurativo: selezione avversa, fallimenti del mercato, menù di contratti, assicurazione obbligatoria Esercizio 1 Ci sono 2000 individui ciascuno con funzione di utilità Von Neumann-Morgestern
Corso di Laurea in Scienze e Tecnologie Biomolecolari. NOME COGNOME N. Matr.
 Corso di Laurea in Scienze e Tecnologie Biomolecolari Matematica e Statistica II Prova di esame dell 11/1/2012 NOME COGNOME N. Matr. Rispondere alle domande nel modo più completo possibile, cercando di
Corso di Laurea in Scienze e Tecnologie Biomolecolari Matematica e Statistica II Prova di esame dell 11/1/2012 NOME COGNOME N. Matr. Rispondere alle domande nel modo più completo possibile, cercando di
CALCOLO COMBINATORIO
 CALCOLO COMBINATORIO 1 Modi di formare gruppi di k oggetti presi da n dati 11 disposizioni semplici, permutazioni Dati n oggetti distinti a 1,, a n si chiamano disposizioni semplici di questi oggetti,
CALCOLO COMBINATORIO 1 Modi di formare gruppi di k oggetti presi da n dati 11 disposizioni semplici, permutazioni Dati n oggetti distinti a 1,, a n si chiamano disposizioni semplici di questi oggetti,
Strutturazione logica dei dati: i file
 Strutturazione logica dei dati: i file Informazioni più complesse possono essere composte a partire da informazioni elementari Esempio di una banca: supponiamo di voler mantenere all'interno di un computer
Strutturazione logica dei dati: i file Informazioni più complesse possono essere composte a partire da informazioni elementari Esempio di una banca: supponiamo di voler mantenere all'interno di un computer
Regressione Mario Guarracino Data Mining a.a. 2010/2011
 Regressione Esempio Un azienda manifatturiera vuole analizzare il legame che intercorre tra il volume produttivo X per uno dei propri stabilimenti e il corrispondente costo mensile Y di produzione. Volume
Regressione Esempio Un azienda manifatturiera vuole analizzare il legame che intercorre tra il volume produttivo X per uno dei propri stabilimenti e il corrispondente costo mensile Y di produzione. Volume
La categoria «ES» presenta (di solito) gli stessi comandi
 gli stessi comandi") Utilizzo delle calcolatrici FX 991 ES+ Parte II PARMA, 11 Marzo 2014 Prof. Francesco Bologna bolfra@gmail.com ARGOMENTI DELLA LEZIONE 1. Richiami lezione precedente 2.Calcolo delle statistiche di regressione:
Utilizzo delle calcolatrici FX 991 ES+ Parte II PARMA, 11 Marzo 2014 Prof. Francesco Bologna bolfra@gmail.com ARGOMENTI DELLA LEZIONE 1. Richiami lezione precedente 2.Calcolo delle statistiche di regressione:
Misure della dispersione o della variabilità
 QUARTA UNITA Misure della dispersione o della variabilità Abbiamo visto che un punteggio di per sé non ha alcun significato e lo acquista solo quando è posto a confronto con altri punteggi o con una statistica.
QUARTA UNITA Misure della dispersione o della variabilità Abbiamo visto che un punteggio di per sé non ha alcun significato e lo acquista solo quando è posto a confronto con altri punteggi o con una statistica.
I marcatori molecolari. Dipartimento di Scienze Agronomiche e Genetica Vegetale Agraria Corso di Genetica Agraria Giovanna Attene
 I marcatori molecolari Dipartimento di Scienze Agronomiche e Genetica Vegetale Agraria Corso di Genetica Agraria Giovanna Attene Marcatori molecolari del DNA I marcatori molecolari sono sequenze di DNA
I marcatori molecolari Dipartimento di Scienze Agronomiche e Genetica Vegetale Agraria Corso di Genetica Agraria Giovanna Attene Marcatori molecolari del DNA I marcatori molecolari sono sequenze di DNA
 Stima per intervalli Nei metodi di stima puntuale è sempre presente un ^ errore θ θ dovuto al fatto che la stima di θ in genere non coincide con il parametro θ. Sorge quindi l esigenza di determinare una
Stima per intervalli Nei metodi di stima puntuale è sempre presente un ^ errore θ θ dovuto al fatto che la stima di θ in genere non coincide con il parametro θ. Sorge quindi l esigenza di determinare una
Leasing secondo lo IAS 17
 Leasing secondo lo IAS 17 Leasing: Ias 17 Lo Ias 17 prevede modalità diverse di rappresentazione contabile a seconda si tratti di leasing finanziario o di leasing operativo. Il leasing è un contratto per
Leasing secondo lo IAS 17 Leasing: Ias 17 Lo Ias 17 prevede modalità diverse di rappresentazione contabile a seconda si tratti di leasing finanziario o di leasing operativo. Il leasing è un contratto per
Aspettative, consumo e investimento
 Aspettative, consumo e investimento In questa lezione: Studiamo come le aspettative di reddito e ricchezza futuro determinano le decisioni di consumo e investimento degli individui. Studiamo cosa determina
Aspettative, consumo e investimento In questa lezione: Studiamo come le aspettative di reddito e ricchezza futuro determinano le decisioni di consumo e investimento degli individui. Studiamo cosa determina
Basi di matematica per il corso di micro
 Basi di matematica per il corso di micro Microeconomia (anno accademico 2006-2007) Lezione del 21 Marzo 2007 Marianna Belloc 1 Le funzioni 1.1 Definizione Una funzione è una regola che descrive una relazione
Basi di matematica per il corso di micro Microeconomia (anno accademico 2006-2007) Lezione del 21 Marzo 2007 Marianna Belloc 1 Le funzioni 1.1 Definizione Una funzione è una regola che descrive una relazione
Introduzione agli Algoritmi Genetici Prof. Beatrice Lazzerini
 Introduzione agli Algoritmi Genetici Prof. Beatrice Lazzerini Dipartimento di Ingegneria della Informazione Via Diotisalvi, 2 56122 PISA ALGORITMI GENETICI (GA) Sono usati per risolvere problemi di ricerca
Introduzione agli Algoritmi Genetici Prof. Beatrice Lazzerini Dipartimento di Ingegneria della Informazione Via Diotisalvi, 2 56122 PISA ALGORITMI GENETICI (GA) Sono usati per risolvere problemi di ricerca
LA STATISTICA si interessa del rilevamento, dell elaborazione e dello studio dei dati; studia ciò che accade o come è fatto un gruppo numeroso di
 STATISTICA LA STATISTICA si interessa del rilevamento, dell elaborazione e dello studio dei dati; studia ciò che accade o come è fatto un gruppo numeroso di oggetti; cerca, attraverso l uso della matematica
STATISTICA LA STATISTICA si interessa del rilevamento, dell elaborazione e dello studio dei dati; studia ciò che accade o come è fatto un gruppo numeroso di oggetti; cerca, attraverso l uso della matematica
Funzioni. Funzioni /2
 Funzioni Una funzione f è una corrispondenza tra due insiemi A e B che a ciascun elemento di A associa un unico elemento di B. Si scrive: f : A B l'insieme A si chiama il dominio della funzione f, l'insieme
Funzioni Una funzione f è una corrispondenza tra due insiemi A e B che a ciascun elemento di A associa un unico elemento di B. Si scrive: f : A B l'insieme A si chiama il dominio della funzione f, l'insieme
Gestione della politica monetaria: strumenti e obiettivi corso PAS. Mishkin, Eakins, Istituzioni e mercati finanziari, 3/ed.
 Gestione della politica monetaria: strumenti e obiettivi corso PAS 1 Anteprima Con il termine politica monetaria si intende la gestione dell offerta di moneta. Sebbene il concetto possa apparire semplice,
Gestione della politica monetaria: strumenti e obiettivi corso PAS 1 Anteprima Con il termine politica monetaria si intende la gestione dell offerta di moneta. Sebbene il concetto possa apparire semplice,
Analisi bivariata. Dott. Cazzaniga Paolo. Dip. di Scienze Umane e Sociali paolo.cazzaniga@unibg.it
 Dip. di Scienze Umane e Sociali paolo.cazzaniga@unibg.it Introduzione : analisi delle relazioni tra due caratteristiche osservate sulle stesse unità statistiche studio del comportamento di due caratteri
Dip. di Scienze Umane e Sociali paolo.cazzaniga@unibg.it Introduzione : analisi delle relazioni tra due caratteristiche osservate sulle stesse unità statistiche studio del comportamento di due caratteri