Relatore: Prof.ssa Alessandra Piras. Tesi di laurea di: Salvatore Virdis
|
|
|
- Geraldina Viola
- 6 anni fa
- Visualizzazioni
Transcript
1 Relatore: Prof.ssa Alessandra Piras Tesi di laurea di: Salvatore Virdis Anno Accademico
2 SOMMARIO SOMMARIO... 2 INTRODUZIONE... 4 VIVIPARITà E PLACENTAZIONE NELLE DIVERSE CLASSI DI VERTEBRATI... 5 TIPI DI PLACENTE NEI MAMMIFERI NUTRIZIONE ISTIOTROFICA ED EMOTROFICA CIRCOLAZIONE SANGUIGNA PLACENTARE NEI MAMMIFERI FISIOLOGIA COMPARATA PLACENTARE LA PLACENTA UMANA CARATTERISTICHE MACROSCOPICHE DELLA PLACENTA UMANA IMPIANTO DELLA PLACENTA UMANA STRUTTURA E ULTRASTRUTTURA DEL VILLO CORIALE UMANO SINCIZIOTROFOBLASTO CITOTROFOBLASTO LAMINA BASALE DEL TROFOBLASTO ASSE MESENCHIMALE DEL VILLO CELLULE DELL'ASSE MESENCHIMALE SOSTANZA INTERCELLULARE DELL'ASSE MESENCHIMALE CAPILLARI FETALI PERICITI DEI CAPILLARI FETALI ENDOTELIO DEI CAPILLARI FETALI FISIOLOGIA DELLA PLACENTA UMANA CIRCOLAZIONE PLACENTARE BARRIERA PLACENTARE E TRASPORTO ATTRAVERSO LA PLACENTA TRASPORTO ATTRAVERSO LA PLACENTA ACQUA ELETTROLITI NUTRIZIONE DEL FETO AMMINOACIDI, POLIPEPTIDI E PROTEINE LIPIDI GLUCIDI LA PLACENTA COME ORGANO DI ESCREZIONE ORMONI ORMONI STEROIDEI VITAMINE
3 FARMACI IMMUNOLOGIA MATERIALI E METODI RISULTATI OSSERVAZIONI AL MICROSCOPIO OTTICO OSSERVAZIONI AL MICROSCOPIO ELETTRONICO CONCLUSIONI MATERIALE ICONOGRAFICO
4 INTRODUZIONE La placenta umana é un organo temporaneo, il quale partecipa a costituire un' entità anatomo-funzionale inscindibile, denominata "unita feto-placentare". Già da tempi remoti, non soltanto nella specie umana, la placenta ha assunto un'importanza notevole nella considerazione degli studiosi, per il fatto di essere un organo intermediario tra organismo materno e organismo fetale. Fino a qualche tempo fa la placenta veniva esclusivamente considerata come un organo che passivamente mediava gli scambi metabolici e le informazioni tra madre e feto, senza prendere parte attiva a questi scambi. L'individuazione nella placenta di una funzione endocrina ha aperto un nuovo capitolo nella comprensione di quest'organo che può esercitare sul metabolismo materno-fetale, proprio grazie alla sua attività endocrina, un'importante controllo. Comunque la più evidente delle funzioni placentari é quella che concerne gli scambi metabolici tra madre e feto; a livello di quest'organo infatti si creano le strutture che permettono una strettissima vicinanza tra sangue fetale e sangue materno, pur non realizzandosi un mescolamento. A livello di quella che e stata definita" barriera placentare" si individuano strutture altamente specializzate che selezionano le sostanze che possono passare da madre a feto e viceversa. In questo processo di controllo svolgerebbe un ruolo fondamentale la membrana basale del trofoblasto. Verranno di seguito esposti i dati bibliografici riguardanti la placenta nelle varie specie animali, quindi i dati sulla struttura, ultrastruttura e funzioni principali della placenta umana e infine i dati sperimentali riguardanti le ricerche oggetto della presente tesi. 4
5 VIVIPARITà E PLACENTAZIONE NELLE DIVERSE CLASSI DI VERTEBRATI I mammiferi, nei corso dell'evoluzione, sono passati dalla condizione di oviparità a quella di viviparità. Lo sviluppo dell'embrione, che primitivamente avveniva al di fuori del corpo materno, gradualmente comincia a localizzarsi nell'utero a spese di sostanze fornite dalla madre. Parimenti anche la cellula uovo, originariamente ricca di vitello, si e modificata riducendo il deutoplasma divenuto superfluo. Nelle tre sottoclassi dei mammiferi attuali, Monotremi, Marsupiali e Placentati, si ritrovano le tappe di questa evoluzione che comporta necessariamente anche un diverso sviluppo degli annessi fetali. Prima di giungere a questa condizione abbiamo tutta una serie di rapporti più o meno stretti tra madre e feto che si sviluppano in categorie sistematiche considerate inferiori rispetto all'adattamento alla vita sulla terra. Nel corso dell'evoluzione, le specie vivipare hanno preso origine da antenati ovipari come adattamento a tempi progressivamente più lunghi tra fecondazione e covata delle uova. Per raggiungere questa tappa evolutiva si e avuta l'esigenza di sviluppare determinati annessi embrionali (schema 1). Ciclostomi, pesci e anfibi non possiedono l'amnios e sono detti anamni. Negli embrioni di questi vertebrati, che presentano soltanto la condizione di oviparità, il solo annesso embrionale sviluppato é il sacco vitellino trilaminare costituito dai tre foglietti embrionali ecto, meso ed endoderma. I1 mesoderma diviene molto vascolarizzato e trasporta il materiale nutritivo, assorbito dal vitello mediante l'endoderma, ai tessuti in crescita dell'embrione. Man mano che il corpo cresce, il sacco vitellino si retrae e finalmente si incorpora nella parete addominale ventrale e nell'intestino. In natura esiste tutta una serie di modificazioni rispetto a questo tipo di sviluppo schematizzato. Per esempio, negli Elasmobranchi (squali, razze), lo squilibrio osmotico fra ambiente marino ipertonico e liquidi organici viene compensate con concentrazioni di urea superiori a quelle degli altri vertebrati. A differenza delle specie ovipare, in cui 1'uremia è mantenuta da un rivestimento impermeabile dell'uovo, nello squalo viviparo spinarolo (Squalus acanthias), e la 5
6 madre che nei primi sei mesi di gestazione mantiene la concentrazione di urea nei liquidi uterini (H0GART, 1976). Successivamente l'embrione é in grado di provvedere autonomamente a questa esigenza. Nella specie suddetta, durante la gestazione, che dura da 18 a 22 mesi, la base degli ovidutti modificata a formare un utero, subisce notevoli trasformazioni. L'utero non gravido é rivestito da un singolo strato di epitelio colonnare scarsamente vascolarizzato. Durante la gestazione tale epitelio si ipertrofizza e diviene fortemente convoluto, incrementando la sua area superficiale alcune centinaia di volte. Anche nella Razza vivipara (Urolophus halleri), si ha un incremento dell'area interna dell'utero per la presenza di molti piccoli lembi di tessuto che aumentano considerevolmente in seguito allo sviluppo embrionale intrauterino. Gli scambi gassosi tra madre ed embrione nelle specie vivipare del notidiano cinereo (Heptranchias perlo) e della razza (Urolophus), sono facilitati da un gran numero di filamenti branchiali o da un grande accrescimento delle branchie esterne. Questi adattamenti indicano una bassa concentrazione di ossigeno nell'utero, suggerita in alcune specie da un'emoglobina fetale con maggior affinità per l'ossigeno (GRASSE', 1958). In alcuni casi i capillari vitellini possono assorbire nutrienti, sia dalla massa del tuorlo che dalle sostanze disciolte nei fluidi uterini. Questa onfaloplacenta è stata studiata nello squalo martello (Spyrna tiburo). Circa trecinque mesi dopo la fecondazione, il sacco del tuorlo prende contatto con la parete dell utero e le due superfici si accrescono sotto forma di lembi che si interdigitano sempre più saldamente man mano che il tuorlo si esaurisce. Anche l'approvvigionamento di sangue viene incrementato. Dall'intimità delle relazioni strutturali tra madre e feto, possiamo attenderci strette relazioni metaboliche (GRASSE',1958),(Fig.1). Nei Teleostei vivipari (pesci ossei) la gestazione può essere follicolare od ovarica. Nella prima, le uova sono fecondate all'interno dei follicoli ovarici e lo sviluppo embrionale continua nel follicolo fino all'ovulazione che precede immediatamente la nascita. Questa gestazione follicolare e tipica della famiglia dei Poeciliidae come nel "pesce milione" (Poecilia reticulata) e nei "pesci portaspada" (Xifoforus helleri e X. maculatus). 6
7 7
8 Nella gestazione ovarica le uova sono ovulate nel lume della cavità ovarica e all'interno di essa avviene lo sviluppo embrionale. Un esempio di quanto detto si ha in Zoarces viviparus. Inoltre vi sono pesci che manifestano nelle prime fasi una gestazione follicolare ed in seguito una gestazione ovarica. Nei Poeciliidae, la modificazione strutturale che si riscontra rispetto agli embrioni delle specie ovipare, é la grande espansione del sacco pericardico in due frange che si accrescono sulla parte anteriore dell'embrione e si fondono insieme (fig. 2). Questa struttura, per via d i una ricca vascolarizzazione e per la funzione di scambio di materiali tra madre e feto é stata chiamata dai biologi "pseudoplacenta". Tale annesso embrionale consente il passaggio di ossigeno e acqua e la rimozione di residui azotati e anidride carbonica dall'ambiente che circonda l'embrione. Sono scarse le prove dirette circa il passaggio di materiali organici nell'embrione durante la gestazione. Sostanze a basso peso molecolare marcate radioattivamente (glucosio e amminoacidi), iniettate in femmine gravide del "pesce milione" o del "pesce portaspada" vengono rinvenute solo in tracce nel feto. Composti ad alto peso molecolare, come il complesso "Myofer" di ferro-destrani, possono passare nel feto di Xifophorus abbastanza liberamente; questo fa supporre un processo selettivo nel trasferimento di materiale da madre a feto (HOGARTH, 1976). Rimanendo tra i pesci ossei, nella famiglia dei Jensiidae, le uova sono fecondate all'interno dei follicoli ovarici. Inizialmente, la gestazione é follicolare e si forma una pseudoplacenta pericardiale. Terminate le scorte di tuorlo, l'embrione perde la sua pseudoplacenta ed é ovulato nei lume dell'ovario dove continua il suo sviluppo. L'epitelio di rivestimento del lume ovarico si accresce esternamente in un certo numero di lembi irregolari che attraversano le fenditure branchiali del feto e penetrano nella sua faringe. Presumibilmente il nutrimento é trasportato per questo mezzo direttamente nell'intestino del feto (TURNER, 1940). Nei pesci appartenenti alla famiglia dei Godeidae, si ha una situazione simile che differisce dalla precedente per il fatto che é l'embrione e non l'ovario a formare speciali organi di assorbimento. Si osservano dei filamenti nastriformi trofici che 8
9 crescono in maggior misura nella parte posteriore dell'intestino embrionale. In alcuni membri primitivi della famiglia essi assolvono una funzione respiratoria mentre nelle specie più evolute hanno un ruolo nutrizionale (TURNER, 1940), (fig. 3a). Nella famiglia degli Embiotocidae vivipari, il peso secco dell'embrione durante lo sviluppo aumenta approssimativamente 17 volte. Sia l'ovario che l'embrione mostrano speciali adattamenti per facilitare l'interscambio di diverse sostanze. Le pareti ovariche aumentano la propria superficie durante la gravidanza e divengono altamente vascolarizzate e secernenti. Nel tardo periodo della gestazione le pieghe della parete possono penetrare nella cavità branchiale del feto e venire in diretto contatto con le branchie. Simultaneamente si assiste ad un incremento della superficie assorbente dell'embrione con estensioni a forma di spatola dei tessuti delle pinne mediane e caudali (WEBB e BRETT, 1972), (fig. 3b). Una delle forme più bizzarre di viviparità é quella del genere Hippocampus o cavalluccio marino nei quale l'incubazione delle uova é portata a termine dal maschio. Le uova deposte dalle femmine sono fecondate e accolte in una speciale tasca da covata. All'interno di ciascuna tasca possono essere contenute circa 200 uova, (fig. 4). In alcune specie di Hippocampus come anche nei pesci Nerophis e Signatus la superficie della tasca si trasforma in una struttura placentare con un ricco apporto di sangue ed i n alcuni casi una secrezione nutritiva (GRASSE',1958). Anche in alcuni pesci in cui la viviparità si sviluppa nella cavità buccale come nel pesce gatto (Gateichtys), abbiamo una modificazione nella struttura epiteliale che assume la funzione di una pseudoplacenta. Infatti l epitelio secerne i nutrienti necessari agli embrioni e assolve anche una funzione battericida. La secrezione dei carboidrati e stata individuata per via istochimica. Gli Anfibi esistenti sono suddivisi i n tre gruppi i quali presentano forme vivipare. Tra gli Urodeli, la salamandra nera (Salamandra atra) è l unica che genera due piccoli dopo un periodo di gestazione di circa un anno. L'unico adattamento morfologico alla viviparità e che le branchie esterne degli embrioni differiscono da quelle di individui allo stesso stadio delle specie ovipare. Questo indica una funzione assorbente accresciuta. Il solo anuro che mostra una vera viviparità é la 9
10 piccola rana (Nectophrynoides occidentalis). Questo anflbio vive ad altitudini di 1200 metri, nell Africa Occidentale, in un'area con stagioni alternativamente umide e secche. 10
11 11
12 La fecondazione avviene alla fine di una stagione umida e la nascita alla metà della successiva; ciò suggerisce che la viviparità é un adattamento per evitare l'esposizione delle uova a condizioni secche durante lo sviluppo (HOGARTH, 1976). Nelle regioni interne dell'ovidutto sono presenti villi vascolari e negli embrioni la coda é slargata, presumibilmente come organo di assorbimento. Gli ovari, come quelli di altri vertebrati, contengono strutture che ricordano i corpi lutei dei Mammiferi e che sembrano essere in grado di sintetizzare progesterone. Probabilmente sono stimolati dalla prolattina secreta dalla ghiandola pituitaria. In un altro Anuro, del genere Gastrotheca, le uova sono portate in una tasca formata sul dorso della femmina. Le larve possiedono branchie molto espanse a forma di campana, cosi grandi da circondare completamente gli embrioni e applicate a creste vascolari all'interno della tasca marsupiale (NEEDHAM, 1950), (fig. 5). La maggior parte degli Anamni di cui si é parlato sopra depone le proprie uova nell'acqua, ma vi sono altri gruppi di vertebrati megalecitici (Rettili, Uccelli e Monotremi) che depongono le uova sulla terra. Sono tutti caratterizzati dalla presenza di un amnios, costituito internamente da ectoblasto extraembrionale rivestito esternamente da somatopleura mesodermica, che forma una cavità ripiena di liquido che circonda l'embrione. Questa cavità ha la funzione di permettere lo sviluppo embrionale al di fuori dell'acqua, mantenendo umido l'embrione e proteggendolo dalle scosse e aderenze. Il corion, o sierosa, si forma durante la chiusura delle pieghe amniotiche e risulta delimitato esternamente dall'ectoblasto extraembrionale e internamente dalla somatopleura. Lo sviluppo del celoma extraembrionale separa completamente o incompletamente la somatopleura extraembrionale (amnios e corion) dalla splancnopleura extraembrionale (sacco vitellino viscerale). Anche nei Rettili abbiamo delle forme di pseudoplacentazione più o meno evolute. 12
13 13
14 Pochi rettili, ad esempio certi serpenti e lucertole come la lucertola vivipara (Lacerta vivipara) e la vipera comune (Vipera aspis) sono ovovivipare. Le uova vengono mantenute nell'ovidutto fino alla schiusa. In questi l unica relazione tra le uova e l ovidutto sono gli scambi gassosi. Un gran numero di rettili vivipari ha un sacco del tuorlo che permette l assunzione di diverse sostanze dai fluidi uterini. Nei serpenti, come il serpente giarrettiera (Tamnophis sirtalii) e nelle lucertole, come lo scinco europeo (Calcides tridactilus), il sacco del tuorlo viene a contatto con il rivestimento uterino e forma un onfaloplacenta. I tessuti embrionali e materni formano delle pieghe interdigitanti che sono separate solo da una membrana dell uovo molto sottile (H0GARTH, 1976). Oltre all'onfaloplacenta descritta, in questo raggruppamento troviamo placente corio-allantoidee (WEEKES,1930 e 1935). Queste differiscono da specie a specie. In alcuni casi la placenta può avere spessore ridotto sicché i vasi della madre e del feto risultano in intimo contatto. In altri casi i capillari materni sono rigonfi e l epitelio materno é chiaramente secernente, mentre le cellule corioidee possono slargarsi ed in alcuni casi formano delle estroflessioni che penetrano fra le cellule dell epitelio uterino. In Thamnophis, i nuovi nati presentano un contenuto in proteine più elevato rispetto a quello presente nelle uova durante il periodo della fecondazione. Glicina marcata radioattivamente e iniettata in una femmina gravida compare dopo un breve periodo di tempo nell'embrione. Oltre alla glicina si ha il passaggio di ioni sodio e molecole più grandi come il blu di metilene (peso molecolare 320). 14
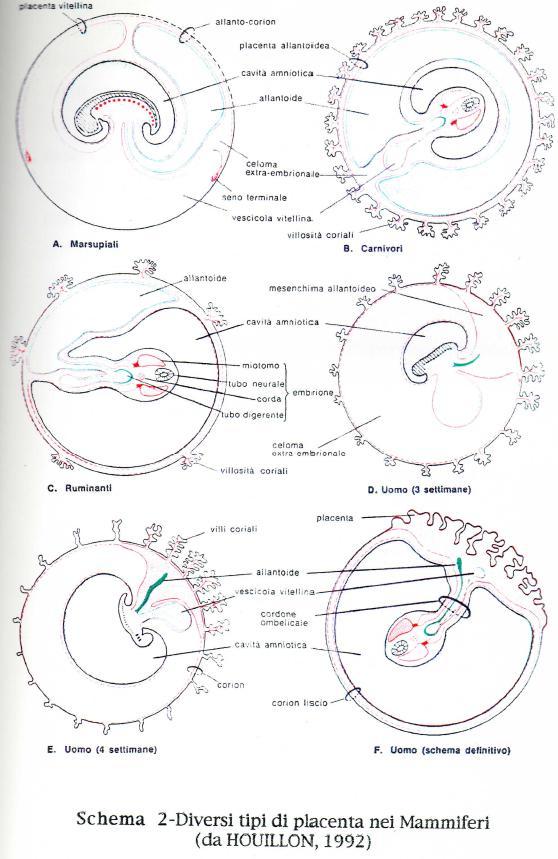
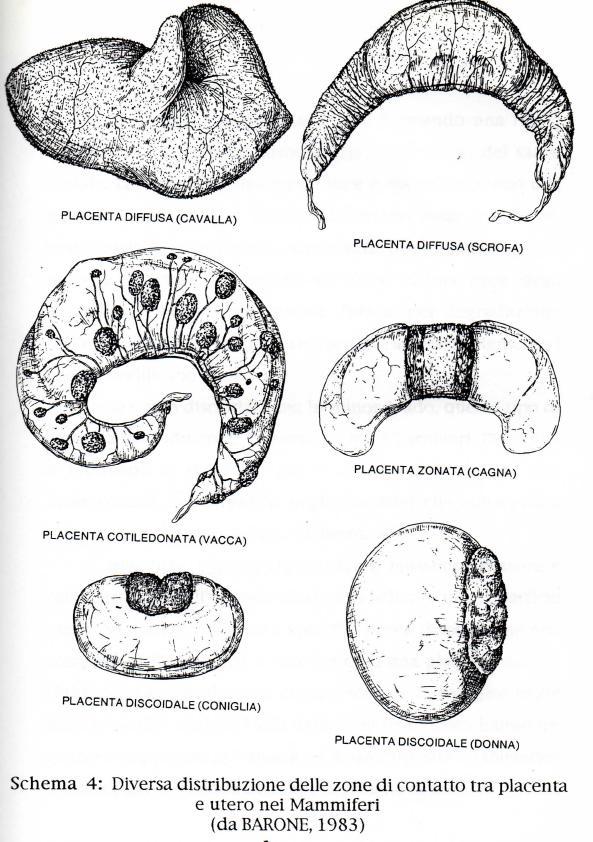
15 TIPI DI PLACENTE NEI MAMMIFERI Lo sviluppo dei Mammiferi è diverso da quello degli altri vertebrati per il particolare grado di adattamento raggiunto da questa classe. Alcune caratteristiche riguardanti la zona di sviluppo embriogenetica, nell'uovo allo stadio di blastocisti, può considerarsi come una delle tante prove di discendenza filogenetica di questo gruppo da antenati molto vicini ai rettili primitivi. Tuttavia lo sviluppo di particolari annessi embrionali, quali la placenta, e lo sviluppo intrauterino hanno determinato la perdita del vitello delle uova dei mammiferi, utilizzato come nutrimento per il compimento dello sviluppo embrionale dagli animali ovipari (HOUILLON, 1992). Nelle uova dei Marsupiali, diversamente dai Rettili, non si riscontra la membrana testacea, e perciò il corion può venire a diretto contatto con i tessuti materni. I Marsupiali, con periodo di gestazione più breve, per esempio l Opossum (Didelphys) ed i l Canguro (Macropus), sembrano dipendere completamente dal sacco vitellino nelle loro relazioni con l'utero, mentre l allantoide assume un significato respiratorio. L'onfaloplacenta trilaminare costituisce un organo importante per la nutrizione e la respirazione. I Marsupiali con periodi di gestazione più lunga, come il Parameles (ratto canguro), hanno una placenta corio-allantoidea di complessita pressappoco simile a quella più altamente specializzata delle lucertole (HILL, 1925; FLYNN, 1923 e 1930; PEARSON, 1949). La placenta é un organo caratteristico dei Mammiferi Euteri e deriva dalla compenetrazione ed unione del corion con la mucosa uterina. Si conoscono quattro diversi tipi di placenta: diffusa, cotiledonare, zonata e discoidale. La placenta diffusa risulta una forma primitiva; le altre placente presentano un rapporto più intimo fra le parti fetali e materne. La placenta é un annesso embrionale proprio delle specie animali vivipare, per mezzo della quale l'embrione stabilisce contatti diretti con il corpo materno che gli fornisce il nutrimento. Nei mammiferi la placenta e essenzialmente di tre tipi. Nel primo tipo si stabilisce una connessione tra la parete uterina e quella del corion a cui si addossa il sacco del tuorlo ricco di vasi vitellini. Questo tipo di placenta é detta corio-vitellina, e si trova in alcuni Marsupiali (Didelphys, 15
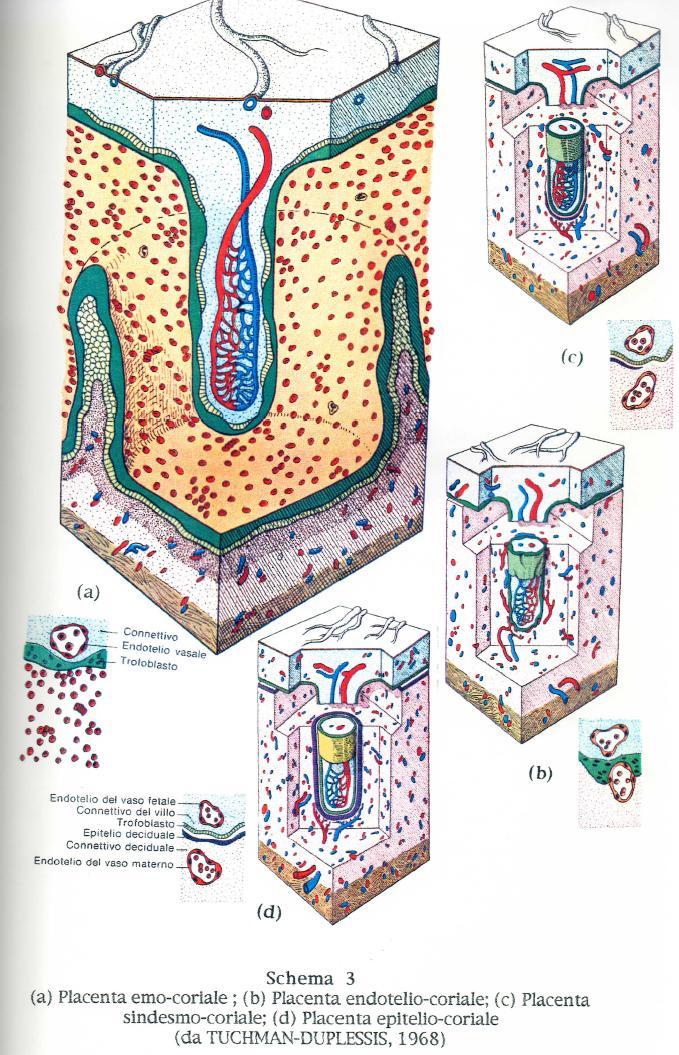
16 Macropus). Nel secondo tipo, la connessione con la parete uterina é stabilita da quella parte del corion a cui si addossa l'allantoide, che ne induce la trasformazione in corion frondoso. In questo caso sono i vasi allantoidei che trasportano le sostanze dalla madre all'embrione. Questo tipo d i placenta è detta corio-allantoidea, e si trova in tutti gli Euteri come pure in alcuni Marsupiali (Parameles, Dasyurus). II terzo tipo è la placenta anallantoidea che corrisponde a quella dell Uomo, dei Primati etc, in cui i vasi coriali si sviluppano, per induzione temporanea da parte dell'allantoide, nel peduncolo mesenchimatico allantoideo. Nei Mammiferi superiori si possono trovare residui di placenta corio-vitellina sia per un breve periodo che permanentemente; essi hanno una funzione sussidiaria, (Schema 2). Quando l'embrione dei mammiferi giunge nell'utero, la zona pellucida che lo circondava, precocemente si dissolve e l'embrione, ora allo stadio di blastocisti é bagnato dal liquido uterino. Questo liquido, prodotto dalle ghiandole tubulari della parete uterina é versato nel lume dell utero e contiene sostanze organiche. II giovane embrione può assorbire alcune di queste sostanze attraverso il trofoblasto protettivo finche non si sia stabilita una più stretta connessione con la parete uterina. II suo ulteriore sviluppo dipende completamente dalle sostanze fornitegli per diffusione dai tessuti della parete uterina. Affinché queste sostanze fluiscano in modo continuo e abbondante é essenziale uno stretto rapporto t r a i tessuti embrionali ed i tessuti materni. Tuttavia si e osservato che queste connessioni più o meno strette tra i tessuti materni e fetali possono essere diverse entro la classe dei mammiferi ed anche entro la stessa sottoclasse dei veri mammiferi placentati (euteri). In base a questo tipo di connessioni ed in base agli strati tissutali che formano la barriera placentare, nei 1909 Grosser classificò la placenta in quattro tipi, ai quali Mosmann nel 1926 ne aggiunse un quinto. (a) epitelio-coriale a sei strati (cavallo, maiale, cammello...etc.) (b) sindesmo-coriale a cinque strati con assenza dell'epitelio uterino (Ruminanti) (c) endotelio-coriale a quattro strati, con perdita dell'epitelio della lamina propria della mucosa uterina (Carnivori); (d) emo-coriale a tre strati, tutti di pertinenza fetale (manca anche l'endotelio dei capillari materni (Primati, Uomo, Insettivori, Chirotteri); 16
17 17
18 (e) emo-endocoriale a un solo strato, rappresentato dall'endotelio dei capillari coriali (Roditori, Lagomorfi). La suddetta classificazione, basata su osservazioni al microscopio ottico, talvolta si é rivelata non corrispondente al vero e troppo approssimativa nella rigida elencazione degli strati. Precedenti studi effettuati al microscopio elettronico ascrivono la placenta del bovino e della pecora (e dei Ruminanti in genere) al tipo epitelio-coriale, anche se con il procedere della gravidanza, in aree più o meno limitate della mucosa uterina l epitelio può cadere (BARONE, 1983). Anche nella tipica placenta epitelio-coriale del cavallo, a livello della corona coriale, zona coriale interposta tra encefalo-corion e allanto-corion, cade l'epitelio uterino con conseguente formazione delle coppe endometriali. Alle placente di tipo emo-coriale vengono oggi riportate anche quelle del coniglio e della cavia, ritenute un tempo di tipo emo-endoteliocoriale, in quanto é risultato che l'endotelio dei capillari fetali non é a diretto contatto con il sangue materno. La classificazione proposta da Steven nei 1975 raggruppa le placente nei soli tre tipi: a) epitelio-coriale (Artiodattili, Perissodattili, Cetacei, Lemuridi); b) endotelio-coriale (Carnivori pressoché tutti, Pinnipedi); c) emo-coriale (Insettivori, Roditori, Lagomorfi, Sirenidi, Primati). Per ciò che riguarda l'organizzazione istologica si può dire che il corion, conservando generalmente la sua integrità, presenta con l'endometrio rapporti che vanno dalla semplice assimilazione, all'erosione più o meno profonda, dapprima dell epitelio poi della lamina propria della mucosa uterina ed infine dei vasi materni stessi (Schema 3). La placenta epitelio-coriale é il tipo più primitivo che si accompagna con una placenta diffusa. Tra le specie domestiche é presente negli Equidi, nei Suidi e nella Vacca. L'epitelio uterino connesso con questo tipo d i placenta si mantiene nella sua integrità. Si osserva dunque una semplice opposizione di parti coriali e uterine diversamente ingrossate ed é perciò denominata semiplacenta. 18
19 La parte fetale e caratterizzata da assenza di sinciziotrofoblasto. È rivestita dal citotrofoblasto costituito da un unico strato di cellule batiprismatiche, sotto il quale si osserva un connettivo molto vascolarizzato. 19
20 20
21 II tutto costituisce i villi corioallantoidei semplici appena ramificati, raggruppati in folti ciuffi, ciascuno dei quali sorretto da un peduncolo. Quest'ultimo costituisce negli Equidi un microcotiledone. Tale raggruppamento di villi é particolare dei Suidi nei quali determina pieghe complesse. La parte materna non é altro che lo strato superficiale della mucosa uterina, un epitelio cubico continuo sia in superficie, sia nel fondo delle ghiandole uterine. Queste ultime differiscono in disposizione a seconda delle specie. Negli Equidi sono molto sviluppate fin dal principio della gravidanza e, riunite a gruppi, sboccano nei lume uterino in minuscole depressioni infundiboliformi formando delle cripte più o meno profonde. Tale organizzazione e più diffusa nei Suidi. Nelle caruncole della Vacca non esistono delle vere ghiandole ma soltanto delle cripte profonde. II rapporto tra le due parti varia a seconda della specie. II più semplice lo abbiamo negli Equidi dove ogni cripta uterina riceve un microcotiledone; è separata dalle sue vicine da travate di connettivo che portano la rete vascolare materna. I due epiteli, materno e trofoblastico, non sono in contatto ovunque: rimangono separati nell'intervallo tra i cotiledoni mediante degli interstizi: le areole o "microlacune embriotrofiche", nelle quali si raccoglie il latte uterino o embriotrofo; d'altra parte, i villi non raggiungono il fondo delle ghiandole, che é occupato da una modesta quantità di secreto. La disposizione é molto più irregolare ed alquanto complicata nella Scrofa, ma il contatto tra i due epiteli rimane ancora molto incompleto, pur essendo essi sempre perfettamente continui. Durante la gravidanza si formano pure delle zone di distacco e l'embriotrofo si accumula negli interstizi separando gli epiteli. In ogni caso, le zone di contatto fra il trofoblasto e l'epitelio uterino presentano un ingranamento reciproco e stretto di microvilli delle superfici cellulari. Abbiamo inoltre delle differenze istologiche locali proprie di ciascuna specie, (Schema 3 d). La placenta sindesmo-coriale é presente nei Ruminanti con placenta cotiledonare. È caratteristica della Pecora e della Capra mentre è presente in modo incostante, tardivo e probabilmente accidentale nella Vacca. È una via di mezzo tra le placente adeciduate e le deciduate. La placenta é suddivisa in masse isolate, arrotondate e ovoidali, e spesso di numero elevato, ognuna delle quali é 21
22 detta placentoma. Questo é unito alla parete uterina mediante il peduncolo della caruncola materna. II diametro dei placentomi può raggiungere e anche superare sette o otto centimetri (Vacca) sulla parte media del conceptus, ma decresce molto verso le estremità. Ogni placentoma comporta una caruncola uterina sulla quale é in qualche modo modellato e ingranato un cotiledone fetale. La caruncola e formata da una giustapposizione di profonde cripte, ognuna delle quali riceve un microcotiledone mentre il cotiledone non é che un voluminoso raggruppamento di microcotiledoni. II trofoblasto e sempre costituito da cellule cubiche e presenta inoltre cellule giganti particolari. L'epitelio materno subisce un'erosione più o meno estesa sulla sommità delle travate che separano le cripte. Questa erosione é molto limitata o addirittura manca nella Vacca dove praticamente la struttura é ancora del tipo epiteliocoriale. La distruzione dell'epitelio materno e più estesa nella Pecora e nella Capra. Ne risulta che il trofoblasto viene direttamente in contatto con la lamina propria della mucosa uterina dove ha sede la rete capillare materna. Questo infatti corrisponde ad uno stato di transizione verso il tipo emocoriale. In corrispondenza delle zone desquamate esistono delle lacune topograficamente simili alle microlacune embriotrofiche degli Equidi ma nelle quali si accumulano piccole quantità di sangue stravasato dai capillari materni. Queste "lacune di riassorbimento sanguigno", più sviluppate nella Pecora che nella Capra, sono bordate da un epitelio più alto, differenziato in vista di un assorbimento attivo. Dal complesso di queste disposizioni deriva che al momento della nascita una distruzione ridotta dell'endometrio segnerà la separazione delle due parti della placenta (Schema 3 c). Nella placenta endotelio-coriale é presente un sinciziotrofoblasto che intacca profondamente l'endometrio. Perciò le placente di questo tipo sono dette placente vere in contrapposizione alle semiplacente viste precedentemente. La struttura endoteliocoriale é quella delle placente zonate quasi particolare dei Carnivori. Tuttavia sono anche osservate in alcuni Insettivori (Talpe) e Tardigradi (Bradipo). Ancora prima dell'impianto, la parte profonda delle ghiandole uterine si dilata e si ramifica (strato spongioso), mentre l'epitelio delle parti superficiali (strato 22
23 compatto dell'endometrio) si assottiglia molto. Questo epitelio viene ben presto riassorbito e i villi coriali, molto robusti, penetrano sia nello stroma dell'endometrio sia nella zona dilatata delle ghiandole, di cui non saranno rispettate neanche le parti più profonde. In seguito, i villi vengono rimaneggiati in modo da formare una rete complessa, mentre il sinciziotrofoblasto prolifera tra le ghiandole uterine fino a riassorbire quasi completamente il connettivo dell'endometrio ed inglobare la sua rete capillare, che é la sola ad essere risparmiata. Così l'endometrio vascolare materno viene a trovarsi direttamente in contatto con il trofobasto. Sono presenti inoltre piccole lacune di riassorbimento sanguigno, dove il sangue è stravasato dai capillari materni e assorbito da particolari villi del corion. Al momento del parto, la separazione avviene a livello dello strato spongioso, che assicura successivamente la ricostruzione dell'endometrio (schema 3b). Si può inoltre collegare al tipo endoteliocoriale la placenta detta endotelioendoteliale di certi topi ragno. In questi animali, i villi del corion fetale si approfondano ugualmente nell'endometrio fino a giungere in contatto con i capillari materni. Ma qui il trofoblasto è ridotto ad un sottile strato sinciziale nel quale al microscopio elettronico si evidenzia la presenza di fenestrature. Teoricamente tra il sangue materno e fetale non vi é quindi altra barriera che la parete delle reti capillari. La placenta emo-coriale è presente nei Gliri, negli Insettivori, nei Chirotteri e nei Primati; nel tipo strutturale discoidale comporta la più profonda e intima unione tra il corion e la mucosa uterina. In questo tipo di placenta, il trofoblasto penetra attraverso la mucosa, la lamina propria e l'endotelio vasale, prendendo contatto con i l sangue materno (schema 3a). Abbiamo due distinti casi, a seconda che la blastocisti aderisca poco precocemente e parzialmente all'endometrio (Coniglio) o si annidi rapidamente e totalmente nella mucosa (Uomo). Nel primo caso, l istogenesi della placenta si inizia in modo analogo a quello dei Carnivori. Nei secondo caso, la blastocisti, una volta completata la sua penetrazione, si sviluppa nel contesto stesso della mucosa da cui sospinge le diverse parti. Basandosi sull'ultrastruttura del trofoblasto Enders (1965) ha proposto di dividere le placente emocoriali in tre sottotipi secondo il numero degli strati 23
24 riconoscibili in questo rivestimento. Si possono cosi riconoscere placente: emotricoriali (Ratto, Topo, Criceto), emodicoriali (Coniglio) ed emomonocoriali (Uomo). Nei Roditori e Lagomorfi (Ratto, Cavia, Coniglio) la barriera placentare é costituita da un solo strato, rappresentato dall'endotelio dei capillari coriali; per tale motivo la placenta prende il nome di emo-endoteliale. È stato anche dimostrato che il sinciziotrofoblasto è derivato dal citotrofoblasto (ENDERS, 1965; BOYD e HAMILTON, 1966) e che, indipendentemente dal numero degli strati, vi é soltanto una membrana basale trofoblastica e che essa si trova sempre alla base dello strato più vicino del mesenchima fetale. Questa membrana basale può rappresentare una parte importante del meccanismo di barriera. Esistono due varietà di placente emocoriali: (a) nella varietà labirintica, più diffusa e di cui la Coniglia fornisce un esempio, il corion forma delle travate anastomizzate che delimitano degli spazi tubuliformi o canalicolari; questi comunicano formando una rete complessa percorsa dal sangue materno; (b) la varietà villosa, che si riscontra particolarmente nella donna di cui parleremo in seguito. Nella placenta diffusa il corion é coperto su tutta o quasi tutta la sua estensione da un tappeto di piccoli rilievi rossi o brunastri debolmente ingranati nelle cripte della mucosa uterina. Al microscopio si evidenzia che questi rilievi sono dei microcotiledoni costituiti da piccoli ciuffi di villi. Questi nei Suidi sono sostituiti da un sistema di pieghe irregolari suddivise in pieghe secondarie e perfino terziarie. La placenta diffusa offre l'utilizzo del massimo di superficie e cioè sembra compensare in certa misura la debolezza nell'ingranaggio delle parti materne con quelle fetali che conservano la loro integrità. Questo tipo di placente si riscontrano negli Equidi, nei Suidi, nei Camelidi, nei Tragulidi, nei Cetacei, nei Sirenidi e nei Lemuridi. Nei Suidi una piccola parte rimane liscia a ciascuna estremità dell'unità feto-placentare: la placenta viene detta in questo caso diffusa ma incompleta (Schema 4). La placenta zonata si sviluppa formando una fascia completa, larga e spessa attorno alla parte media del sacco coriale. Le estremità rimangono lisce e libere 24
25 formando una paraplacenta. I villi in seguito diventano assai complessi, costituendo delle travate che si ancorano profondamente nella mucosa uterina. L'erosione di quest'ultima crea degli ematomi, il cui riassorbimento forma, per degradazione dell emoglobina alla periferia dell'organo, ammassi pigmentati che ne modificano il colore. Come il tipo discoidale, ma in minor grado, questo tipo di placenta é deciduato. È comune in tutti i Carnivori, Pinnipedi e Fissipedi, e si trova più o meno chiaramente nei Proboscidati, nei Sirenidi e negli Iracoidei che conservano tuttavia una placenta diffusa (schema 4). La placenta discoidale realizza il massimo spessore e concentrazione dell'organo, che occupa soltanto una superficie quasi circolare (due in rare specie). Questa disposizione può comparire subito (Gliri) o succedere ad una placenta diffusa (Primati). La paraplacenta occupa sempre la maggior parte della superficie coriale. I villi coriali del trofoblasto hanno un grosso sviluppo con la formazione di un dispositivo labirintico o trabecolare a seconda della specie. Anche la mucosa uterina subisce forti modificazioni. Questo tipo di placenta é presente nei Gliri, negli Insettivori, nei Chirotteri e nei Primati; nelle scimmie Cinomorfe è bidiscoidale, cioè si forma un disco da ciascun lato del conceptus. Esistono degli aspetti intermedi per es. tra gli Elefanti dove abbiamo una placenta diffusa; tuttavia nella parte centrale del sacco coriale si sviluppa una zona più spessa che si approfonda maggiormente nell'endometrio; vi é quindi la coesistenza di una placenta diffusa e una zonata. In effetti nella maggior parte delle specie inizialmente abbiamo una placentazione diffusa ma le differenze si accentuano molto presto durante lo sviluppo proporzionalmente all'attività del trofoblasto e al grado di erosione dell'endometrio (Schema 4). 25
26 26
27 NUTRIZIONE ISTIOTROFICA ED EMOTROFICA La nutrizione placentare può essere effettuata con modalità istiotrofica od emotrofica. La nutrizione istiotrofica è caratteristica delle placente per opposizione e quella emotrofica delle placente vere, ma le due modalità possono coesistere. Nei primi stadi della gravidanza, cioè prima dell'impianto, la nutrizione é sempre istiotrofica anche quando, successivamente, verrà sostituita dall'emotrofica, come nel caso delle placente emocoriali. La nutrizione istiotrofica consiste nell'assorbimento da parte dell'epitelio dei villi coriali del prodotto di secrezione delle ghiandole della mucosa uterina che viene riversato nell'interstizio tra questa ed il corion. Tale prodotto viene detto istiotrofo o latte uterino ed é immesso nella circolazione ombelicale (BARONE,1983). La nutrizione emotrofica consiste nell'assunzione, sempre da parte dell'epitelio dei villi coriali, delle sostanze nutritizie direttamente dal sangue materno, con gli stessi meccanismi, quindi, che stanno alla base degli scambi nutrizionali tra sangue e tessuti. CIRCOLAZIONE SANGUIGNA PLACENTARE NEI MAMMIFERI La placenta é sede di una circolazione attivissima. Si ritiene, ad esempio, che la quantità di sangue che attraversa la placenta di una vacca quasi a termine di gravidanza sia di litri nelle 24 ore. In questa i suoi vasi evolvono contemporaneamente alla sua struttura e la loro disposizione quindi si modifica molto durante la gravidanza. Essi formano due reti distinte, una fetale e l'altra materna perfettamente indipendenti l una dall'altra; i due tipi di sangue dunque non si mescolano mai. A maturazione, la rete fetale é alimentata dalle ramificazioni delle arterie ombelicali che portano sangue non ossigenato. Molto numerose e frequenti nelle placente diffuse, le diramazioni di queste arterie formano, nelle placente cotiledonari, un arborizzazione di rami poco flessuosi di cui ogni ramificazione termina in un cotiledone. Negli altri tipi di placenta i rami sono molto flessuosi: nelle placente discoidali hanno una disposizione radiata a partire dall'inserzione del cordone prima che le loro ramificazioni si aggomitolino 27
28 in grandi territori ben delineati, ognuno dei quali corrisponde con un lobo dell'organo. In ogni caso, le ultime due diramazioni sono le arteriole afferenti delle unità placentari. Che si tratti di microcotiledoni o di villi di ogni ordine, queste arterie sono sprovviste di cellule muscolari lisce nel loro tragitto intraplacentare, mentre il tronco stesso dell'arteria ombelicale e le sue grosse divisioni possiedono una tonaca muscolare ben sviluppata. Hanno disposizione centrale nell'asse del cotiledone o dei villi. In ognuno di questi ultimi emettono delle collaterali che alimentano una rete capillare sub-trofoblastica molto ricca, che invia, spesso, delle anse o delle digitazioni fino al contatto delle cellule dell'epitelio. È drenata da venule che spesso raggiungono dei collettori assiali vicini alle arterie. Tuttavia, nella pecora, le vene efferenti sembra conservino una posizione periferica. Nelle placente labirintiche, le arteriole e le venule si situano entrambe al centro delle trabecole che percorrono in tutta la loro lunghezza alimentando reti capillari subtrofoblastiche. Le radici delle vene ombelicali sono sempre molto più voluminose e meno flessuose di quelle delle arterie corrispondenti. Esse portano all'embrione il sangue ossigenato e ricco di sostanze nutritive. La disposizione é molto più variabile dal lato materno. II sangue è portato da diramazioni delle arterie uterine che si sono differenziate inizialmente dalle reti del miometrio e dagli strati profondi dell endometrio. Queste diramazioni sono naturalmente molto più grosse ma meno numerose nelle placente discoidali rispetto alle placente diffuse o cotiledonari. Hanno una disposizione spiralata e molto flessuosa. I rami terminali formano, nelle placente adeciduate, arborizzazioni disposte come quelle dei cotiledoni ma allogate nei setti che separano le cripte dell'endometrio ed inversamente orientate, in modo da formare con esse due sistemi paralleli nei quali due tipi di sangue circolano in senso contrario. Nelle placente emocoriali, la rete materna decorre sulla superficie dell'organo e fornisce arteriole le cui ultime diramazioni si aprono beanti sia in spazi tubuliformi intertrabecolari (placente labirintiche), sia negli spazi perivillosi (placenta villosa, Uomo). In quest'ultimo caso la disposizione degli elementi anatomici imprime al sangue un movimento vorticoso attorno agli elementi coriali, prima di riportarlo verso le aperture che danno accesso alle venule uterine. 28
29 Nel cordone e nella placenta fetale sono state osservate cellule muscolari lisce che hanno la stessa origine di quelle che formano lo strato connettivale dei vasi ombelicali. FISIOLOGIA COMPARATA PLACENTARE Esistono delle differenze riguardo alle funzioni placentari dovute alle particolari conformazioni della placenta. Per esempio i protidi non riescono ad attraversare la placenta in forma complessa o specifica. La placenta possiede un'attività proteolitica propria; è in grado di scindere e sintetizzare sotto un'altra forma specifica differenti elementi materni. Quest'ultimo meccanismo potrebbe spiegare le difficoltà con la quale gli anticorpi materni superano la placenta nei Mammiferi domestici. Le placente epiteliocoriali, in particolare, costituiscono a questo riguardo una vera barriera; infatti, nei giovani ungulati il ruolo essenziale per stabilire l'immunità, è devoluto al colostro (primo latte). Un ruolo importante nel controllo di questi scambi tra madre e feto è stato attribuito alla struttura istologica della barriera placentare. II numero degli strati tissutali diminuisce andando dalle placente epiteliocoriali, che ne hanno sei (due epiteli, due strati di connettivo e due endoteli vascolari) alle placente emocoriali che ne hanno tre e persino due. Questi ultimi sono nettamente più permeabili a certe sostanze come gli anticorpi. Si deve notare inoltre che i neonati più sviluppati e più autonomi alla nascita sono proprio quelli degli Ungulati, la cui placenta é epitelio coriale, mentre le placente emocoriali appartengono, salvo rare eccezioni (Cavia), alle specie in cui la prole é più inetta. È stato dimostrato che la gonadotropina coriale e probabilmente un ormone coriale di crescita (la prolattina), sono presenti nel sinciziotrofoblasto dei villi e sono probabilmente sintetizzati qui (MIDGLEY e PIERCE, 1962; SCIARRA e Coll., 1963; YOSHIDA, 1964). Molte sono le caratteristiche in comune fra gli scambi gassosi placentari e quelli polmonari. La maggior differenza è costituita dal fatto che a livello placentare gli scambi avvengono fra liquido e liquido, a livello polmonare fra gas e liquido. Le 29
30 arterie ombelicali convogliano sangue desossigenato dal feto alla placenta mentre le vene ombelicali trasferiscono sangue ossigenato nella direzione opposta. Numerosi sono i fattori in grado di influenzare il trasporto placentare dell'ossigeno: le curve materna e fetale di dissociazione dell'emoglobina, lo spessore da attraversare, l'area in sui possono avvenire gli scambi, l'ossigeno utilizzato da tessuto placentare, l'affinità per l'ossigeno, i rapporti spaziali tra vasi materni e fetali e la velocità del flusso ematico materno e fetale (LONGO, 1972; COMLINE e SILVER, 1974). La curva di dissociazione dell'emoglobina fetale é spostata a sinistra rispetto a quella materna nella specie ovina, caprina, suina e bovina ma non in quella equina. Ciò significa che a qualsiasi tensione parziale di ossigeno, il sangue fetale é più ricco di ossigeno del sangue materno. Nella pecora, nella capra e nella vacca, il minimo gradiente di ossigeno fra il feto e la madre é di circa 20 mm Hg, mentre nella cavalla é di appena 2-4 mm Hg. Questa notevole affinità del sangue fetale per l'ossigeno può essere dovuta ad un particolare tipo di emoglobina (ruminanti), o alla presenza di un'elevata concentrazione d i 2-3 difosfoglicerato (DPG), che sposta la curva materna di dissociazione dell'ossigeno verso destra (scrofa). In quest'ultimo caso, l'emoglobina materna e fetale sono identiche, ma la minor differenza nella concentrazione in DPG fra eritrociti materni e fetali giustifica la ridotta differenza fra le due pressioni parziali di ossigeno. Sono stati suggeriti 4 possibili sistemi per gli scambi gassosi a livello placentare. L'anidride carbonica diffonde liberamente dal sangue fetale a quello materno. Le curve di cessione dell'anidride carbonica evidenziano una minor affinità del sangue fetale rispetto al sangue materno per la CO2, durante il passaggio dell ossigeno a livello placentare. Ciò favorisce la diffusione della CO2 dal feto alla madre. Importanti sono le funzioni di scambi placentari tra madre e feto di acqua e sali, carboidrati, proteine, lipidi, ormoni, vitamine, virus, germi e farmaci che presentano delle variazioni intraspecifiche molto complesse. 30
31 LA PLACENTA UMANA CARATTERISTICHE MACROSCOPICHE DELLA PLACENTA UMANA La placenta umana a termine di gravidanza appare in forma discoidale con un diametro massimo di 20 cm e uno spessore alla periferia di 0,50 cm e al centro 2-4 cm. Il suo peso é di circa grammi, cioè 1/6 del peso fetale. Presenta una faccia fetale liscia, in quanto ricoperta dall'amnios, e una faccia materna piuttosto irregolare per la presenza di 15, 30 lobulazioni separate da solchi. Questa conformazione biologica ha importanti implicazioni immunologiche, poiché l'insieme della placenta con il feto può essere visto come un "allotrapianto naturale "resistente al rigetto". I lobi prendono il nome di cotiledoni, mentre i solchi corrispondono alla base di altrettanti sepimenti o setti placentari che dalla placca basale si affondano nello spazio intervilloso senza quasi mai raggiungere la placca coriale e quindi non realizzando una completa compartimentazione dello spazio stesso. In una sezione sagittale della placenta "in situ", andando dalla parte rivolta verso l'utero alla parte rivolta verso il feto, si distinguono nella placenta a termine: la decidua basale, i villi, il corion, l'amnios (Schema 5; Fig. 6). 31

32 32
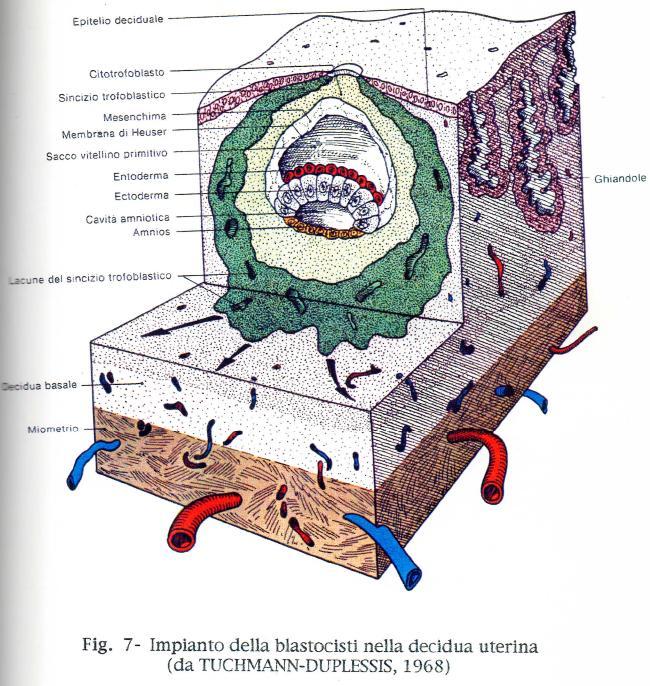
33 IMPIANTO DELLA PLACENTA UMANA La blastocisti umana si impianta generalmente nella parte superiore della parete posteriore del corpo dell'utero, in prossimità della linea sagittale mediana. Quando le cellule del trofoblasto vengono a contatto con l'epitelio uterino, cominciano a proliferare e, ben presto, si attaccano alla parete dell'utero. Con il progredire del differenziamento della placenta, diviene sempre più evidente che il trofoblasto forma il parenchima della parte fetale della placenta e costituisce l'elemento regolatore della barriera placentare. L'impianto della blastocisti umana é di tipo interstiziale; la blastocisti si approfonda infatti nel connettivo dell'endometrio e corrodendo le pareti vascolari viene circondata dal connettivo stesso. Questa sede interstiziale viene raggiunta verso l'11 giorno e l'epitelio dell'utero ricopre la sede dell'annidamento (Fig. 7). L'embrione é situato in una zona lievemente rilevata, brillante e traslucida del diametro di poco meno di un millimetro, circondata da una regione di colore rosso vivo che indica una modificazione dei vasi sanguigni nello stroma adiacente. L'osservazione microscopica delle sezioni di questo impianto di undici giorni dimostra che si é avuto un rapido accrescimento e differenziamento del trofoblasto. A quest'epoca sono chiaramente evidenti due componenti del trofoblasto: uno strato interno di citotrofoblasto primitivo, costituito da cellule distinte, ed uno spesso strato esterno di sinciziotrofoblasto primitivo. Il sincizio presenta ora degli spazi, detti lacune, che contengono sangue materno. Le lacune comunicano tra di loro e con i sinusoidi e le vene materne; queste connessioni vascolari consentono l'inizio della circolazione materna della placenta. La comparsa a quest'epoca nel circolo materno di un ormone prodotto dalla placenta, la gonadotropina corionica umana (HCG), costituisce la dimostrazione che si e stabilita una connessione con il sangue materno. Dati recenti ottenuti nel Macacus rhesus indicano che questo aumento iniziale delle gonadotropine corioniche in circolo mantiene attivo il corpo luteo, stimolando la secrezione da parte dell'ovaio del progesterone che mantiene la gravidanza fino a quando inizia la produzione di progesterone da parte della placenta, verso il ventitreesimo giorno di gravidanza (ATKINSON e Coll.,1975). 33
34 34
35 A questo stadio precoce dello sviluppo si stanno differenziando anche altri due annessi embrionali che non partecipano alla formazione della placenta: l'amnios e il sacco vitellino. La terza settimana di gravidanza (dal 14 al 21 giorno) é un periodo di intenso accrescimento e di differenziamento del trofoblasto, l'epoca in cui si stabiliscono importanti rapporti nella placenta (fig.8). Verso il 15 giorno la circolazione materna attraverso il sinciziotrofoblasto diviene pienamente funzionante, poiché le lacune divengono ampie e confluenti e si connettono con il sangue delle arterie spirali e con le vene dell'endometrio. Dalla superficie del corion verso l'esterno cominciano a formarsi cordoni di trofoblasto, detti villi coriali primari, per la rapida proliferazione del citotrofoblasto, che costituisce un fondamentale meccanismo cellulare di espansione della parte fetale della placenta (fig 8 a e b). Dopo il 15 giorno nella porzione prossimale, attaccata al corion dei villi compare del mesenchima, che si estende progressivamente verso l'estremità distale di accrescimento dei villi stessi. Man mano che al loro interno si forma un'asse mesenchimale, i villi primari si trasformano gradualmente in villi secondari. Ogni villo secondario é formato da un'asse centrale di mesenchima, circondato da uno strato continuo di citotrofoblasto, che é a sua volta rivestito da una lamina di sinciziotrofoblasto (Fig. 8 c e d). II sangue materno scorre attraverso gli ampi spazi intercomunicanti formatisi per la confluenza delle lacune del primitivo sinciziotrofoblasto, e che vengono ora indicati complessivamente con il termine di spazio intervilloso. In questo stadio precoce della gravidanza, la superficie del sinciziotrofoblasto é direttamente bagnata dal sangue materno circolante. Le estremità distali dei villi secondari sono ora colonne solide (colonne cellulari del citotrofoblasto) che si uniscono alla periferia formando il guscio trofoblastico che riveste tutto l'impianto e costituisce il limite più esterno dei tessuti dell'embrione. È principalmente costituito da citotrofoblasto, ma contiene anche alla periferia tratti irregolari di sinciziotrofoblasto, che in parte penetrano abbastanza profondamente nell endometrio e prendono contatto con i vasi sanguigni dell'utero corrodendoli. La formazione delle colonne di citotrofoblasto e l'organizzazione della lamina trofoblastica costituiscono un meccanismo di allungamento dei villi e di espansione circonferenziale della parte fetale della placenta. 35
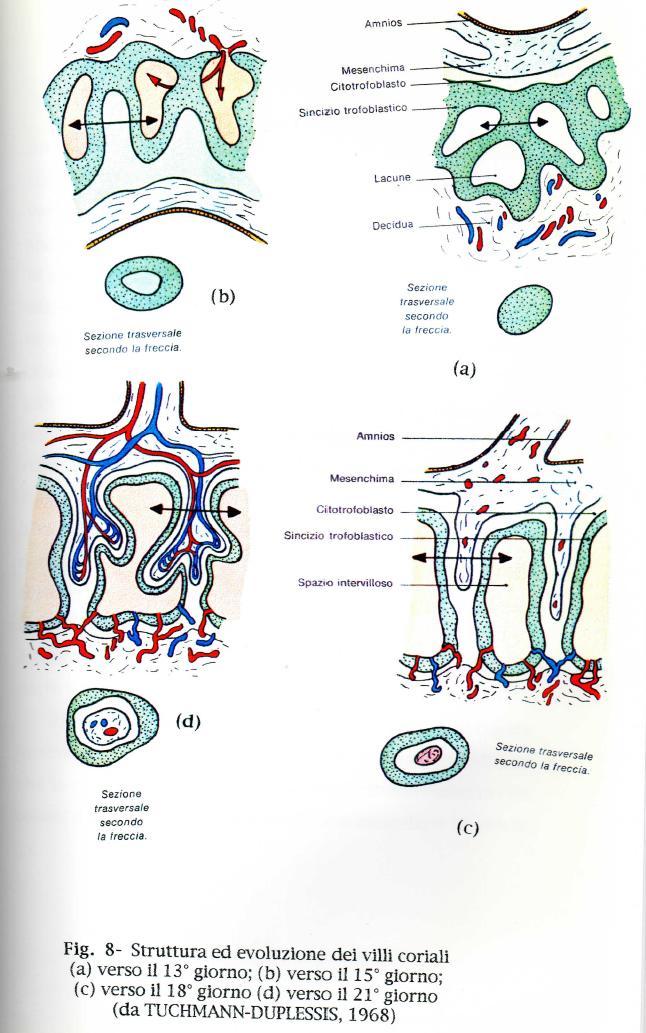
36 36
37 Recenti osservazioni al microscopio elettronico di questa iniziale penetrazione nello stroma dell'endometrio, indicano che le cellule del trofoblasto vengono a diretto contatto con le cellule deciduali materne. Questo intimo mescolarsi di cellule con differente genotipo é di grande interesse dal punto di vista immunologico ed é stato indicato per ora come complesso "deciduotrofoblastico" (TEKELIOGLU-UYSAL e Coll., 1975). In uno stadio successivo della gravidanza in aree localizzate di certi villi il citotrofoblasto prolifera formando isole cellulari del citotrofoblasto. Nell'asse mesenchimale dei villi compaiono i vasi sanguigni dell'embrione e si formano così i villi coriali terziari (Fig. 9). Compare anche l abbozzo del cordone ombelicale, mediante la formazione di un peduncolo addominale. Questo abbozzo endodermico e mesodermico unisce la porzione caudale del disco embrionale al corion, formando una placenta corioallantoidea. Il peduncolo allantoideo della maggior parte dei mammiferi contiene un diverticolo endodermico dell'intestino posteriore che, insieme al mesoderma dell'allantoide, forma il sacco allantoideo. Nell'uomo e nella scimmia l'allantoide resta rudimentale, pur rivestendo l'importante ruolo d'induttore della trasformazione del corion in corion frondoso, dando origine ai villi placentari, ai quali arrivano i vasi allantoidei. Successivamente il peduncolo addominale, all'interno del quale é situato il peduncolo allantoideo, si differenzia e si allunga, formando il cordone ombelicale che contiene i vasi ombelicali. Cosi i vasi sanguigni di recente formatisi nei villi coriali della placenta si connettono, mediante i vasi ombelicali, con il corpo dell'embrione; verso la fine della terza settimana, il sangue fetale comincia a circolare nei capillari dei villi. Ai villi coriali della placenta arriva ora il sangue fetale ed essi pescano nei sangue materno e gli scambi fisiologici vengono pertanto ad essere notevolmente aumentati. 37
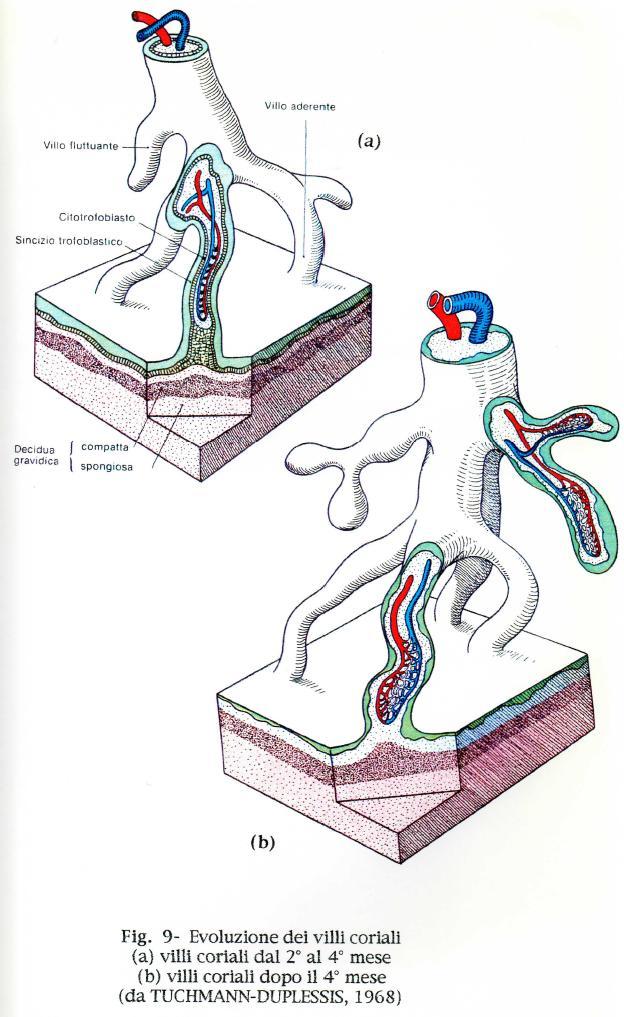
38 38
39 STRUTTURA E ULTRASTRUTTURA DEL VILLO CORIALE UMANO II trofoblasto rappresenta lo strato superficiale dei villi coriali, in diretto rapporto con il sangue materno. Esso risulta costituito dal sinciziotrofoblasto, dal citotrofoblasto o strato delle cellule d i Langhans e da una lamina basale continua. SINCIZIOTROFOBLASTO Lo stato di fusione di cellule trofoblastiche, nello strato sinciziale, avviene molto precocemente, a partire dalla 7 a all'8 a giornata di vita e prelude all'annidamento, grazie all'attitudine del sincizio di erodere la mucosa uterina e la parete dei vasi sanguigni materni. Lo strato sinciziale, subisce un ampliamento grazie alla confluenza in questo degli elementi del citotrofoblasto. Questo fenomeno si osserva per tutto l'arco della gravidanza, fino alle ultime fasi. Nei preparati di microscopia ottica, si apprezza l'affinità del citoplasma per i coloranti acidi, l'intensa basofilia nucleare e la irregolarità della superficie esposta al sangue materno. La superficie del sinciziotrofoblasto é estremamente irregolare per via delle numerose microvillosità che pescano nelle lacune sanguigne materne. I nuclei nel citoplasma sinciziale tendono a disporsi in gruppi più o meno numerosi, e la cromatina al loro interno presenta delle zone di addensamento. Nelle sezioni tangenziali dei villi si osserva che nel citoplasma sinciziale compaiono dei vacuoli otticamente vuoti, interpretati come cisterne dilatate ed inclusioni lipidiche, estratte dai solventi organici utilizzati nell'allestimento dei preparati. II citoplasma sinciziale si distingue da quello delle cellule di Langhans per la sua differente affinità tintoriale. Lo spessore é variabile e aumenta in prossimità dei nuclei. I villi liberi non hanno un asse longitudinale rettilineo ma si incurvano a formare delle anse più o meno strette. I villi inoltre sono ramificati e la loro superficie é di solito irregolare, tale da simulare in sezione obliqua fenomeni di accrescimento realmente presenti e numerosi nelle prime fasi della gestazione (KUSTERMANN, 1981; KAUFMANN e Coll., 1987). In sezioni ultrafini al microscopio elettronico, si apprezza la ricchezza 39
40 in organuli e la presenza di microvillosità apicali e basali del plasmalemma che aumenta la superficie di scambio. Le microvillosità si osservano oltre che sulla parte terminale del villo, anche su tutto il tronco (HERBST e MULTIER, 1970). La distribuzione di queste microvillosità non é omogenea, infatti esistono zone in cui sono più numerose rispetto ad altre. Questo rispecchia una diversa attitudine funzionale del sinciziotrofoblasto per cui, due autori (BURGOS e RODRIGUEZ, 1966) distinguono nel sincizio placentare una zona e una zona. Una orientata verso il trasporto e l'altra verso l'attività di sintesi. L'ipotesi e stata ripresa da altri ricercatori (VACEK, 1974; FOX, 1978), e si é arrivati alla certezza che esistano zone differenziate nel sincizio. L'importanza ai fini di scambi metabolici delle microvillosità, é suffragata dalla presenza di numerose caveole che preludono alla costituzione di vescicole pinocitotiche osservate al microscopio elettronico. II microvillo presenta un asse microfilamentoso centrale. Indagini immunoistochimiche hanno individuato la presenza di actina nell'asse microfilamentoso delle microvillosità (TEDDE, TEDDE-PIRAS, 1985); è stato inoltre osservato che l asse microfilamentoso si ingrana con un ben sviluppato "terminal web" sito sulla superficie del sinciziotrofoblasto (KING, 1983). All esterno del villo, i microvilli possono assumere un aspetto morfologico irregolare o disporsi a formare dei ciuffi, i cui filamenti presentano lunghezze omogenee e si impiantano su un peduncolo comune. Alla superficie del sincizio é possibile osservare oltre alle caveole, che testimoniano fenomeni di micropinocitosi, formazioni globose, peduncolate, da identificare come vescicole secretorie. La presenza di queste vescicole in superfici meno ricche di microvillosità, fa intuire una diversa qualità funzionale. Le differenze morfologiche nelle varie fasi di gravidanza del sinciziotrofoblasto, riflettono diverse attività enzimatiche e recettoriali molto complesse, fenomeni di trasporto e della recezione di segnali trofici, che influenzano le capacità di sintesi dell'elemento sinciziale (HULSTAERT e Coll., 1973; MARTIN e Coll., 1974; JENKINSON e Coll., 1976; NELSON e Coll., 1976; JOHNSON e FAULK, 1978; KING, 1981, 1982; WASSERMAN e Coll., 1983; PARMLEY e Coll., 1984; PARMLEY e Coll., 1985). Anche il comportamento delle attività fosfatasiche alcaline, varia nel corso della gestazione. 40
41 È infatti presente nel pieno spessore nella prima metà della gestazione e si riduce esclusivamente alla parte superficiale, apicale e basale del plasmalemma al termine (TEDDE, 1987). Anche la morfologia dei nuclei del trofoblasto varia durante l'arco della gravidanza. Questi si raggruppano tendenzialmente in aree relativamente ristrette e presentano, in sezione, forma ovalare. Nel primo trimestre di gravidanza, hanno contorni regolari e la cromatina risulta riunita in zolle cromofile congiunte alla periferia del carioplasma. Con l'avanzare dell'età gestazionale, i contorni nucleari divengono più irregolari; le zolle di cromatina divengono più voluminose e aumentano le dimensioni dei singoli nuclei. A metà del periodo gestazionale si osservano raggruppamenti di nuclei circondati da un sottile alone di citoplasma che sembrano tendere al distacco. Tale fenomeno è noto da tempo ed è stato interpretato come evento di segregazione di nuclei non più atti a sovraintendere ai fenomeni di sintesi cellulare e, pertanto, destinati a venir espulsi dal citoplasma sinciziale (JONES e FOX, 1977; CANTLE e Coll., 1987). Riguardo all'aumento delle dimensioni nucleari nel corso della gravidanza, metodiche stereologiche applicate alla microscopia elettronica hanno dimostrato l'aumento della frazione di volume dei nuclei del sincizio espressa in relazione al volume cellulare. Questa frazione tende a passare da un valore di 16,12 nel primo trimestre ad uno di 22,80 nel secondo e di 24,42 nel terzo (TEDDE e KUJAWA, 1978), (tab. 1). Analisi al microscopio ottico, hanno rivelato un passaggio delle dimensioni superficiali dei microvilli da valori di 70 2 nel primo trimestre di gravidanza, a valori di nei terzo trimestre (TEDDE e DELRIO, 1981; DELRIO e TEDDE, 1981), (tab. 2). Nel sincizio placentare non avvengono divisioni cariocinetiche, tuttavia i risultati conseguiti dalle indagini riportate, suggeriscono 1'ipotesi che nei sinciziotrofoblasto della placenta umana si verifichino fenomeni di endomitosi con formazioni di nuclei poliploidi. Questo particolare comportamento dei nuclei è stato messo in relazione con uno stato di iperattività nucleare. Questo dimostra la presenza di numerosi inclusi nucleari, sia di tipo granulo-filamentoso, che vescicolare, da tempo messi in evidenza nel sinciziotrofoblasto (VACEK, 1970; TEDDE, 1971). II citoplasma sinciziale presenta una discreta opacità agli elettroni con una grandissima quantità di organuli citoplasmatici e di inclusioni presenti. 41
42 42
43 Possono essere messi in evidenza mitocondri a creste tubulo-vescicolari, numerose cisterne del reticolo endoplasmatico agranulare e granulare, apparati di Golgi, lisosomi, vescicole secretorie a contenuto denso e numerose inclusioni lipidiche. Questi particolari organuli citoplasmatici indicano che a livello sinciziale avvengono importanti fenomeni di sintesi steroidea. La sintesi é orientata verso la produzione di estrogeni e progesterone; produzione che può essere in parte inibita, anche in vitro, con inibitori dell'attività aromatasica, quali nicotina, cotienina e anabesina (BARBIERI e Coll., 1986). Inoltre, la presenza del reticolo granulare, apparato del Golgi e vescicole secretorie indicano un coinvolgimento del sinciziotrofoblasto anche nella produzione di sostanze proteiche e glicoproteiche. Sono state compiute ricerche con metodiche stereologiche applicate alla microscopia elettronica al fine di valutare in termini quantitativi la presenza dei singoli organuli citoplasmatici nei tre trimestri di gravidanza e nelle placente a termine (TEDDE e GENEROSO, 1976; TEDDE e KUJAWA, 1978). Dai risultati appare come il reticolo endoplasmatico granulare 43
44 tenda ad aumentare con il procedere della gravidanza; ciò è in accordo con l'aumento di produzione di determinate sostanze di natura proteica, quali enzimi, ormoni, principi attivi. La frazione di volume di alcuni organuli come mitocondri e lisosomi e delle inclusioni citoplasmatiche (gocce lipidiche e vescicole secretorie), si mantengono immutate fino al termine. L'apparente riduzione della frazione di volume relativa al reticolo endoplasmatico agranulare non é risultata statisticamente significativa. Nella placenta a termine, i valori della frazione di volume decadono significativamente, tranne che per i lisosomi. La riduzione improvvisa degli organuli citoplasmatici, al termine della gravidanza, normalmente indica fenomeni involutivi massivi, che coinvolgono in toto il sinciziotrofoblasto, riducendone bruscamente e irreversibilmente la capacita di sintesi. Le caratteristiche morfologiche riscontrate al microscopio elettronico, si accordano con le ipotesi di una partecipazione diretta del sinciziotrofoblasto alla produzione di ormoni e di sostanze attive placentari. Con le utilizzazioni di varie metodiche in vivo e in vitro e con l'utilizzazione di tecniche imunocitochimiche e immunochimiche è stata dimostrata l induzione da parte del sinciziotrofoblasto, di numerosi enzimi: di HCS ("somatomammotropina coriale umana"); quest'ormone viene detto anche HPL (lattogeno placentare umano), la cui molecola è sovrapponibile a quella dell'ormone GH (adenoipofisario), HCT ("tirostimolina coriale umana"), SP1, SP2, SP3, PP5, PP12, relaxina, renina, ACE, ACTH, MSH, catepsina D, vasopressina, LH-RH, prostacicline, acetilcolina, nonchè gli ormoni steroidei quali il progesterone, l'estrone, l'estradiolo e l'estriolo (JOSIMOVICH e MACLAREN, 1962; KAPLAN e GRUMBACH, 1964; HENNEN, 1965; SITTERI e MACDONALD, 1966; DALION e BOTON, 1967; DICZFALUSY, 1967; BRODY, 1969; HELLIG e Coll., 1970; BOHM, 1971; DUJARDIN e Coll., 1977; LIOTTA, 1977; SILER-KHODR e KHODR, 1978; GASPARD, 1980; TAKEMORI e Coll., 1981;CONTRACTOR e Coll., 1982; TAN e ROUSSEAU, 1982; PATTILO e Coll., 1983; RAKOCZI e Coll., 1983; LEVENTER e ROWELL., 1984; TAIRA e Coll., 1985; AL-TIMINI e FOX., 1986; MULDER e Coll., 1986; FUJIMOTO e Coll., 1986; ROSEN, 1986; VETTENRANTA e Coll., 1986; GOSSRAN e Coll., 1987). 44
45 Esistono prove istoimmunologiche che testimoniano la sintesi della frazione di HCG a livello sinciziale (GASPARD, 1980). Controllando la secrezione in vitro da parte del trofoblasto di HPL, HCG e SP1, WINIKOFF e BRAUNSTEIN (1985) ritennero di non escludere che il tessuto placentare a termine possa contenere un inibitore della liberazione di HCG. Attraverso lo studio compiuto su villi dall'ottava alla dodicesima settimana e al termine, si é dimostrato che il sinciziotrofoblasto della placenta umana produce renina. I valori assoluti dell'attività reninica, sono tre volte più elevati nelle fasi precoci rispetto al termine di gravidanza e non vi è relazione tra attività reninica corionica e attività reninica plasmatica materna, il che fa escludere l'origine materna della renina liberata dai villi placentari (TEDDE e Coll., 1986,1987; BAGELLA e Coll., 1987). La particolare estensione del sinciziotrofoblasto e la sua particolare posizione superficiale, in diretto contatto con il sangue materno, sono fattori che stanno ad indicare il ruolo che il sinciziotrofoblasto gioca nei meccanismi di trasporto transplacentare. Tali meccanismi sono rappresentati dalla diffusione semplice, da una diffusione facilitata, dal trasporto attraverso sistemi chiusi di membrane e dal trasporto attivo mediante carriers specifici (COX e CHALMERS, 1953; YAMAGUCHI e Coll., 1960, 1964; NAKANO, 1963; GITLIN e Coll., 1964, 1965; DANCIS, 1965; MACDONALD e Coll., 1965; KOHLERe FARR, 1966; HAY e Coll., 1971; GITLIN, 1973; BOYD e HAMILTON, 1975; JOSIMOVICH, 1975; FOX, 1978; MIRKIN, 1978: CARPENTER, 1980). Studi autoradiografici eseguiti su villi coriali incubati per tempi differenti in presenza di sostanze marcate (C-Urea,C-Alanina,C-Destrano,C-Inulina), hanno rivelato un meccanismo di trasporto attivo per l'urea e l alanina; l'inulina non é in grado di attraversare lo strato trofoblastico e il destrano prende la via intercellulare (FENU e Coll., 1988). Sulle basi di osservazioni condotte al microscopio elettronico é stato possibile evidenziare che il sinciziotrofoblasto é costituito da numerose entità polinucleate, denominate "unita sinciziali" (TEDDE, 1982). Esse sono separate da fessure profonde dirette dall'apice verso la base, al cui livello si trovano desmosomi e giunzioni serrate (METZ e Coll., 1979; TEDDE, 1982). 45
46 II sinciziotrofoblasto presenta nel suo plasmalemma apicale e basale, una notevole quantità di microvillosità, di caveolae, di introflessioni della membrana, che rappresentano una tappa di formazione delle vescicole pinocitotiche; queste ultime, numerose, non si identificano sempre con chiarezza a causa della presenza delle cisterne del reticolo endoplasmatico agranulare, con le quali possono confondersi. Il sistema di trasporto attivo per pinocitosi è sicuramente presente nello strato sinciziale come a livello degli altri strati del villo coriale. Nei citoplasma sinciziale sono compresi corpi multivescicolari, da intendersi quali sistemi segregati di trasporto percellulare. CITOTROFOBLASTO Lo strato delle cellule di Langhans o del citotrofoblasto è facilmente identificabile, anche nelle sezioni istologiche comuni. Infatti la sua posizione risulta più profonda rispetto a quella sinciziale e risulta rivolta verso l'asse mesenchimale del villo. Le cellule di Langhans presentano un'afflnità tintoriale differente rispetto al sinciziotrofoblasto; infatti risultano meno basofile rispetto a quest'ultimo. Attraverso lo studio compiuto su sezioni sottili, si é potuto osservare che per tutto il primo trimestre di gravidanza il citotrofoblasto é costituito da uno strato continuo di cellule, che media il rapporto tra sinciziotrofoblasto ed asse mesenchimale del villo coriale. Le cellule del citotrofoblasto permangono fino al termine. Il loro numero si riduce, e per il loro diradarsi e disperdersi nel contesto dello strato trofoblastico in espansione, é raro osservare la loro presenza nella gravidanza avanzata. Nel primo trimestre, le cellule del citotrofoblasto presentano una forma globosa e possiedono u n nucleo voluminoso a contorni regolari. La cromatina risulta dispersa e le cellule risaltano per una cromofilia citoplasmatica differente rispetto a quella sinciziale. A partire dalla 14 a o 15 a settimana, le cellule di Langhans iniziano a distanziarsi e, negli spazi lasciati liberi, si approfondano propaggini citoplasmatiche dallo strato sinciziale. In questo modo si fanno più evidenti le unita cellulari, le quali tendono ulteriormente a diradarsi, fino ad apparire diminuite progressivamente. Questo é 46
47 un fenomeno soltanto apparente determinato da un progressivo e sensibile aumento della superficie dei villi coriali non accompagnata da particolare proliferazione degli elementi del citotrofoblasto. Negli ultimi mesi di gravidanza e nelle placente a termine, le cellule di Langhans presentano una forma allungata, con I'asse maggiore parallelo alla superficie del villo, mentre non si modifica l'affinità tintoriale del loro citoplasma che rimane pittosto scarsa (TEDDE, 1970). È possibile osservare la divisione cariocinetica delle cellule di Langhans durante tutto l'arco della gravidanza. Alcune ricerche hanno dimostrato che l'attività proliferativa è piuttosto intensa fino alla 20 a settimana; essa tende a decrescere progressivamente pur non scomparendo che nelle fasi più tardive della gestazione (TEDDE e TEDDE PIRAS, 1978b), (diagramma 1). In osservazioni compiute su sezioni semifini al microscopio ottico, si osservano particolari elementi di Langhans, il cui citoplasma presenta un'affinità tintoriale di intensità intermedia tra quella caratteristica dello strato citotrofoblasto e quella del sincizio sovrastante. Su indagini compiute al microscopio elettronico è possibile affermare che si tratta di elementi del citotrofoblasto in procinto di confluire nello strato sinciziale. Al microscopio elettronico si osserva un notevole incremento di organuli citoplasmatici delle cellule di Langhans. 47
48 I mitocondri assumono creste tubulo-vescicolari e inoltre compaiono cisterne nel reticolo endoplasmatico liscio e lisosomi. Questi sono elementi destinati a confluire nei sincizio e che, pertanto, possiamo considerare come elementi "intermedi" (TEDDE, 1970). Le cellule di Langhans, situate profondamente al sincizio, sono a questo connesse da numerosi apparati giunzionali di tipo desmosomiale. Le giunzioni desmosomiali sono presenti anche tra cellule di Langhans contigue (TEDDE e TEDDE-PIRAS, 1978a; METZ e WEIHE, 1980). In studi condotti con la tecnica del freeze-fracture é stato possibile osservare occasionalmente gap-juncton tra le cellule di Langhans contigue e tra queste ed il sincizio (De VIRGILIIS e Coll., 1982). Il nucleo di queste cellule è voluminoso, ha contorni regolari e ha cromatina dispersa. II citoplasma differentemente da quello sinciziale, presenta scarsa densità elettronica. Gli organuli citoplasmatici sono rappresentati da mitocondri allungati, a creste lineari, da sparse cisterne del reticolo endoplasmatico granulare e da sviluppati complessi di Golgi. Sono 48
49 presenti in gran quantità anche ribosomi liberi, per lo più riuniti in polisomi, ed è presente una gran quantità di glicogeno. Si osservano molte vescicole lisce a contenuto denso in prossimità dell'apparato di Golgi da interpretare come vescicole secretorie. Nelle zone periferiche del citoplasma vi sono delle vescicole di piccole dimensioni rivestite da una membrana su cui aderisce un caratteristico materiale filamentoso, le cosiddette "coated vesicles". II ruolo di queste vescicole non e stato chiarito completamente. Secondo delle ipotesi, attraverso tali vescicole la cellula può inglobare delle proteine dal microambiente circostante, secondo altre esprimerebbero un meccanismo di recupero di membrana del plasmalemma, visto l'enorme aumento di quest'ultimo a causa degli intensi fenomeni di esocitosi. Attraverso indagini morfometriche compiute lungo tutto I'arco della gravidanza, si è osservato che i mitocondri, le vescicole secretorie e le "coated vesicles" aumentano in quantità, con il procedere dell'età gestazionale, a livello delle cellule di Langhans (TEDDE e KUJAWA, 1983), (tab. 3 e 4). Da questo risultato si ottiene una conferma circa l'attività secretoria svolta dal citotrofoblasto. Le caratteristiche citoplasmatiche delle cellule di Langhans, inducono a pensare a fenomeni di sintesi, orientate alla produzione di proteine plasmatiche A associate alla gravidanza (DABASHI e Coll., 1984) e di HCG. II coinvolgimento da parte delle cellule di Langhans nella sintesi delle sub-unità della gonadotropina corionica, é stato messo in evidenza attraverso indagini di isto-immunologia (CHUNG e Coll., 1969; COTTE e Coll., 1980; GASPARD, 1980; NELSON e Coll., 1986). Per mezzo di studi isto-immunologici é stata dimostrata la sintesi di Inibina e di CRF (ormone di liberazione per l'acth) da parte delle cellule di Langhans (PETRAGLIA e Coll., 1987a, 1987b). Questa attività secretoria, suggerisce l'ipotesi che il citotrofoblasto sia costituito da due tipi cellulari distinti, alcune secernenti ed altre di grande capacità proliferativa, destinate a confluire nei sincizio. Non é tuttavia da escludere la possibilità che uno stesso tipo cellulare possieda entrambe le espressioni funzionali. Intorno alle cellule di Langhans si sviluppa un sistema di canalicoli intercellulari ampiamente comunicanti, nei quali pescano numerose microvillosità sia del sincizio che delle stesse cellule di Langhans. Quando le suddette cellule 49
50 sostituiscono uno strato continuo (primo trimestre di gravidanza), il sistema canalicolare intercellulare si spinge anche tra gli elementi contigui, per aprirsi sul versante profondo della lamina basale del trofoblasto. La presenza di microvillosità e di caveolae nei plasmalemmi prospicienti il sistema canalicolare intercellulare, testimoniano l'esistenza di fenomeni intensi di scambio tra le cellule e l'ambiente extracellulare; ciò permette di considerare gli spazi intercellulari come utilizzati per il passaggio di sostanze dal sincizio al citotrofoblasto e viceversa (TEDDE e TEDDE PIRAS, 1978; TEDDE e Coll.,1993). 50
51 51
52 LAMINA BASALE DEL TROFOBLASTO Tra epitelio e tessuto connettivo é sempre interposta una lamina extracellulare chiamata membrana basale, atta a regolare gli scambi nutritizi tra epitelio e tessuto connettivo. La membrana basale difficilmente si vede nei preparati istologici ma può essere evidenziata con reazioni acido periodico-schiff (PaS) e metodi di impregnazione argentica. La lamina basale continua, situata in profondità rispetto il sincizio ed alle cellule di Langhans, presenta variabili caratteristiche costitutive, spessore e rapporti nei corso della gravidanza. Nei primo trimestre risulta avere un rapporto esclusivamente con le cellule del citotrofoblasto che costituisce uno strato continuo. Quando le cellule di Langhans si diradano, con l'avanzare della gravidanza, il sincizio entra in rapporto con la lamina basale in maniera sempre più estesa. Il plasmalemma del sinciziotrofoblasto emette numerose e sottili evaginazioni che delimitano uno spazio labirintico a ridosso della lamina basale, lo "spazio subsinciziale" (TEDDE, 1970). Nei primi mesi di gravidanza la lamina basale è costituita da materiale a scarsa densità elettronica. Nei progredire della gravidanza si osserva la comparsa di una trama microfilamentosa che va rinforzandosi. La lamina basale contiene laminina e collagene IV (AMENTA e Coll., 1986). La lamina basale passa da valori dello spessore di 0,15-0,18 all'ottava e nona settimana, bruscamente a valori di 0,40-0,48 alla diciassettesima-diciottesima settimana, rimanendo invariato fino al termine (TEDDE e DELRIO, 1980), (grafico 2). II brusco aumento di spessore si osserva in relazione a caratteristiche funzionali fino a quel momento assenti. Non è escluso che siano fattori endocrinologici a provocare modificazioni morfologiche a carico della barriera placentare. L aumento della trama microfibrillare può essere messo in relazione con l'aumento del trasporto trans-placentare. È possibile osservare modificazioni profonde in lamine basali presenti in organi a secrezione endocrina, laddove le lamine basali sono sottoposte a continue modificazioni in ragione dell'atteggiamento funzionale dell'epitelio ghiandolare (FAZZARI, 1929, 1951; ALLARA, 1940, 1950, 1955; BALBONI, 1962, 1973). Le 52
53 caratteristiche morfologiche della lamina basale del trofoblasto possono, pertanto, riflettere lo stato d'essere funzionale del sinciziotrofoblasto e delle cellule di Langhans. 53
54 ASSE MESENCHIMALE DEL VILLO L'asse mesenchimale del villo rappresenta la porzione centrale del villo coriale. È costituito da tessuto connettivo giovane, formato da cellule a contorni molto irregolari ed abbondante sostanza intercellulare, molto fluida e scarsamente formata. Anche nell'asse mesenchimale, le caratteristiche morfologiche si modificano nei corso della gestazione. CELLULE DELL'ASSE MESENCHIMALE La popolazione cellulare dell'asse mesenchimale del villo coriale è costituita da almeno quattro tipi cellulari: le cosiddette cellule "reticolari", i fibroblasti, i fibrociti e i macrofagi. Le cellule "reticolari" devono il loro nome al fatto che dal loro corpo cellulare piuttosto piccolo, si dipartono delle evaginazioni laminari che raggiungono altre evaginazioni laminari di altre cellule "reticolari", costituendo cosi una rete tridimensionale particolarmente evidente nel primo trimestre di gravidanza. II corpo cellulare di queste ha forma fusiforme che in sezione trasversale gli fa assumere un caratteristico aspetto aracnoide. Le propaggini laminari vanno a costituire quasi dei "canali" interstiziali. Questi, nei preparati istologici, dimostrano un contenuto fluido, cromofobo e trasparente agli elettroni. È stato possibile rilevare al microscopio elettronico che risultano riccamente anastomizzati tra loro e che le loro caratteristiche tendono a modificarsi con il procedere della gestazione (ZACCHEO e CASTELLUCCI, 1981). I canali interstiziali sono considerati come sede di movimenti di liquido tissutale e come deputati al trasferimento dei soluti in senso materno fetale. Ciò è verosimile, considerando il rapporto che solitamente i canali contraggono con il sistema vascolare del villo nel primo trimestre di gravidanza (KAUFMANN e Coll., 1977; CASTELLUCCCI e Coll, 1984). Dal secondo trimestre in poi, le cellule "reticolari" sembrano retrarre i loro prolungamenti perdendo contatto con gli elementi vicini; di conseguenza, i canali interstiziali scompaiono e gli spazi vuoti lasciati da questi ultimi, vengono occupati da sostanza intercellulare amorfa e da fascetti di fibre collagene (KAUFMANN e Coll., 1977). 54
55 I fibroblasti rappresentano un tipo cellulare presente durante tutto l'arco della gestazione. Sono particolarmente numerosi fino al sesto, settimo mese e tendono a rendersi più rari fino al termine. II nucleo dei fibroblasti è voluminoso, di forma ovalare, a cromatina dispersa e sovente nucleolato. Il citoplasma è abbondante, ha spiccata affinità per i coloranti basici e presenta contorni irregolari a causa della presenza di profonde evaginazioni della membrana cellulare. In sezioni semifini colorate con blu di touluidina, il citoplasma appare tipicamente granuloso. Nel citoplasma osservato al microscopio elettronico si mette in evidenza una quantità di RNA, sia sotto forma di ribosomi liberi, che aggregati a cisterne appiattite del reticolo endoplasmatico granulare. Le cisterne sono in contiguità con mitocondri, di forma allungata, con creste lamellari e matrice densa. II plasmalemma sovente si invagina per costituire profonde tasche, all'interno delle quali si trovano formazioni microfilamentose da interpretare come filamenti di tropocollagene secreto dalle cellule e destinato ad aggregarsi per costituire le caratteristiche fibrille collageniche a struttura periodica. I fenomeni di collagenizzazione dell'asse mesenchimale risultano più evidenti nelle placente del secondo trimestre di gravidanza. I fibrociti dell'asse stromale del villo sono rari nei corso della prima metà della gravidanza, mentre tendono a farsi più numerosi via via che ci si avvicina al termine. Presentano forma fusata, il nucleo allungato secondo l asse maggiore della cellula, la cromatina addensata in zolle grossolane. II citoplasma è scarso ed i contorni cellulari sono irregolari per via delle lunghe evaginazioni spesso ramificate. Al microscopio elettronico si osservano scarsi granuli citoplasmatici. I fibrociti sono segno di una avvenuta maturazione del connettivo del villo e indicano un esaurimento funzionale dei fibroblasti che hanno completato l attività di sintesi e secrezione del tropocollagene. Nelle placente a termine i fibrociti, tendono a disporsi concentricamente ai vasi sanguigni fetali costituenti strati cellulari appiattiti alternati a strati acellulari costituiti da sostanza intercellulare amorfa a media densità elettronica (TEDDE, 1970). I macrofagi placentari, detti anche cellule d i Hofbauer, sono elementi cellulari voluminosi, presenti nell'asse del villo durante tutto l'arco della gravidanza. II citoplasma presenta una caratteristica eosinofilia. II loro nucleo presenta 55
56 dentellature del profilo e la cromatina pare condensata alla periferia del citoplasma. Nel citoplasma sono presenti vacuoli di varia dimensione, localizzati in posizione periferica. Queste cellule presentano una buona attività cariocinetica (CASTELLUCCI, 1987). Al microscopio elettronico si osservano costantemente evaginazioni citoplasmatiche, numerosi vacuoli, lisosomi primari e secondari, in posizione perinucleare, cisterne del reticolo endoplasmatico e piccoli mitocondri a matrice densa (TEDDE, 1970; CASTELLUCCI e Coll., 1980; DEMIR e HERBENGI, 1984; MARTINOLI e Coll., 1984). Durante la prima meta della gestazione, i macrofagi tendono a disporsi a stretta continuità con la lamina basale del trofoblasto che in posizione perivascolare. Inoltre risultano costanti rapporti citoplasmatici delle cellule di Hofbauer con i periciti dei capillari fetali. Le cellule d i Hofbauer giocano un ruolo importante nel bloccare le sostanze a prevalente significato antigenico, che provenendo dal sangue materno, riescono ad attraversare gli strati epiteliali superficiali del villo. È stato possibile, attraverso metodiche specifiche, evidenziare recettori specifici per alcune delle immunoglobuline (MOSKALEWSKI e Coll., 1975). I macrofagi placentari inoltre sembrano attivare la proliferazione dei linfociti T. Sono inoltre positivi agli anticorpi monoclonali anti-macrofagi OKM1, Leu M2 e 4F2. Secernono, inoltre, lisozima(zaccheo e Coll., 1984). I rapporti stretti con la lamina basale e con le formazioni perivascolari delle cellule di Hofbauer sono strettamente inerenti alle specifiche funzioni di difesa dei tessuti fetali in un periodo in cui il corpo fetale non ha ancora raggiunto il completamento dei processi differenziativi a carico dei sistemi tissutali di difesa specifica. Durante la seconda metà del periodo di gravidanza, infatti, sembrano essere meno evidenti questi rapporti. SOSTANZA INTERCELLULARE DELL'ASSE MESENCHIMALE La sostanza intercellulare del villo coriale modifica le sue caratteristiche durante l'arco della gravidanza. Essa passa da una notevole fluidità del primo trimestre ad un processo di collagenizzazione per opera dell'attività secretoria dei fibroblasti, nel secondo e terzo trimestre. Già dalla settima settimana si individuano esili fibre collagene che tendono a collocarsi in aderenza con lo strato profondo della 56
57 lamina basale del trofoblasto. I fascetti di fibre collagene divengono via via più numerosi e si dispongono in maniera casuale negli spazi intercellulari dell'asse mesenchimale. Verso il termine della gravidanza, i fasci di fibrille assumono caratteristiche di feltro di collagene immerso in una matrice amorfa a media densità elettronica di natura polisaccaridica, vista la sua spiccata PaS-positività. La collagenizzazione dell'asse del villo coriale deve essere interpretata come maturazione del mesenchima, che si accompagna ad una più spiccata selettività negli scambi metabolici materno fetali. Solamente nella placenta a termine o in zone ristrette della placenta al terzo trimestre, il connettivo del villo presenta un quadro morfologico che configura uno stato di sclerosi connettivale, segno inequivocabile di esaurimento delle capacita funzionali dei tessuti placentari. CAPILLARI FETALI I capillari sanguigni fetali variano notevolmente per numero e dimensione nelle sezioni trasversali del villo terminale. Solitamente la posizione preferenziale risulta eccentrica e a ridosso della lamina basale del trofoblasto, tanto che GETZOWA e SADOWSKI (1950), coniarono il termine "membrana vascolo sinciziale" riferito al complesso sincizio-lamina basale endotelio. Nel primo trimestre di gravidanza, i capillari appaiono sprovvisti di lamina basale, la quale inizia a comparire attorno alla l4 a -l5 a settimana. Dapprima essa risulta discontinua e si ispessisce progressivamente fino al termine. In alcuni punti la lamina basale del trofoblasto si fonde completamente con quella dei capillari. In rapporto con le cellule endoteliali dei capillari sanguigni sono presenti elementi allungati denominati periciti. In principio ad essi era stata assegnata una funzione di regolazione del letto capillare, ma le caratteristiche morfologiche e ultrastrutturali ne escludono questa funzione. PERICITI DEI CAPILLARI FETALI I periciti sono cellule appiattite addossate all'endotelio dei capillari fetali. In sezioni semifini osservate al microscopio ottico si osserva, nelle prime fasi della 57
58 gravidanza, una caratteristica basofilia citoplasmatica. I periciti sono costantemente presenti nei primi mesi di gravidanza, mentre tendono a scomparire o a modificare notevolmente le loro caratteristiche ultrastrutturali con il procedere della gestazione (LISTER, 1963; TEDDE, 1970, 1973). Essi non formano uno strato continuo ed appaiono aderenti al versante esterno delle cellule endoteliali. Tuttavia risultano separate da esse da uno spazio variabile, che solitamente é dell'ordine di qualche centinaio di Å. I periciti, durante la formazione della lamina basale dei capillari, risultano compresi tra essa e le cellule endoteliali. I prolungamenti dei periciti spesso, superano la lamina basale, quando é presente, e contraggono stretti rapporti di contiguità con le cellule di Hofbauer. I periciti presentano un nucleo ovalare, a contorni regolari e cromatina addensata alla periferia del carioplasma. II citoplasma contiene numerosi piccoli mitocondri a creste lamellari e matrice chiara; é ricco di ribosomi aggregati alle cisterne di uno sviluppatissimo reticolo endoplasmatico granulare. Le cisterne del reticolo sono solitamente dilatate e ripiene di un materiale fioccoso, microgranulare, a media densità elettronica. Le caratteristiche ultrastrutturali e il numero di periciti possono variare. Metodiche istoimmunologiche, hanno evidenziato la sintesi di immunoglobuline della classe IgG (TEDDE, 1973). Ciò significa che anche la placenta può partecipare alla formazione delle immunoglobuline circolanti nel sangue fetale (MILLAR e Coll., 1972; JOHNSON e Coll., 1977a, 1977b), senza con questo escludere la possibilità di un loro passaggio nella circolazione fetale dal sangue materno attraverso la placenta, come ampiamente documentato in letteratura (GITLIN e Coll., 1964; KOHLER e FARR, 1966; GITLIN, 1973; OVARY, 1973). L'attività proteosintetica é limitata ai primi mesi di gravidanza, come risultato alle prove di immunofluorescenza. Questo dato è confortato da modifiche importanti come riduzione del reticolo endoplasmatico granulare e scarse cisterne appiattite e riduzione dei ribosomi e dei mitocondri. Anche il rapporto tra periciti e cellule endoteliali si modifica con allontanamento evidente delle due parti. ENDOTELIO DEI CAPILLARI FETALI 58
59 Lo spessore dell'endotelio dei capillari fetali varia notevolmente in relazione allo stato di contrazione o dilatazione del vaso. II calibro dei vasi risulta variabile e presenta maggior ampiezza durante il terzo trimestre di gravidanza (AHN e PANIGEL, 1964; LISTER, 1964 b). Nelle prime fasi della gestazione i capillari fetali occupano una posizione centrale nell'asse del villo e si spostano eccentricamente, ponendosi a ridosso dello strato trofoblastico man mano che avanza il periodo gestazionale. Al microscopio elettronico, le cellule endoteliali dimostrano una matrice citoplasmatica densa ed un voluminoso nucleo di forma ovalare. La superficie adluminale, presenta sottili evaginazioni irregolari che aumentano di numero col procedere della gravidanza. Nel citoplasma sono presenti scarsi organuli: piccoli mitocondri a matrice densa, sparse cisterne del reticolo endoplasmatico granulare, ribosomi riuniti in polisomi. Si osservano inoltre numerosi granuli di glicogeno e numerose vescicole pinocitotiche. Queste si formano sotto forma di invaginazioni sferoidali, caveolae, che conferiscono un caratteristico aspetto dentellato al profilo cellulare. Gli organuli citoplasmatici e le vescicole pinocitotiche divengono rare nelle placente a termine, mentre si conservano in gran quantità i granuli di glicogeno. Nel citoplasma endoteliale si osservano fascetti compatti di microfilamenti disposti parallelamente al maggior asse delle cellule endoteliali. Essi si trovano in contatto con i granuli di glicogeno e, attraverso indagini istoimmunologiche, sono risultati filamenti actinici (TEDDE, 1982). Secondo NICOLOV e SCHIEBLER (1981), nell'endotelio dei capillari fetali si possono riconoscere due tipi cellulari: cellule del primo tipo, ricche di filamenti di natura contrattile, e predominanti rispetto alle cellule del secondo tipo a funzione secretoria. Tali risultati mettono in evidenza la capacita contrattile delle cellule endoteliali. L'utilizzo di determinate sostanze vasoattive, in indagini condotte in vitro, hanno reso possibile la formulazione dell ipotesi che, a livello placentare, il flusso sanguigno e le stesse funzioni di trasporto abbiano una regolazione distrettuale che si esplicita anche a carico della componente vascolare più periferica. 59
60 FISIOLOGIA DELLA PLACENTA UMANA La placenta svolge diverse funzioni come la respirazione, il trasporto di materiale nutritizio verso il feto, l'eliminazione dei suoi metaboliti, e lo scambio endocrino. Viste le molteplici funzioni svolte dalla placenta, questa può essere considerata come organo analogo nello svolgimento dei compiti degli organi più disparati. Tuttavia la placenta è da considerare un organo completamente distinto da tutti gli altri. CIRCOLAZIONE PLACENTARE Una caratteristica molto importante dal punto di vista funzionale della placenta riguarda il sistema circolatorio materno e fetale (fig. 10). La mescolanza tra sangue materno e fetale normalmente non avviene; comunque alcune cellule sanguigne materne e fetali possono attraversare la barriera placentare e entrare nella circolazione parallela (BOYD e HAMILTON, 1970). I villi sono vascolarizzati dal sistema arterio-capillare-venoso che trasporta sangue fetale, e sono bagnati esternamente dal sangue materno che circola nello spazio intervilloso, interamente rivestito dal trofoblasto. Nei primi stadi dell'annidamento, si formano lacune che successivamente si aprono nei sinusoidi venosi materni dilatati a loro volta, connessi con il plesso venoso basale dell'endometrio. La circolazione inizialmente risulta molto lenta. Le pareti dei sinusoidi e dei capillari scompaiono progressivamente fino ad interessare le venule materne e le arteriole, mano a mano che l'invasione trofoblastica dell'endometrio procede. Attorno all'ottava settimana avvengono le prime sicure comunicazioni con lo spazio intervilloso mediante sottili fessure nel guscio trofoblastico. Le pareti delle arterie terminali presentano ipertrofia dei rivestimenti endoteliali ed alterazioni degenerative della loro muscolatura. Vi sono inoltre delle migrazioni di cellule citotrofoblastiche al loro interno che probabilmente hanno funzione nel controllo delle pressioni del sangue prima di giungere allo spazio intervilloso. 60
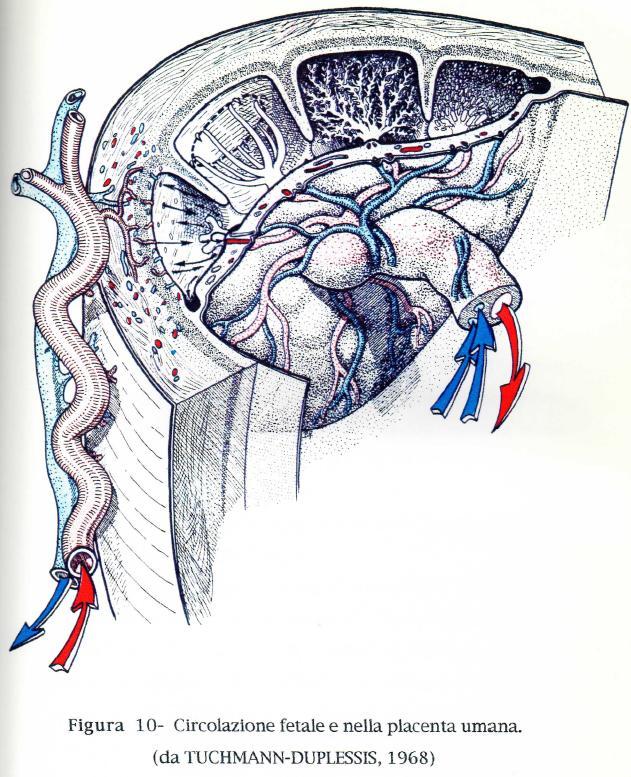
61 61
62 Gli sbocchi delle arterie dell endometrio negli spazi intervillosi della placenta a termine si trovano distribuiti sull'intera placca basale. Inoltre orifici delle vene provenienti dallo spazio intervilloso si trovano normalmente sopra tutta la superficie della placca basale e normalmente manca il seno marginale (HAMILTON e BOYD, 1951, 1960; RAMSEY, 1955, 1959; BOYD e HAMILTON, 1970). Vi è quindi un ampio drenaggio dello spazio intervilloso per tutta la superficie basale della placenta. Lo spazio intervilloso é dunque una complicata anastomosi artero-venosa contenuta entro il tessuto fetale rappresentato dal trofoblasto. La pressione del sangue arterioso al punto di entrata é stata stimata in mm di Hg. Quando il sangue raggiunge lo spazio intervilloso la sua pressione diminuisce a 10 mm di Hg nell'utero rilasciato, comunque questa può raggiungere i mm di Hg durante le contrazioni uterine. La pressione nelle vene uterine non supera gli 8 mm di Hg (WILKIN, 1965). Le contrazioni del miometrio possono quindi diminuire la facilita di passaggio del sangue dallo spazio intervilloso nelle vene uterine. II volume della placenta matura é di circa 500 ml di cui 350 ml sono ricoperti dal volume dei villi fetali. Lo spazio intervilloso può contenere circa 150 ml di sangue materno. La velocità del flusso utero-placentare aumenta durante la gravidanza da circa 50 ml al minuto a dieci settimane (MARTIN, 1965) fino a ml al minuto al termine (BROWNE e VEALL, 1953). È stato calcolato che il volume del sangue fetale che scorre attraverso i villi fetali é di circa 400 ml al minuto. Attraverso studi eseguiti su placente umane e di scimmia con la tecnica autoradiografica (RAMSEY, 1956; BORELL e Coll., 1958), si è dimostrato che iniettando materiale radioopaco nell'aorta, questo passa nella placenta, e che ogni singola arteria si distribuisce ad un lobo. Sembra che vi siano circa dodici arterie che penetrano nella placenta del macaco e 30 nella placenta umana. La pressione delle arterie sembra sufficiente a spingere profondamente il sangue nello spazio intervilloso, evitando cosi il passaggio diretto nelle vene. Nell'uomo, la struttura di uno spazio intervilloso non è compatibile con l'esistenza di un controflusso. Tuttavia la presenza di stretti canali fra i villi terminali potrebbe essere la via preferenziale che agisce in base al principio del 62
63 controflusso (MOSSMAN, 1959). Nei punti di scambio generalmente il sangue fetale scorre in direzione opposta rispetto a quello materno presente nei seni. Si é visto che i singoli villi sono bagnati da sangue materno di tipo arterioso, venoso o intermedio secondo la loro posizione. Questa disposizione é stata chiamata flusso multivilloso (BARTELS e Coll., 1962). Alcuni ricercatori hanno valutato la superficie dei villi esposti al sangue materno nello spazio intervilloso della placenta a termine. Le stime sulla estensione variano da 4 a 14 m 2 (WILKIN e BURSZTEIN, 1958). 63
64 BARRIERA PLACENTARE E TRASPORTO ATTRAVERSO LA PLACENTA Le strutture attraverso le quali avviene il passaggio di sostanze vengono chiamate barriere placentari o membrane placentari. Nella placenta umana la barriera é costituita da tessuti di origine fetale. È una membrana costituita dal trofoblasto dei villi liberi, dalla membrana basale, dallo stroma extravascolare dei villi, dalla membrana basale del capillare fetale e dalla parete del capillare. Esiste un'oscillazione riguardante lo spessore della barriera nelle diverse fasi della gravidanza e nelle diverse aree di ciascun villo. Negli stadi precoci della gravidanza lo spessore della membrana é di circa 0,025 mm ed è composta, come é stato su esposto, da sei strati: il rivestimento endoteliale del capillare fetale e la sua membrana basale, lo stroma mesenchimale del villo, il citotrofoblasto e la sua membrana basale, il sinciziotrofoblasto. Quest'ultimo risulta relativamente spesso nei primi quattro mesi di gestazione e si assottiglia col procedere della gravidanza fino a formare una sottile membrana nella fase terminale. Negli ultimi mesi di gravidanza i villi più piccoli presentano una membrana non più spessa di 0,002 mm che separa il sangue fetale dal sangue materno. Questa risulta costituita da cinque strati che persistono fino al termine di gravidanza: endotelio fetale, la sua membrana basale, il tessuto reticolare e il sincizio con la sua membrana basale. Risulta inoltre una notevole vicinanza tra la membrana basale endoteliale del vaso fetale e la membrana basale del sincizio, tale da essere indicata, come già si é detto, membrana "vascolo-sinciziale" (GETZOWA e SADOWSKY, 1959). Molti villi placentari esposti sulla superficie materna possono ricoprirsi di uno strato di materiale fibrinoide che può diminuire la permeabilità della barriera. Alcuni ricercatori hanno ritenuto che villi liberi nella placenta siano forniti di aree specializzate nel trasporto, intercalate a zone a cui spettano funzioni quali la sintesi e la secrezione. La microscopia elettronica, come già detto, ha rilevato due regioni sinciziali diverse, una sottile, la zona, e una più spessa, la zona (BURGOS e RODRIGUEZ, 1966; VECEK, 1974; FOX, 1978). La zona, dello spessore compreso tra i 2 e i 14, possiede numerosi microvilli irregolari impiantati sulla superficie basale irregolare. La superficie basale si invagina formando ampie cavità (pinosomi), vescicole e canalicoli. Le pareti dei pinosomi 64
65 sono più spesse di quelle delle strutture citoplasmatiche rivestite da membrana. Vi é oltre a una membrana unitaria trilaminare dello spessore di 80 A, uno strato di brevi filamenti sottili (il "terminal web") e un componente granulare, probabilmente un glicocalice, sulla superficie esterna (BOYD e Coll., 1968). Le vescicole e i vacuoli sono collegati con "canalicoli apicali". I vacuoli e le vescicole possono contenere granuli che presentano addensamenti centrali. Nella zona si possono trovare eccezionalmente alcuni componenti del reticolo endoplasmatico rugoso, ma i componenti canalicolari e vescicolari sono del tipo liscio. La zona presenta un reticolo endoplasmatico ben sviluppato e possiede numerose cisterne contenenti materiali a bassa densità elettronica. Alle membrane sono attaccati singoli ribosomi o poliribosomi. Abbiamo inoltre la presenza di numerosi microtubuli, lisosomi, pinosomi e gocciole lipidiche. La barriera comprende la membrana basale del sincizio e quella dell'endotelio capillare. Nonostante numerosi studi, non é tutt'oggi ben noto il meccanismo di assorbimento delle sostanze da parte del sinciziotrofoblasto. II passaggio delle diverse sostanze potrebbe avvenire sia per via trans-cellulare che per via extracellulare. TRASPORTO ATTRAVERSO LA PLACENTA Lo studio sulla permeabilità placentare é legato ai sistemi enzimatici. Essi comprendono la fosfatasi alcalina e acida, la colinesterasi ed enzimi proteolitici e glicolitici (PAGE e GLANDENING, 1955; PAGE,1957). Le sostanze in grado di attraversare la barriera placentare sono state suddivise in 4 gruppi. II primo gruppo comprende sostanze in grado di attraversare la barriera per semplice diffusione. Queste sostanze sono l'h20, l O2 e gli elettroliti essenziali per la vita. II secondo gruppo comprende sostanze necessarie alla nutrizione del feto, quali glucosio, amminoacidi, lipidi e vitamine (schema 6). Queste sostanze attraversano la barriera mediante trasporto attivo. II terzo gruppo é costituito da sostanze che regolano l'attività metabolica; il meccanismo del loro trasferimento non é ben noto, comunque si ritiene che alcune di esse (tiroxina, idrocortisone e gonadotropina) diffondono lentamente attraverso la placenta. II quarto gruppo 65
66 comprende macromolecole di importanza immunologica. II trasporto tra spazio intervilloso e sistema vacuolare del sincizio può avvenire per pinocitosi. Vi sono delle variazioni nella velocità del trasporto nelle due direzioni, evidenziato dalla diversa concentrazione di alcune sostanze nel sangue fetale e in quello materno (VILLEE, 1960; DANCIS, 1962). La concentrazione maggiore di alcuni costituenti nella circolazione fetale é ovviamente a vantaggio per il feto. II passaggio avviene in presenza di un gradiente di concentrazione; questo necessita un trasporto attivo transplacentare, che richiede un consumo di energia metabolica (SCHREINER, 1965). II passaggio di ossigeno e anidride carbonica attraverso la barriera placentare risulta ovviamente fondamentale per la vita del feto. Lo scambio di questi gas avviene per semplice diffusione. La quantità di ossigeno trasferita dipende per gran parte dalla differenza fra pressione parziale di ossigeno nel sangue materno e in quello fetale, velocità del flusso di sangue nello spazio intervilloso, del flusso di sangue nei villi coriali, lo spessore e l'area della superficie della barriera placentare. I gas attraversano la barriera placentare per effetto dei gradienti di diffusione misurati al termine di gravidanza, di circa 20 mm di Hg per l'ossigeno dalla madre al feto, e di 5 mm di Hg per l'anidride carbonica dal feto alla madre (SEED, 1968). II trasferimento risulta facilitato per la maggiore affinità dell'emoglobina fetale per l'ossigeno. Nell'utero sono state riscontrate pressioni parziali di ossigeno minori e di anidride carbonica maggiori di quelle osservate dopo la nascita (WULF, 1962). ACQUA Anche l'acqua risulta un costituente essenziale per la vita cellulare. Anche questo elemento diffonde attraverso la barriera placentare. È stato dimostrato che fra la quattordicesima settimana e il termine della gravidanza la velocità di passaggio per unità di peso della placenta cresce di cinque volte (HELLMAN e Coll., 1948). 66
67 II passaggio più consistente di H2O per unità di peso placentare si ha alla 36 a settimana a cui segue un rapido declino durante le ultime settimane di gravidanza (FLEXNER e Coll., 1948). Verso la fine della gravidanza il feto riceve più di 3 litri e mezzo di H2O all'ora e una quantità quasi equivalente ritorna alla circolazione materna (HUTCHINSON e Coll., 1959; SCOGGIN e Coll, 1964). 67
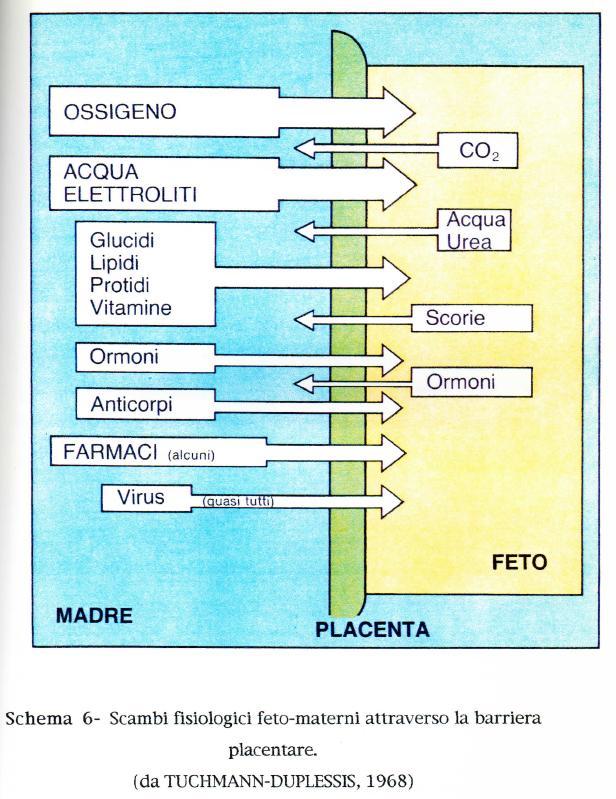
68 68
69 ELETTROLITI Elettroliti come sodio, potassio, magnesio, calcio e fosforo attraversano più o meno facilmente la placenta. Calcio e fosforo si trovano in concentrazioni più elevate nel sangue fetale e questo riduce la mobilità rispetto agli altri ioni. Si è osservato un notevole aumento di permeabilità della placenta umana al sodio radioattivo col progredire della gestazione (FLEXNER e Coll., 1948), (grafico 3). II passaggio più consistente per unita di peso della placenta si osserva alla trentaseiesima settimana. Segue un rapido declino durante le ultime settimane di gravidanza. II livello di ferro contenuto nel siero fetale é di due o tre volte superiore a quello materno; quindi tenuto conto che la capacità del siero fetale di legare il ferro, non é maggiore di quello materno, indica che la barriera placentare può concentrare selettivamente questo ione dalla parte fetale (SEEDS, 1968). 69
70 70
71 NUTRIZIONE DEL FETO Concentrazioni maggiori di amminoacidi essenziali e polipeptidi semplici sono state riscontrate nel plasma fetale rispetto a quello materno; questo dovrebbe indicare l'esistenza di un processo attivo (KRYSZTOPOSKY, 1949). Questa maggior concentrazione facilita una rapida sintesi delle proteine e una conseguente rapida crescita del feto (CHRISTENSEN e STREICHER, 1948). L'esistenza di una forma di trasporto é stata dedotta dall'utilizzo di un isomero ottico dell'amminoacido istidina, utilizzato in egual misura nelle due forme D ed L sulla superficie materna della barriera placentare. Nei nati 10 minuti dopo il rilevamento si é osservato che era maggiore la concentrazione dell'isomero L rispetto al D non esistente in natura. Poiché i due isomeri dello stesso aminoacido non diffondono attraverso la placenta alla stessa velocità, si è pensato ad un trasporto che faciliti il passaggio della forma naturale (PAGE e Coll., 1957). Esperimenti su animali hanno rivelato il passaggio rapido attraverso la placenta di aminoacidi marcati contro gradiente di concentrazione; questo può avvenire per mezzo di un meccanismo di trasporto che sfrutta energia metabolica (DANCIS e SHAFRAN, 1958). Esiste anche una competizione nel passaggio degli aminoacidi (CHRISTENSEN e STEICHER, 1948). AMMINOACIDI, POLIPEPTIDI E PROTEINE Generalmente é accettato il passaggio di amminoacidi attraverso la placenta dalla circolazione materna a quella fetale. Gli amminoacidi sono utilizzati per la sintesi delle proteine del plasma da parte del feto (DANCIS, 1960, 1962). II passaggio di proteine risulta fondamentale per la nutrizione del feto e per il trasporto di proteine come anticorpi e antigeni in quantità immunologicamente efficaci. 71
72 L'alimentazione proteica materna, in effetti, rappresenta la maggiore fonte di amminoacidi per feto. Un diminuito apporto proteico materno,prima e durante la gravidanza, può essere considerato fattore di ritardo nell'accrescimento fetale. LIPIDI Gli acidi grassi liberi sono trasferiti rapidamente nelle due direzioni attraverso la barriera placentare (DANCIS, 1969). L'entità del flusso transplacentare varia in relazione alla concentrazione di acidi grassi nel versante materno (ELPHICK e Coll., 1978). Nel coniglio e nell'uomo é stato osservato che l'acido arachidonico viene impegnato nel circolo placentare in quantità maggiori di quanto ci si aspetterebbe dalla concentrazione degli acidi grassi del sangue materno. La placenta, inoltre, assume acidi grassi dai fosfolipidi e da esteri del colesterolo del sangue materno ed é in grado di convertire l'acido linoleico in acido arachidonico. I grassi neutri sembrano passare attraverso la placenta umana mediante pinocitosi e perciò alcuni lipidi fetali sono di origine materna (YAMAGUCHI e Coll., 1960, 1964). II sangue materno contiene una concentrazione di grassi e sostanze lipidiche considerevolmente più alta di quella del feto. In ogni caso il contributo materno agli acidi grassi fetali varia in funzione della dieta. GLUCIDI II glucosio è il substrato principale per il metabolismo ossidativo del feto e risulta anche un importante precursore per la sintesi di glicogeno e di lipidi del feto. La placenta contiene grandi quantità di glicogeno, gran parte del quale é sintetizzato dal glucosio materno. La concentrazione di glicogeno placentare raggiunge la sua punta massima all'8 a settimana di gestazione, per poi declinare con l'avanzare della gravidanza; tale diminuzione é accompagnata da un corrispondente aumento del glicogeno del fegato fetale (LONGO,1972). 72
73 II passaggio di glucosio attraverso la placenta é in relazione alla concentrazione di questo e dalla disponibilità di carrier. Inoltre l'entità del trasporto é direttamente proporzionale alla superficie placentare e inversamente proporzionale al suo spessore. II feto comunque riceve il 75% del suo fabbisogno energetico dal glucosio di origine materna e lo stesso metabolismo intrinseco placentare é largamente glucosio dipendente. 73
74 LA PLACENTA COME ORGANO DI ESCREZIONE Nei neonati mancanti dei due reni, il prodotto del metabolismo dell'azoto può essere trasferito al sangue materno attraverso la barriera placentare. L'urea, la creatinina e la creatina passano attraverso la barriera placentare per semplice diffusione. La concentrazione dell'urea nel liquido amniotico é molto maggiore che nel sangue fetale. La bilirubina del plasma fetale si trova tutta nella forma libera, non coniugata, ed è probabilmente escreta per diffusione attraverso la placenta. La bilirubina coniugata presente nel plasma materno non passa nella circolazione fetale. ORMONI Steroidi coniugati attraversano facilmente la barriera placentare. Il livello di estriolo e quello di progesterone sono più alti nel plasma fetale rispetto a quello materno (AITKEN e Coll., 1958). Al secondo mese di gravidanza, queste sostanze sono elaborate dalla placenta. Al termine i livelli dei corticosteroidi nella madre e nel feto sono simili (GAMZELL, 1954; MIGEON e Coll., 1956). Tirosina marcata con isotopi radioattivi attraversa lentamente la barriera placentare (GRUNBACH e WERNER, 1956; MYANT, 1958). Anche gli androgeni possono attraversare la placenta (BURNS, 1961). Alcuni autori hanno trovato che la placenta é impermeabile all'insulina (ROSA e FANARD, 1958; BUSE e Coll., 1962; SPELLACY e Coll., 1964). Esperimenti con insulina marcata con I 131, hanno dimostrato che questa attraversa facilmente la barriera nelle due direzioni (GITLIN e Coll., 1965). Gli ormoni sintetizzati dalla placenta sono le gonadotropine e gli steroidi. Sono 4 gli ormoni di produzione placentare: le gonadotropina corionica umana (HCG), i lattogeni placentari umani (HPL), estrogeni e progesterone (grafico 4 e 5). Accanto a questi ormoni la placenta é in grado di produrne altri il cui significato funzionale è incerto, quali: l'ormone tireotropico-corionico (simile al TSH) e la proopio-melanocortina, precursore dell'acth, dell'msh e delle endorfine. 74
75 75
76 La sintesi della gonadotropina corionica umana (HCG) viene condotta dal sinciziotrofoblasto placentare. Durante la gravidanza la concentrazione dell'ormone risulta superiore nel sangue materno rispetto a quello fetale. La secrezione di questo ormone é stata da alcuni autori localizzata nel sincizio, altri pensano che il luogo di secrezione sia il citotrofoblasto. La comparsa dell'hcg nelle donne gravide avviene nella seconda settimana dopo l'ovulazione (grafico 4). L'HCG é una glicoproteina con azione luteotrofica simile a quella dell'lh ipofisario, capace di determinare nell'ovaio lo sviluppo del corpo luteo e di stimolarne la secrezione di progesterone e di estrogeni. II lattogeno della placenta umana (HPL) é sintetizzato dalla placenta. È presente in concentrazione più alta nella placenta che nel siero materno e la sua concentrazione cresce durante la gravidanza, raggiungendo il massimo nell ultimo trimestre (SIMMER, 1968). L'ormone é sintetizzato dal sinciziotrofoblasto placentare, dimostrato dalla sua produzione anche in assenza del feto; esso passa esclusivamente nella circolazione materna (KAPLAN e GRUMBACH, 1965). ORMONI STEROIDEI La produzione degli ormoni steroidei da parte del sinciziotrofoblasto é stata ampiamente dimostrata. II progesterone può essere ottenuto dalla placenta umana a tutti gli stadi della gestazione (DICZFALUSY,1952; SALHANICK e Coll., 1952; DICZFALUSY e TROEN, 1961; RYAN, 1962). II progesterone, necessario per la prosecuzione della gravidanza, nel secondo e terzo trimestre viene prodotto in maggior parte dalla placenta. Infatti, la rimozione del corpo luteo dopo il trentaquattresimo giorno di gravidanza, non produce aborto né diminuisce significativamente il pregnadiolo urinario (DICZFALUSY e BORELL, 1961), (grafico 5). II pregnadiolo urinario materno continua a crescere anche dopo la morte del feto, il quale perciò partecipa in scarsa misura alla secrezione del progesterone placentare. Le funzioni del progesterone sono quelle di inibire la motilità della muscolatura uterina, permettendo un adattamento dell'utero ai progressivi accrescimenti del 76
77 feto e di stimolare la formazione e la crescita della decidua endometriale, assicurando cosi la connessione anatomica tra madre e feto. Gli estrogeni sono stati isolati dalla placenta ed il loro tasso urinario aumenta durante la gravidanza, essendo strettamente associati allo sviluppo placentare (grafico 5). Ricerche in vitro dimostrano essere secreti dal sinciziotrofoblasto. II fatto che l'estrogeno non sia secreto dalle ovaie materne é dimostrato dal fatto che in seguito ad ovariectomia bilaterale eseguita oltre la sesta settimana di gravidanza, non si nota una diminuzione significativa della quantità di estrogeno escreta. Dopo l'espulsione della placenta, l'escrezione di estrogeno diminuisce rapidamente. Inoltre, in seguito a rimozione del feto, o alla sua morte, l'escrezione dell'estrogeno cade rapidamente. Quindi la placenta si avvale del feto per sintetizzare estrogeni, ma non progesterone (VILLEE e TSAI, 1969). Gli estrogeni placentari sono rappresentati per il 90% dall'estriolo. Questi ormoni permettono uno sviluppo dimensionale dell'utero, un aumento delle dimensioni vaginali e dei genitali esterni e lo sviluppo delle mammelle e della ghiandola mammaria. VITAMINE II passaggio placentare delle vitamine è molto complesso e generalmente non é un processo di semplice diffusione (WILKIN, 1958). Le vitamine idrosolubili attraversano la placenta con maggiore facilita rispetto a quelle liposolubili. La placenta può accumulare vitamine B, D ed E, e può partecipare attivamente al metabolismo dell'acido ascorbico e del Flavin-Adenin-Dinucleotide. Si é dimostrata, inoltre, la presenza della vitamina C nel sinciziotrofoblasto e nello stroma dei villi, e la vitamina A nei villi (PAPADIA, 1956). La vitamina A liposolubile ha concentrazione inferiore nel sangue fetale che in quello materno, mentre la vitamina D e le idrosolubili hanno concentrazioni più alte nel sangue fetale che nel sangue materno. Questo é in favore di un meccanismo di trasporto attivo contro gradiente, con consumo di energia metabolica. 77
78 FARMACI II transito dei farmaci attraverso la barriera placentare è stato ampiamente documentato da numerosi autori. II peso molecolare dei medicamenti é molto importante per la determinazione della velocità di transito. I farmaci con peso molecolare inferiore a 600 passano in modo relativamente facile (MOYA, TJORNDYKE, 1962), mentre quelli con peso molecolare superiore a 1000 non passano in maniera apprezzabile. Si può dire che tutti i farmaci ingeriti o iniettati dalla madre raggiungono il feto, sebbene farmaci di alto peso molecolare siano trasmessi a velocità inferiori rispetto a farmaci con basso peso molecolare (APGAR, 1966). La maggior parte dei farmaci di uso generate ha un basso peso molecolare e quindi attraversano la barriera placentare per diffusione, sebbene alcuni passino per trasporto attivo (HAGERMAN e VILLEE, 1960). Questi farmaci includono sedativi, tranquillanti, ipnotici, narcotici, anestetici (locali e generali), miorilassanti, agenti chemioterapici e antibiotici. IMMUNOLOGIA La capacita di riconoscimento e di rigetto di cellule ed organi estranei é una caratteristica di tutti i vertebrati superiori. Nel sangue materno circolano una serie di cellule linfocitarie in grado di riconoscere e neutralizzare determinati antigeni dei gruppi sanguigni incompatibili del feto, o produrre anticorpi contro antigeni trofoblastici. Tuttavia, il rigetto della placenta e del feto non avvengono. L'embrione e la sua placenta rappresentano un tipo particolare di trapianto, a causa del corredo genetico di derivazione sia paterna che materna. Se consideriamo solo il corredo genetico materno, il feto é un isotrapianto, mentre se consideriamo quello paterno, é un omotrapianto. Nel corso dello sviluppo umano il sinciziotrofoblasto é esposto direttamente ai tessuti materni, per cui qualsiasi antigene solubile di origine fetale può passare facilmente nel sangue materno. Inoltre, qualsiasi anticorpo può agire attraversando la barriera placentare. 78
79 Nonostante la grande superficie del corion, al termine di gravidanza, non si spiega l'apparente neutralità di questo e la sua relazione con il sistema immunitario. Una reazione primaria di rigetto inizia con un'identificazione del tessuto estraneo da parte dei linfociti circolanti; poi vi e una risposta proliferativa negli organi linfoidi e infine il tessuto estraneo viene distrutto da speciali linfociti. Nel caso della placenta, il ruolo degli anticorpi nel rigetto del trapianto, se c'é é molto scarso e inefficace (NAJARIAN e FOKER, 1969; WILLIAMS e Coll., 1969). Sembra che la madre produca una risposta immunitaria insufficiente verso la blastocisti nel corso dell'impianto. Nel periodo dell'impianto, piccole cellule di tipo linfocitario si infiltrano nell'endometrio che circonda l'unita feto-placentare. Sono stati invocati sei fattori ipotetici per spiegare la ritenzione dell'unita fetoplacentare come omoinnesto nella madre: 1) Rivestimento del trofoblasto antigenico da parte di una barriera acellulare; 2) Non antigenicità del trofoblasto; 3) Utero come luogo immunologicamente privilegiato; 4) Gli ormoni materni e placentari; 5) Tolleranza immunologica del feto da parte della madre; 6) Aumento di capacita immunologica. Finora non é stata data alcuna spiegazione completamente soddisfacente per spiegare la relazione unica che esiste tra la madre e il feto. 79
80 80
81 MATERIALI E METODI Le ricerche sono state condotte su 73 placente umane in età gestazionale compresa tra la 6 e la 12 settimana e al termine di gravidanza. Le placente precoci sono state ottenute da interruzione volontaria di gravidanza. Alcuni frammenti di placenta, dopo il prelievo ottenuto con strumenti microchirurgici, sono stati fissati nei liquidi di Carnoy, Bouin, formalina al 10%, disidratati con una serie di passaggi in etanolo a concentrazione crescente, chiarificati in xilolo ed inclusi in paraffina. Le sezioni microtomiche sono state colorate con ematossilina-eosina, le tecniche di PaS e blu di toluidina. Altri frammenti sono stati fissati a 4 C in aldeide glutarica al 2,5%, in tampone fosfato a ph 7,4 e tetrossido di osmio all'1%; disidratati con etanolo ed ossido di propilene ed inclusi in Durcupam ACM (Fluca AG). Le sezioni semifini (1-1,5 micron), sono state colorate con blu di toluidina, PAS, Feulgen e sono state studiate al microscopio ottico. Le sezioni ultrafini ( A di spessore), sono state contrastate con acetato di uranile e citrato di piombo ed osservate al microscopio elettronico in trasmissione Zeiss EM 109. Infine alcuni frammenti sono stati sottoposti a tecniche di immunoistochimica. I campioni sono stati fissati in formalina neutra tamponata, disidratati in etanolo, chiarificati in xilolo e inclusi in paraffina. Le sezioni microtomiche ottenute, di circa 3 micron, sono state distese su vetrini portaoggetto polilisinati e fatte aderire ad essi. Sono state, inoltre, sparaffinate con 2 passaggi in xilolo per 5 minuti e dei passaggi in etanolo a concentrazione decrescente. Le sezioni sono state in seguito reidratate attraverso un lavaggio in acqua distillata e un bagno in PBS 0,01 M a ph 7,4. I vetrini, in seguito, sono stati posti in camera umida e le sezioni ricoperte con perossido di idrogeno al 3% e incubate per 5 minuti allo scopo di eliminare le perossidasi endogene. Dopo questo passaggio i vetrini sono stati immersi in un bagno con PBS per due minuti; a questo punto le sezioni sono state cimentate con il reagente bloccante (siero anti capra o anti topo), con una incubazione di 20 minuti. È stato eliminato 81
82 l'eccesso di reagente senza lavare i vetrini e si è applicato l'anticorpo primario (gli anticorpi anti-laminina (SIGMA e PABISCH) alle sezioni e si é lasciato incubare per 60 minuti. Dopo un lavaggio in PBS é stato applicato l anticorpo secondario biotinilato ed incubato per 20 minuti. Infine, le sezioni sono state ricoperte con il complesso Avidina-Biotina-Perossidasi e incubate per 20 minuti. È stato poi applicato per 20 minuti un Reagente Cromogeno costituito da: tampone acetato, AEC e perossido di idrogeno al 3%. I vetrini, lavati con acqua deionizzata per 5 minuti e contrastati con ematossilina di Mayer per 2 minuti, sono stati montati con gelatina glicerinata ed osservati al microscopio ottico. 82
83 RISULTATI Verranno esposti i risultati dell'osservazione di preparati allestiti da sezioni in paraffina, con le routinarie metodiche di colorazione e con metodiche di immunoistochimica, e di sezioni ultrafini allestite per la microscopia elettronica. OSSERVAZIONI AL MICROSCOPIO OTTICO Placente alla 6 a -7 a settimana. L'osservazione dei preparati mostra numerosi villi coriali con i caratteristici elementi costitutivi: lo strato periferico rappresentato dal sinciziotrofoblasto e dal citotrofoblasto, lo stroma del villo e i vasi capillari sanguigni (fig. 1). In questa fase é caratteristico il fatto che gli elementi del citotrofoblasto formano uno strato continuo al di sotto del sincizio. Nei preparati allestiti con la tecnica del PaS, solo di rado é possibile osservare la membrana basale del trofoblasto che è presente solo in qualche tratto del contorno del villo (fig. 2). Utilizzando la tecnica immunoistochimica utilizzata per evidenziare la laminina, questa risulta presente in corrispondenza del versante interno del trofoblasto e intorno ai singoli elementi del citotrofoblasto; questi ultimi mostrano una evidente positività nella zona periferica del citoplasma, mentre appare negativa la regione perinucleare e il nucleo stesso (fig. 7). Placenta alla 12 a -13 a settimana. Col progredire della gravidanza, si modifica l'assetto delle componenti del villo coriale a livello del quale gli elementi del citotrofoblasto formano uno strato discontinuo, profondamente rispetto al sincizio; quest'ultimo presenta una spiccata acidofilia citoplasmatica, numerosi nuclei a cromatina addensata e una superficie esterna notevolmente irregolare (fig. 3). Di frequente si notano vasi capillari sanguigni immersi nello stroma del villo e prevalentemente in posizione eccentrica (fig. 3). Nei preparati allestiti con la tecnica del PaS é possibile notare una evidente membrana basale del trofoblasto che costituisce uno strato continuo 83
84 internamente agli elementi del citotrofoblasto o, nelle zone in cui essi mancano, direttamente nel sinciziotrofoblasto (fig. 4). Anche la sostanza intercellulare dello stroma del villo appare più intensamente PaS-positiva e con un più alto grado di polimerizzazione (fig. 4). La metodica di immunoistochimica per l'evidenziazione della laminina ha dato una positività sia a livello della membrana basale, sia nei contorni degli elementi del citotrofoblasto, sia in alcune introflessioni presenti a livello del sinciziotrofoblasto, sul versante interno (fig. 8). Placente a termine. In questa fase i villi coriali appaiono di solito di calibro minore; si é ridotto lo spessore del trofoblasto, sono sempre più rari gli elementi del citotrofoblasto e appaiono molto numerosi i vasi capillari nel contesto dello stroma (fig.5). Nei preparati allestiti con la tecnica del PaS appare molto evidente una netta membrana basale che assume talvolta un andamento ondulato e si connette con la sostanza intercellulare dello stroma del villo, anch'essa notevolmente polimerizzata (fig. 6). OSSERVAZIONI AL MICROSCOPIO ELETTRONICO Nel riportare queste osservazioni verrà fatta una descrizione degli elementi costitutivi il villo coriale, che pur andando incontro a modificazioni durante il corso della gravidanza, mantengono delle caratteristiche abbastanza omogenee. Relativamente alle variazioni che si verificano nel corso della gravidanza, verrà fatto particolare riferimento alle modificazioni a cui va in contro la membrana basale del trofoblasto. Lo studio di sezioni ultrafini permette di chiarire le caratteristiche strutturali degli elementi sinciziali; risalta infatti la ricchezza in organuli rappresentati da mitocondri prevalentemente a creste tubulari, cisterne del reticolo endoplasmatico granulare ed granulare e complessi di Golgi. Sono inoltre presenti inclusi lipidici (fig. 9). 84
85 II plasmalemma del versante apicale costituisce un gran numero di microvillosità che permettono abbondantissimi scambi con il sangue materno in cui i villi pescano (fig. 9). Anche nella porzione profonda, a livello della zona di contatto con gli elementi del citotrofoblasto, il plasmalemma sinciziale si solleva formando numerosi microvilli che aggettano in canalicoli che circondano gli elementi del citotrofoblasto (fig. 9). Nello strato sinciziale é possibile talvolta notare dei canalicoli a direzione perpendicolare rispetto alla superficie, connessi tra loro da sistemi giunzionali in cui aggettano estroflessioni citoplasmatiche. Spostandosi nella regione più profonda, é possibile osservare le caratteristiche ultrastrutturali delle cellule di Langhans (citotrofoblasto): esse appaiono infatti sotto forma di voluminosi elementi chiari, i cui organuli citoplasmatici sono rappresentati da mitocondri allungati a creste lineari, da cisterne sparse del reticolo endoplasmatico granulare e da complessi di Golgi in sede sovranucleare. Sono numerosi i ribosomi liberi e talvolta una grande quantità di glicogeno (fig. 9). Per quanto riguarda la membrana citoplasmatica, essa si presenta molto irregolare per la presenza di numerosissime microvillosità che sporgono nel sistema canalicolare che circonda gli elementi del citotrofoblasto (fig. 9). In alcune regioni sono presenti apparati funzionali di tipo desmosomiale, sia tra gli stessi elementi del citotrofoblasto che tra elementi del citotrofoblasto ed elementi sinciziali. Al di sotto dello strato di derivazione trofoblastica è possibile osservare una membrana basale di cui verranno descritte le caratteristiche nelle varie fasi di gravidanza. Infine, in posizione variabile rispetto alle formazioni superficiali del villo, troviamo i vasi capillari. Nelle placente intorno alla 6 a -7 a settimana di gravidanza, sulla lamina basale poggiano le cellule del citotrofoblasto che formano, come già detto, uno strato continuo al di sotto del sinciziotrofoblasto (figg. 10 e 11). In questa fase della gravidanza la lamina basale é esile e nel suo contesto non si riconoscono strutture formate; il suo decorso é regolare e parallelo al plasmalemma delle cellule di Langhans che non presentano che scarse e tozze estroflessioni 85
86 citoplasmatiche (fig. 10). Nelle microfotografie elettroniche si può anche osservare che la lamina basale passa a ponte da una cellula di Langhans all'altra, laddove sono presenti canalicoli intercellulari estremamente irregolari. Durante questa fase della gravidanza non si osservano rapporti diretti tra lamina basale trofoblastica e capillari fetali; questi ultimi infatti mantengono ancora nel villo una posizione centrale e risultano, peraltro, essere privi di una loro propria lamina basale. Intorno alla 12 a -13 a settimana di gravidanza si modificano i rapporti fra elementi trofoblastici e lamina basale, nel senso che le cellule del citotrofoblasto si distanziano e tra di esse si interpongono dei lembi di sinciziotrofoblasto, per cui il versante profondo dello strato epiteliale risulta essere formato sia dagli elementi del citotrofoblasto che dal sincizio. Quest'ultimo é caratterizzato dal fatto che il plasmalemma si estroflette in numerose, sottili e lunghe microvillosità che pescano nel cosiddetto spazio subsinciziale (fig. 12); la lamina basale risulta pertanto essere in rapporto con queste estroflessioni citoplasmatiche. Dal punto di vista ultrastrutturale la lamina risulta essere ancora costituita da due strati, uno trasparente ed uno denso, e quest'ultimo presenta nel suo contesto formazioni microfilamentose che decorrono parallele alla superficie del villo; con la lamina basale prendono rapporto stretto le microvillosità del sincizio (fig. 13). In questa fase si modifica la posizione dei capillari fetali che tendono ad occupare, nelle sezioni trasversali dei villi, una posizione eccentrica. L'endotelio presenta una sottile lamina basale discontinua che tende a fondersi con la lamina basale del trofoblasto. Una caratteristica da sottolineare é la presenza di fibrille collagene che aderiscono alla superficie profonda della lamina basale del trofoblasto (nell'insieme questa struttura corrisponde alla membrana basale della microscopia ottica), (fig. 13). Al termine della gravidanza la lamina basale del trofoblasto presenta un aspetto per lo più microfibrillare e molto spesso, nel suo contesto, si osservano delle fibrille collagene (fig. 14). In alcuni punti é possibile individuare diversi piani microfibrillari paralleli e sovrapposti. Nei forti ingrandimenti é possibile rilevare come le strutture microfibrillari siano in stretto rapporto con le fibrille collagene, 86
87 rappresentando per queste ultime quasi un sistema di ancoraggio. La densità della lamina basale é più accentuata rispetto ai precedenti periodi e le microvillosità del versante profondo del sinciziotrofoblasto, in questa fase più irregolari, lunghe, a decorso sinuoso, spesso penetrano per un tratto discreto nel suo spessore, contraendo quindi con essa rapporti sempre più stretti (fig. 14). Molto numerose sono, peraltro, le fibrille collagene che aderiscono, in fascetti più o meno compatti, alla superficie profonda della lamina basale. Si osserva costantemente, nei preparati, una fusione della lamina basale trofoblastica con la lamina basale dei capillari sanguigni fetali, situati a ridosso dello strato epiteliale, a costituire la già citata membrana vasculo-sinciziale. Va in ultimo sottolineato che data l estrema rarità delle cellule di Langhans residue, la lamina basale resta in rapporto, quasi esclusivamente, con il sinciziotrofoblasto. 87
88 CONCLUSIONI Dai dati su esposti, sia bibliografici che sperimentali, risulta evidente che il villo coriale va incontro a notevoli trasformazioni nel corso della gravidanza. Accanto alle trasformazioni strutturali del sincizio e del citotrofoblasto, molto evidenti sono le modificazioni a cui va incontro la membrana basale del trofoblasto. In particolare a livello di essa sono evidenti due fatti principali: 1) le caratteristiche strutturali ed ultrastrutturali della lamina basale del trofoblasto non sono le stesse durante tutto l'arco della gravidanza, ma variano nei diversi periodi; 2) varia anche lo spessore della membrana stessa in maniera caratteristica, tanto che é possibile individuare due fasi distinte della gravidanza, durante le quali lo spessore rimane omogeneo. Mentre nei primi mesi la lamina basale, che all'osservazione al microscopio ottico appare PaS negativa, è costituita da materiale astrutturato a scarsa densità elettronica, con il progredire della gravidanza si assiste alla comparsa di una trama delicata di microfilamenti. L'immagine corrispondente al microscopio ottico é una membrana basale nettamente PaS positiva. Nelle prime fasi della gravidanza essa appare costituita da laminina, a cui si aggiunge in tempi successivi il collagene IV. La laminina é stata osservata anche sotto forma di granuli, a livello degli elementi del citotrofoblasto e negli spazi canalicolari che circondano questi elementi. In considerazione del fatto che la parete del villo coriale risulta attraversata da sistemi canalicolari che permettono il transito di sostanze dal versante esterno all'asse mesenchimale, e che questi canalicoli sono ampiamente comunicanti con i canalicoli che contornano le cellule del citotrofoblasto, la presenza di laminina, a questo livello, giocherebbe un ruolo importante nella regolazione del passaggio attraverso la barriera placentare. Per quanto riguarda lo spessore della membrana basale, come noto dalla letteratura (TEDDE e DELRIO, 1980) esso rimane pressoché costante fino al 4-5 mese per restare praticamente invariato fino al termine. Mentre in numerose patologie la modificazione e l'ispessimento della membrana basale é legato ad alterazioni molto profonde delle interazioni stroma parenchima (MARTINEZ- HERNANDEZ e Coll., 1983), per quanto riguarda la placenta, l'incremento brusco 88
89 di spessore si verifica in concomitanza con l acquisizione, da parte del corpo fetale, di funzioni assenti fino a quel momento (attività secretoria delle principali ghiandole endocrine, efficienza dei sistemi di difesa specifici ecc...). Quindi si potrebbe pensare che a livello placentare, in questo stadio, si differenzino strutture che determinano, con la loro presenza e per le loro caratteristiche, una migliore selettività degli scambi materno-fetali. Si realizzerebbe una condizione simile a quella che si realizza a livello del glomerulo renale in cui unico elemento realmente selettivo nel passaggio delle sostanze attraverso la barriera di filtrazione é la membrana basale. Sono note peraltro condizioni particolari in cui sia lo stato d essere della madre (fumo, patologie particolari), sia condizioni sperimentali, possono modificare l'assetto della membrana basale stessa (TEDDE e Coll., 1991; VAN DER VELDE, 1985). Da sottolineare, infine, che in base ai nostri risultati potrebbero giocare un ruolo importante nella costituzione della membrana basale le cellule del citotrofoblasto a livello delle quali é stata notata una positività alla laminina. Con il procedere della gravidanza si realizzerebbe una specializzazione della membrana basale con la possibilità di una interazione profonda tra sangue fetale e sangue materno. Deriva da questo che la membrana basale debba essere considerata un elemento importante nella costituzione della «barriera placentare». 89

90 MATERIALE ICONOGRAFICO 90

91 91

92 92
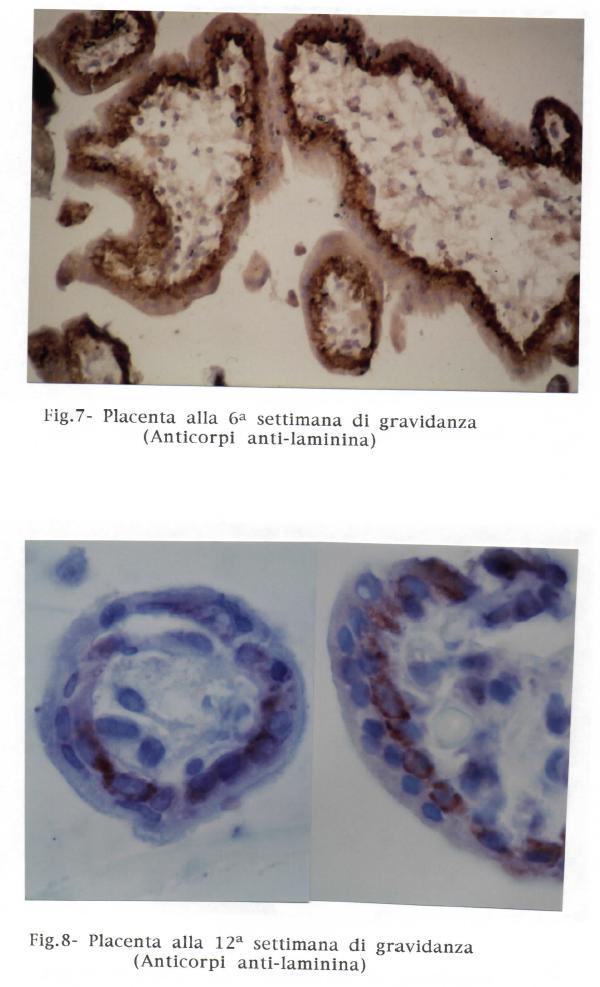
93 93

94 94
95 95
96 96
UNIVERSITA DI BARI ALDO MORO
 UNIVERSITA DI BARI ALDO MORO Scuola di specializzazione in Fisiopatologia della riproduzione degli animali domestici (Direttore: Prof. Giovanni M Lacalandra) Disciplina: Morfologia ed anatomia topografia
UNIVERSITA DI BARI ALDO MORO Scuola di specializzazione in Fisiopatologia della riproduzione degli animali domestici (Direttore: Prof. Giovanni M Lacalandra) Disciplina: Morfologia ed anatomia topografia
Il concepimento (fecondazione) avviene circa 14 gg prima delle mestruazioni e subito dopo l'ovulazione.
 avviene circa 14 gg prima delle mestruazioni e subito dopo l'ovulazione.") Il concepimento (fecondazione) avviene circa 14 gg prima delle mestruazioni e subito dopo l'ovulazione. Se i cicli sono irregolari, la data del concepimento e quindi la durata della gravidanza e l'epoca
Il concepimento (fecondazione) avviene circa 14 gg prima delle mestruazioni e subito dopo l'ovulazione. Se i cicli sono irregolari, la data del concepimento e quindi la durata della gravidanza e l'epoca
La Circolazione ALESSIA JULIANO MAX RICCARDO 2^A TRIUGGIO
 La Circolazione ALESSIA JULIANO MAX RICCARDO 2^A TRIUGGIO APPARATO CARDIOCIRCOLATORIO CUORE ARTERIE VENE CAPILLARI SANGUE PLASMA, GLOBULI ROSSI, GLOBULI BIANCHI E PIASTRINE HA FUNZIONI NUTRITIVA E DIFENSIVA.
La Circolazione ALESSIA JULIANO MAX RICCARDO 2^A TRIUGGIO APPARATO CARDIOCIRCOLATORIO CUORE ARTERIE VENE CAPILLARI SANGUE PLASMA, GLOBULI ROSSI, GLOBULI BIANCHI E PIASTRINE HA FUNZIONI NUTRITIVA E DIFENSIVA.
L embrione si sviluppa grazie agli scambi tra sangue materno e fetale. Problema: come fare in modo che sangue materno e fetale restino separati ma
 L embrione si sviluppa grazie agli scambi tra sangue materno e fetale. Problema: come fare in modo che sangue materno e fetale restino separati ma non troppo distanti tra loro? LA PLACENTA E formata dalla
L embrione si sviluppa grazie agli scambi tra sangue materno e fetale. Problema: come fare in modo che sangue materno e fetale restino separati ma non troppo distanti tra loro? LA PLACENTA E formata dalla
I MAMMIFERI Sottoclasse dei Prototeri o Monotremi Sottoclasse dei Metateri o Marsupiali Sottoclasse degli Euteri o Placentati
 I MAMMIFERI Sottoclasse dei Prototeri o Monotremi: le uova si sviluppano al di fuori del corpo materno o all interno ma senza nutrimento fornito dalla madre; Sottoclasse dei Metateri o Marsupiali: embrioni
I MAMMIFERI Sottoclasse dei Prototeri o Monotremi: le uova si sviluppano al di fuori del corpo materno o all interno ma senza nutrimento fornito dalla madre; Sottoclasse dei Metateri o Marsupiali: embrioni
UCCELLI IL IL POLLO COME MODELLO
 IL IL POLLO COME MODELLO Vertebrati amnioti Facile allevamento e deposizione regolare delle uova Grandi dimensioni degli embrioni Sviluppo diretto e relativamente rapido Possibilità di manipolazione chirurgica
IL IL POLLO COME MODELLO Vertebrati amnioti Facile allevamento e deposizione regolare delle uova Grandi dimensioni degli embrioni Sviluppo diretto e relativamente rapido Possibilità di manipolazione chirurgica
L apparato riproduttore femminile
 L apparato riproduttore femminile 2 Ovaie 2 tube di Falloppio Utero Vagina Forma ovale GONADI femminili (OVAIE) All interno dell addome Producono gli ormoni femminili: gli estrogeni e il progesterone permettono
L apparato riproduttore femminile 2 Ovaie 2 tube di Falloppio Utero Vagina Forma ovale GONADI femminili (OVAIE) All interno dell addome Producono gli ormoni femminili: gli estrogeni e il progesterone permettono
SVILUPPO EMBRIONALE (PARTE PRIMA)
") SVILUPPO EMBRIONALE (PARTE PRIMA) Lo sviluppo embrionale è il processo che, per successive mitosi porta dallo zigote all'individuo completo. Lo sviluppo embrionale comprende in realtà vari aspetti, che
SVILUPPO EMBRIONALE (PARTE PRIMA) Lo sviluppo embrionale è il processo che, per successive mitosi porta dallo zigote all'individuo completo. Lo sviluppo embrionale comprende in realtà vari aspetti, che
In base alle caratteristiche delle miofibrille
 Tessuto muscolare Rende possibili sia i movimenti del corpo nell insieme che quelli delle singole parti. Il tessuto muscolare è dotato di contrattilità oltre che di eccitabilità. In base alle caratteristiche
Tessuto muscolare Rende possibili sia i movimenti del corpo nell insieme che quelli delle singole parti. Il tessuto muscolare è dotato di contrattilità oltre che di eccitabilità. In base alle caratteristiche
4^ settimana: eventi fondamentali
 LA QUARTA LA QUARTA 4 mm In ogni phylum di animali c è uno stadio dello sviluppo detto stadio filotipico, in cui gli embrioni di tutte le classi del phylum si assomigliano. La prima fila rappresenta lo
LA QUARTA LA QUARTA 4 mm In ogni phylum di animali c è uno stadio dello sviluppo detto stadio filotipico, in cui gli embrioni di tutte le classi del phylum si assomigliano. La prima fila rappresenta lo
2. Negli Anfibi la circolazione e doppia ma incompleta. Il cuore di una rana ha pertanto:
 Test n. 3 Dalle olimpiadi delle Scienze Naturali 2004 1. L uomo, come tutti i vertebrati, possiede un sistema circolatorio chiuso. Nei mammiferi la circolazione è doppia e completa, poiché il sangue ossigenato
Test n. 3 Dalle olimpiadi delle Scienze Naturali 2004 1. L uomo, come tutti i vertebrati, possiede un sistema circolatorio chiuso. Nei mammiferi la circolazione è doppia e completa, poiché il sangue ossigenato
IL SISTEMA CIRCOLATORIO UMANO
 Funzione Vasi sanguigni Aorta Vene cave IL SISTEMA CIRCOLATORIO Arterie polmonari UMANO Vene polmonari Struttura Circolazione Sistematica e polmonare FUNZIONI Distribuzione delle sostanze nutritive dall
Funzione Vasi sanguigni Aorta Vene cave IL SISTEMA CIRCOLATORIO Arterie polmonari UMANO Vene polmonari Struttura Circolazione Sistematica e polmonare FUNZIONI Distribuzione delle sostanze nutritive dall
I leucociti o globuli bianchi sono cellule coinvolte nella risposta immunitaria. Grazie al loro intervento il corpo umano si difende dagli attacchi
 GLOBULI BIANCHI I leucociti sono cellule del sangue provviste di nucleo e si trovano nel circolo sanguigno, nel sistema linfatico e nei tessuti. La loro caratteristica assenza di pigmentazione gli conferisce
GLOBULI BIANCHI I leucociti sono cellule del sangue provviste di nucleo e si trovano nel circolo sanguigno, nel sistema linfatico e nei tessuti. La loro caratteristica assenza di pigmentazione gli conferisce
La circolazione dei vertebrati
 Il cuore dei vertebrati La circolazione dei vertebrati Nei mammiferi e negli uccelli il sangue venoso è completamente separato dal sangue arterioso. La circolazione viene quindi detta doppia e completa.
Il cuore dei vertebrati La circolazione dei vertebrati Nei mammiferi e negli uccelli il sangue venoso è completamente separato dal sangue arterioso. La circolazione viene quindi detta doppia e completa.
PLACENTA E ANNESSI EMBRIONALI
 Dipartimento di Scienze Ginecologiche, Ostetriche e della Riproduzione PLACENTA E ANNESSI EMBRIONALI Prof. Nicola Colacurci LA PLACENTA Annesso embrionale insieme con: Liquido amniotico Funicolo i l ombelicale
Dipartimento di Scienze Ginecologiche, Ostetriche e della Riproduzione PLACENTA E ANNESSI EMBRIONALI Prof. Nicola Colacurci LA PLACENTA Annesso embrionale insieme con: Liquido amniotico Funicolo i l ombelicale
APPUNTI DI ANATOMIA UMANA
 APPUNTI DI ANATOMIA UMANA L ORGANIZZAZIONE DEL CORPO UMANO ANATOMIA E FISIOLOGIA Anatomia e fisiologia sono due scienze che studiano il corpo umano, in particolare: Anatomia: L anatomia studia la forma
APPUNTI DI ANATOMIA UMANA L ORGANIZZAZIONE DEL CORPO UMANO ANATOMIA E FISIOLOGIA Anatomia e fisiologia sono due scienze che studiano il corpo umano, in particolare: Anatomia: L anatomia studia la forma
Embriologia del pollo
 Embriologia del pollo Anatomia funzionale degli uccelli domestici Virgilio BOTTE - Gaetano PELAGALLI Edi-Ermes, Milano, 1982 Il frequente uso delle uova embrionate di uccelli per indagini microbiologiche
Embriologia del pollo Anatomia funzionale degli uccelli domestici Virgilio BOTTE - Gaetano PELAGALLI Edi-Ermes, Milano, 1982 Il frequente uso delle uova embrionate di uccelli per indagini microbiologiche
Funzioni principali. L apparato circolatorio Prof.ssa Silvia Recchia. Funzioni secondarie. Come è fatto...
 Funzioni principali L apparato circolatorio Prof.ssa Silvia Recchia L apparato circolatorio è deputato al trasporto di nutrienti e ossigeno a tutti gli organi e tessuti di un organismo insieme al sistema
Funzioni principali L apparato circolatorio Prof.ssa Silvia Recchia L apparato circolatorio è deputato al trasporto di nutrienti e ossigeno a tutti gli organi e tessuti di un organismo insieme al sistema
Cellule tessuti e corpo in generale
 Cellule tessuti e corpo in generale Il corpo umano è una macchina straordinaria e meravigliosa. È fatta di oltre 200 ossa, 450 muscoli, 100.000 capelli in testa, 130.000.000 di recettori sul fondo della
Cellule tessuti e corpo in generale Il corpo umano è una macchina straordinaria e meravigliosa. È fatta di oltre 200 ossa, 450 muscoli, 100.000 capelli in testa, 130.000.000 di recettori sul fondo della
Apparato circolatorio
 Apparato circolatorio COME SIAMO FATTI? Il nostro corpo è composto da cellule. Più cellule con la stessa funzione formano un tessuto. Un insieme di diversi tessuti organizzati in una struttura unica con
Apparato circolatorio COME SIAMO FATTI? Il nostro corpo è composto da cellule. Più cellule con la stessa funzione formano un tessuto. Un insieme di diversi tessuti organizzati in una struttura unica con
Tema C Capitolo 10 L apparato riproduttore
 1. Le cellule per la riproduzione 2. La mitosi e la meiosi 3. L apparato riproduttore maschile 4. L apparato riproduttore femminile 5. Il ciclo riproduttivo femminile 6. Come si forma una nuova vita 7.
1. Le cellule per la riproduzione 2. La mitosi e la meiosi 3. L apparato riproduttore maschile 4. L apparato riproduttore femminile 5. Il ciclo riproduttivo femminile 6. Come si forma una nuova vita 7.
L apparato digerente. La digestione inizia in bocca.
 L apparato digerente Il corpo umano è costituito principalmente da acqua, proteine, grassi, zuccheri, Sali minerali e vitamine. Durante le sue attività l organismo consuma queste sostanze che perciò devono
L apparato digerente Il corpo umano è costituito principalmente da acqua, proteine, grassi, zuccheri, Sali minerali e vitamine. Durante le sue attività l organismo consuma queste sostanze che perciò devono
La Riproduzione e l Apparato Riproduttivo Umano
 La Riproduzione e l Apparato Riproduttivo Umano La Riproduzione Perché gli esseri viventi si riproducono? La riproduzione (o procreazione) è il processo attraverso il quale gli esseri viventi fanno sì
La Riproduzione e l Apparato Riproduttivo Umano La Riproduzione Perché gli esseri viventi si riproducono? La riproduzione (o procreazione) è il processo attraverso il quale gli esseri viventi fanno sì
una cavità corporea, nel lume di un organo oppure sulla superficie esterna del
 Il sistema endocrino è costituito da diverse ghiandole endocrine e da molte cellule secretrici di ormoni, localizzate in organi che hanno anche altre funzioni. Vi sono due tipi di ghiandole le ghiandole
Il sistema endocrino è costituito da diverse ghiandole endocrine e da molte cellule secretrici di ormoni, localizzate in organi che hanno anche altre funzioni. Vi sono due tipi di ghiandole le ghiandole
ANATOMIA MICROSPICA APPARATO DIGERENTE 1
 ANATOMIA MICROSPICA APPARATO DIGERENTE 1 -Metodo di studio -Cenni canale alimentare -Labbra -Dente -Lingua -Esofago -Cardias -Stomaco -Piloro -Intestino Tenue -Intestino Crasso -Appendice -Cenni Peritoneo
ANATOMIA MICROSPICA APPARATO DIGERENTE 1 -Metodo di studio -Cenni canale alimentare -Labbra -Dente -Lingua -Esofago -Cardias -Stomaco -Piloro -Intestino Tenue -Intestino Crasso -Appendice -Cenni Peritoneo
IL LAVORO DEI CITTADINI STRANIERI
 IL LAVORO DEI CITTADINI STRANIERI Roma e provincia nel 2014 Indice I numeri più significativi... 2 Le conseguenze della Crisi economica... 3 Il contesto nazionale... 3 I numeri di Roma: la condizione occupazionale...
IL LAVORO DEI CITTADINI STRANIERI Roma e provincia nel 2014 Indice I numeri più significativi... 2 Le conseguenze della Crisi economica... 3 Il contesto nazionale... 3 I numeri di Roma: la condizione occupazionale...
Gli animali che abitano il mare. Guarda le immagini e dopo rispondi alle domande..
 Gli animali che abitano il mare Guarda le immagini e dopo rispondi alle domande.. 1 2 3 4 5 1 1) Hai mai visto questi animali? SÌ NO QUALCUNO 2) Se li hai visti, dove? ALLA TELEVISIONE SUL TUO LIBRO DI
Gli animali che abitano il mare Guarda le immagini e dopo rispondi alle domande.. 1 2 3 4 5 1 1) Hai mai visto questi animali? SÌ NO QUALCUNO 2) Se li hai visti, dove? ALLA TELEVISIONE SUL TUO LIBRO DI
SOLUZIONI. Soluzione diluita: è costituita da un liquido (solvente) in cui è disciolta una piccola quantità di un altra sostanza (soluto).
 in cui è disciolta una piccola quantità di un altra sostanza (soluto).") SOLUZIONI Soluzione diluita: è costituita da un liquido (solvente) in cui è disciolta una piccola quantità di un altra sostanza (soluto). Concentrazione di una soluzione: Molarità: Frazione molare: 1 SOLUZIONI
SOLUZIONI Soluzione diluita: è costituita da un liquido (solvente) in cui è disciolta una piccola quantità di un altra sostanza (soluto). Concentrazione di una soluzione: Molarità: Frazione molare: 1 SOLUZIONI
GHIANDOLE ANNESSE ALL APPARATO APPARATO DIGERENTE. Fegato, Pancreas, Cistifellea
 GHIANDOLE ANNESSE ALL APPARATO APPARATO DIGERENTE Fegato, Pancreas, Cistifellea Gli organi Fegato. Secerne la bile (emulsionante dei grassi) e proteine del sangue (fibrinogeno, albumine) Metabolizzazione
GHIANDOLE ANNESSE ALL APPARATO APPARATO DIGERENTE Fegato, Pancreas, Cistifellea Gli organi Fegato. Secerne la bile (emulsionante dei grassi) e proteine del sangue (fibrinogeno, albumine) Metabolizzazione
MEMBRANA IN COLLAGENE NATIVO AD ALTA RESISTENZA BIO-MECCANICA. Fogli per la sostituzione/riparazione tissutale possono essere costituiti da:
 MEMBRANA IN COLLAGENE NATIVO AD ALTA RESISTENZA BIO-MECCANICA Fogli per la sostituzione/riparazione tissutale possono essere costituiti da: - reti di polimeri bioinerti non riassorbibili (i quali danno
MEMBRANA IN COLLAGENE NATIVO AD ALTA RESISTENZA BIO-MECCANICA Fogli per la sostituzione/riparazione tissutale possono essere costituiti da: - reti di polimeri bioinerti non riassorbibili (i quali danno
Introduzione alla Biologia della Cellula. Lezione 1 L unità base degli organismi viventi: la cellula
 Introduzione alla Biologia della Cellula Lezione 1 L unità base degli organismi viventi: la cellula Teoria cellulare TUTTI gli organismi sono composti da una o più CELLULE (unicellulari/pluricellulari)
Introduzione alla Biologia della Cellula Lezione 1 L unità base degli organismi viventi: la cellula Teoria cellulare TUTTI gli organismi sono composti da una o più CELLULE (unicellulari/pluricellulari)
ANAT ANA O T MIA G ENERALE GENERALE U MANA UMANA
 CORSO DI FORMAZIONE TECNICO EDUCATORE REGIONALE LA MACCHINA DELL UOMO: CENNI DI FISIOLOGIA, ANATOMIA DELL APPARATO LOCOMOTORE TERMINOLOGIA ANATOMICA Relatore: Dott.Michele Bisogni Medico chirurgo Specializzando
CORSO DI FORMAZIONE TECNICO EDUCATORE REGIONALE LA MACCHINA DELL UOMO: CENNI DI FISIOLOGIA, ANATOMIA DELL APPARATO LOCOMOTORE TERMINOLOGIA ANATOMICA Relatore: Dott.Michele Bisogni Medico chirurgo Specializzando
METRORRAGIE NEL III TRIMESTRE DI GRAVIDANZA
 Università degli Studi di Ferrara Prof. Pantaleo Greco METRORRAGIE NEL III TRIMESTRE DI GRAVIDANZA Placenta previa Distacco intempestivo di placenta Placenta previa La placenta si definisce previa quando
Università degli Studi di Ferrara Prof. Pantaleo Greco METRORRAGIE NEL III TRIMESTRE DI GRAVIDANZA Placenta previa Distacco intempestivo di placenta Placenta previa La placenta si definisce previa quando
APPARATO RESPIRATORIO: quali sono le principali funzioni?
 APPARATO RESPIRATORIO: quali sono le principali funzioni? Funzioni dell apparato respiratorio 1. Fa entrare ed uscire l aria nel corpo (ventilazione) 2. Fornisce un ampia superficie di scambio tra aria
APPARATO RESPIRATORIO: quali sono le principali funzioni? Funzioni dell apparato respiratorio 1. Fa entrare ed uscire l aria nel corpo (ventilazione) 2. Fornisce un ampia superficie di scambio tra aria
Vertebr ati. Vertebr. ati. Rettili
 Vertebr Vertebr ati ati Pesc Pesc ii Anfib Anfib ii Rettili Rettili Scheletr Scheletr oo interno interno Uccell Uccell ii Mammife Mammife ri ri Pesci I pesci sono vertebrati a sangue freddo vale a dire
Vertebr Vertebr ati ati Pesc Pesc ii Anfib Anfib ii Rettili Rettili Scheletr Scheletr oo interno interno Uccell Uccell ii Mammife Mammife ri ri Pesci I pesci sono vertebrati a sangue freddo vale a dire
LA NEVE E LE VALANGHE
 3^LEZIONE INTERVENTO METEOROLOGO LUCA LOMBROSO Il ciclo dell acqua LA NEVE E LE VALANGHE Prima di capire cos è la neve è opportuno approfondire la conoscenza del liquido più comune sulla Terra: l acqua
3^LEZIONE INTERVENTO METEOROLOGO LUCA LOMBROSO Il ciclo dell acqua LA NEVE E LE VALANGHE Prima di capire cos è la neve è opportuno approfondire la conoscenza del liquido più comune sulla Terra: l acqua
MIDOLLO SPINALE Nervi spinali e gangli sensitivi
 MIDOLLO SPINALE Nervi spinali e gangli sensitivi Encefalo CENTRALE (SNC) SISTEMA NERVOSO PERIFERICO (SNP) Nervi Midollo spinale Encefalo (contenuto nella scatola cranica) Midollo spinale (contenuto nel
MIDOLLO SPINALE Nervi spinali e gangli sensitivi Encefalo CENTRALE (SNC) SISTEMA NERVOSO PERIFERICO (SNP) Nervi Midollo spinale Encefalo (contenuto nella scatola cranica) Midollo spinale (contenuto nel
Capitolo 1 Strutture e funzioni degli animali: i concetti unificanti
 Capitolo 1 Strutture e funzioni degli animali: i concetti unificanti ANATOMIA : FISIOLOGIA : struttura di un organismo; funzioni delle varie parti di un organismo; Arto anteriore Polso Primo dito Rachide
Capitolo 1 Strutture e funzioni degli animali: i concetti unificanti ANATOMIA : FISIOLOGIA : struttura di un organismo; funzioni delle varie parti di un organismo; Arto anteriore Polso Primo dito Rachide
Il legno legno scorza cilindro centrale corteccia epidermide libro legno cambio
 Il legno Comunemente si indica come legno la parte del fusto degli alberi che si trova sotto la cosiddetta scorza (comunemente detta corteccia). Nel 1 anno di vita del fusto si distingue una zona a funzione
Il legno Comunemente si indica come legno la parte del fusto degli alberi che si trova sotto la cosiddetta scorza (comunemente detta corteccia). Nel 1 anno di vita del fusto si distingue una zona a funzione
Inquinamento da elettromagnetismo. Cos'è Perché è importante Come agire
 Inquinamento da elettromagnetismo Cos'è Perché è importante Come agire Inquinamento da elettromagnetismo cos'è DEFINIZIONE: È una forma particolare di inquinamento ambientale, non si ha una vera immissione
Inquinamento da elettromagnetismo Cos'è Perché è importante Come agire Inquinamento da elettromagnetismo cos'è DEFINIZIONE: È una forma particolare di inquinamento ambientale, non si ha una vera immissione
OVOGENESI E CICLI FEMMINILI
 OVOGENESI E CICLI FEMMINILI Anatomia del sistema riproduttivo femminile. A destra, sezione di ovaio nella zona corticale (ingrandimento del riquadro rosso). E, epitelio di rivestimento dell ovaio; TA,
OVOGENESI E CICLI FEMMINILI Anatomia del sistema riproduttivo femminile. A destra, sezione di ovaio nella zona corticale (ingrandimento del riquadro rosso). E, epitelio di rivestimento dell ovaio; TA,
CENNI DI FISIOLOGIA UMANA
 CENNI DI FISIOLOGIA UMANA o L'APPARATO CARDIOCIRCOLATORIO o L'APPARATO RESPIRATORIO o L'APPARATO OSTEOARTICOLARE cuore cervello polmoni scheletro 1 La Fisiologia è la scienza che studia la funzionalità
CENNI DI FISIOLOGIA UMANA o L'APPARATO CARDIOCIRCOLATORIO o L'APPARATO RESPIRATORIO o L'APPARATO OSTEOARTICOLARE cuore cervello polmoni scheletro 1 La Fisiologia è la scienza che studia la funzionalità
Epiteli ghiandolari. origine embriologica. Ghiandole esocrine endocrine. gli elementi secernenti (adenomero* o. epiteliali
 Ghiandole (esocrine ed endocrine) Cellule singole o organi specializzati nella produzione e secrezione di sostanze che svolgono una varietà di funzioni biologiche nell organismo Created by G. Papaccio
Ghiandole (esocrine ed endocrine) Cellule singole o organi specializzati nella produzione e secrezione di sostanze che svolgono una varietà di funzioni biologiche nell organismo Created by G. Papaccio
Gravidanza o no? Patient Diagnostics Test Rapidi della Progettazione Familiare
 Gravidanza o no? Patient Diagnostics Test Rapidi della Progettazione Familiare Gli unici test a domicilio per la pianificazione familiare Test di ovulazione Test precoce di gravidanza Test di gravidanza
Gravidanza o no? Patient Diagnostics Test Rapidi della Progettazione Familiare Gli unici test a domicilio per la pianificazione familiare Test di ovulazione Test precoce di gravidanza Test di gravidanza
- lipidi strutturali, costituenti fondamentali delle membrane cellulari ed intracellulari (fosfolipidi, glicolipidi e colesterolo)
") I lipidi, o grassi, costituiscono un gruppo eterogeneo di sostanze accomunate dalla proprietà fisica della insolubilità nei solventi polari (es. acqua) (idrofobicità) e dalla solubilità nei soventi organici
I lipidi, o grassi, costituiscono un gruppo eterogeneo di sostanze accomunate dalla proprietà fisica della insolubilità nei solventi polari (es. acqua) (idrofobicità) e dalla solubilità nei soventi organici
Tempi transito infundibulo minuti: Magnum ore: 3h istmi ore: 1h:15 Utero ore: 21h vagina minuti: 1-2
 Pollo003 L apparato riproduttore della gallina matura sessualmente è in cavità addominale. Ovaio: fra rene sinistro, polmone sinistro ed appoggia sul sacco aereo addominale sinistro. È sostenuto sotto
Pollo003 L apparato riproduttore della gallina matura sessualmente è in cavità addominale. Ovaio: fra rene sinistro, polmone sinistro ed appoggia sul sacco aereo addominale sinistro. È sostenuto sotto
Alimentazione ed efficienza dei sistemi alimentari
 Corso di laurea magistrale in Scienze e tecnologie animali Alimentazione ed efficienza dei sistemi alimentari Effetti delle tipologie dei consumi alimentari sui flussi della materia e dell energia, sulle
Corso di laurea magistrale in Scienze e tecnologie animali Alimentazione ed efficienza dei sistemi alimentari Effetti delle tipologie dei consumi alimentari sui flussi della materia e dell energia, sulle
Trasporto di O 2 nel sangue
 Trasporto di O 2 nel sangue Il 97% dell O 2 trasportato nel plasma è chimicamente legato all Hb nei globuli rossi, solo il 3% è fisicamente disciolto Trasporto O 2 nel plasma Trasporto O 2 legato ad Hb
Trasporto di O 2 nel sangue Il 97% dell O 2 trasportato nel plasma è chimicamente legato all Hb nei globuli rossi, solo il 3% è fisicamente disciolto Trasporto O 2 nel plasma Trasporto O 2 legato ad Hb
Il DNA, insieme a diverse proteine si organizza in una
 MITOSI E MEIOSI Il DNA, insieme a diverse proteine si organizza in una struttura che è detta CROMOSOMA. I cromosomi sono costituiti da cromatina, che consiste di fibre contenenti DNA e proteine. Quando
MITOSI E MEIOSI Il DNA, insieme a diverse proteine si organizza in una struttura che è detta CROMOSOMA. I cromosomi sono costituiti da cromatina, che consiste di fibre contenenti DNA e proteine. Quando
L APPARATO CIRCOLATORIO
 L APPARATO CIRCOLATORIO DA COSA È COMPOSTO L apparato circolatorio è costituito dal sangue (che provvede al nutrimento di tutte le cellule del corpo)la cui circolazione si compie nelle arterie, nelle vene
L APPARATO CIRCOLATORIO DA COSA È COMPOSTO L apparato circolatorio è costituito dal sangue (che provvede al nutrimento di tutte le cellule del corpo)la cui circolazione si compie nelle arterie, nelle vene
ANATOMIA E FISIO-PATOLOGIA LE POSIZIONI ANATOMICHE, LE CAVITA E LE PARTI DEL CORPO
 ANATOMIA E FISIO-PATOLOGIA LE POSIZIONI ANATOMICHE, LE CAVITA E LE PARTI DEL CORPO CORSO CORSO OSS DISCIPLINA: OSS DISCIPLINA: ANATOMIA PRIMO E FISIO-PATOLOGIA SOCCORSO MASSIMO MASSIMO FRANZIN FRANZIN
ANATOMIA E FISIO-PATOLOGIA LE POSIZIONI ANATOMICHE, LE CAVITA E LE PARTI DEL CORPO CORSO CORSO OSS DISCIPLINA: OSS DISCIPLINA: ANATOMIA PRIMO E FISIO-PATOLOGIA SOCCORSO MASSIMO MASSIMO FRANZIN FRANZIN
La riproduzione nelle piante. Lezioni d'autore
 La riproduzione nelle piante Lezioni d'autore VIDEO Introduzione (I) Come in molti altri esseri viventi, anche nelle piante la riproduzione può essere sessuata o asessuata. Esistono specie che presentano
La riproduzione nelle piante Lezioni d'autore VIDEO Introduzione (I) Come in molti altri esseri viventi, anche nelle piante la riproduzione può essere sessuata o asessuata. Esistono specie che presentano
Impianti di propulsione navale
 Motori diesel 2T Descrizione dei motori ME e RT-Flex Le caratteristiche ed i vantaggi dei motori a controllo completamente elettronico sono già state elencate i precedenza. In questo capitolo saranno illustrati
Motori diesel 2T Descrizione dei motori ME e RT-Flex Le caratteristiche ed i vantaggi dei motori a controllo completamente elettronico sono già state elencate i precedenza. In questo capitolo saranno illustrati
La spermatogenesi. Processo differenziativo degli spz. A partire dagli spermatogoni
 La spermatogenesi Processo differenziativo degli spz A partire dagli spermatogoni La spermatogenesi Sede della spermatogenesi Nel testicolo Tre compartimenti funzionali Interstiziale (vasi, cell. Leydig,
La spermatogenesi Processo differenziativo degli spz A partire dagli spermatogoni La spermatogenesi Sede della spermatogenesi Nel testicolo Tre compartimenti funzionali Interstiziale (vasi, cell. Leydig,
europlus-silentium EFFETTO SULLA TRASMISSIONE DEL RUMORE AL CALPESTIO PROGETTO DI RICERCA ALLESTIMENTO IN LABORATORIO
 EFFETTO SULLA TRASMISSIONE DEL RUMORE AL CALPESTIO Misure di rumore di calpestio su solaio laterocementizio in laboratorio secondo UNI EN ISO 140-6 Nel mese di maggio 2009 è iniziata la sperimentazione
EFFETTO SULLA TRASMISSIONE DEL RUMORE AL CALPESTIO Misure di rumore di calpestio su solaio laterocementizio in laboratorio secondo UNI EN ISO 140-6 Nel mese di maggio 2009 è iniziata la sperimentazione
SC.BIOLOGICHE I Anno - CORSO A LABORATORIO CITOLOGIA-ISTOLOGIA. Esercitazione n 4 CONNETTIVI DI SOSTEGNO
 SC.BIOLOGICHE I Anno - CORSO A LABORATORIO CITOLOGIA-ISTOLOGIA Esercitazione n 4 CONNETTIVI DI SOSTEGNO Epiteliale Di Di rivestimento rivestimento Sensoriale Sensoriale Ghiandolare Ghiandolare Connettivo
SC.BIOLOGICHE I Anno - CORSO A LABORATORIO CITOLOGIA-ISTOLOGIA Esercitazione n 4 CONNETTIVI DI SOSTEGNO Epiteliale Di Di rivestimento rivestimento Sensoriale Sensoriale Ghiandolare Ghiandolare Connettivo
Chimica. Lezione 2 Il legame chimico Parte I
 Chimica Lezione 2 Il legame chimico Parte I GLI ATOMI TENDONO A LEGARSI SPONTANEAMENTE FRA DI LORO, PER FORMARE DELLE MOLECOLE, OGNI QUALVOLTA QUESTO PROCESSO PERMETTE LORO DI RAGGIUNGERE UNA CONDIZIONE
Chimica Lezione 2 Il legame chimico Parte I GLI ATOMI TENDONO A LEGARSI SPONTANEAMENTE FRA DI LORO, PER FORMARE DELLE MOLECOLE, OGNI QUALVOLTA QUESTO PROCESSO PERMETTE LORO DI RAGGIUNGERE UNA CONDIZIONE
Le proprietà colligative delle soluzioni Sappiamo dall esperienza quotidiana che una soluzione ha caratteristiche diverse dal solvente puro.
 Le proprietà colligative delle soluzioni Sappiamo dall esperienza quotidiana che una soluzione ha caratteristiche diverse dal solvente puro. Molte delle sue proprietà, come il sapore, il colore, l odore,
Le proprietà colligative delle soluzioni Sappiamo dall esperienza quotidiana che una soluzione ha caratteristiche diverse dal solvente puro. Molte delle sue proprietà, come il sapore, il colore, l odore,
ANATOMIA ECOGRAFICA PELVI FEMMINILE. A. Stimamiglio
 ANATOMIA ECOGRAFICA PELVI FEMMINILE A. Stimamiglio ECOGRAFIA UTERO Evidenziazione del legamento largo in caso di ascite MISURE UTERINE NELLE DIVERSE ETA' longitudinale AP Trasverso Volume cm cm cm ml
ANATOMIA ECOGRAFICA PELVI FEMMINILE A. Stimamiglio ECOGRAFIA UTERO Evidenziazione del legamento largo in caso di ascite MISURE UTERINE NELLE DIVERSE ETA' longitudinale AP Trasverso Volume cm cm cm ml
Test di depistaggio prenatale del 1 trimestre Informativa per lo studio medico
 Test di depistaggio prenatale del 1 trimestre Informativa per lo studio medico Determinazione del rischio di trisomie fetali attraverso l età materna, la translucenza nucale e i valori di PAPP-A e free
Test di depistaggio prenatale del 1 trimestre Informativa per lo studio medico Determinazione del rischio di trisomie fetali attraverso l età materna, la translucenza nucale e i valori di PAPP-A e free
ATOMI E MOLECOLE. Tutte le varie forme di materia esistenti sono costituite da sostanze semplici (elementi) e da sostanze composte (composti).
 e da sostanze composte (composti).") 1 ATOMI E MOLECOLE Tutte le varie forme di materia esistenti sono costituite da sostanze semplici (elementi) e da sostanze composte (composti). Un elemento (es. il mercurio) è una sostanza che non può
1 ATOMI E MOLECOLE Tutte le varie forme di materia esistenti sono costituite da sostanze semplici (elementi) e da sostanze composte (composti). Un elemento (es. il mercurio) è una sostanza che non può
Rapporti tra malnutrizione e salute
 La nutrizione Rapporti tra malnutrizione e salute La nutrizione è uno dei più importanti determinanti della salute. Esiste una stretta correlazione tra alimentazione non corretta e insorgenza di numerose
La nutrizione Rapporti tra malnutrizione e salute La nutrizione è uno dei più importanti determinanti della salute. Esiste una stretta correlazione tra alimentazione non corretta e insorgenza di numerose
Osmometria ed Emolisi
 Osmometria ed Emolisi 1 ADE: MISURA DELLA RESISTENZA OSMOTICA DEI GLOBULI ROSSI (2/12/08). Il globulo rosso è una piccola cellula del sangue deputata al trasporto dell'ossigeno e priva di nucleo e di organelli.
Osmometria ed Emolisi 1 ADE: MISURA DELLA RESISTENZA OSMOTICA DEI GLOBULI ROSSI (2/12/08). Il globulo rosso è una piccola cellula del sangue deputata al trasporto dell'ossigeno e priva di nucleo e di organelli.
È un tubo lungo cm 10, diametro 2-2,5 cm, in parte cartilagineo in parte membranoso, gli anelli cartilaginei che la formano sono incompleti
 Trachea È costituita da 2 porzioni, cervicale e toracica Si sviluppa dal margine inferiore della cartilagine cricoidea (C6) fino a T4 dove si biforca in due bronchi primari, che divergono ed entrano nei
Trachea È costituita da 2 porzioni, cervicale e toracica Si sviluppa dal margine inferiore della cartilagine cricoidea (C6) fino a T4 dove si biforca in due bronchi primari, che divergono ed entrano nei
APPARATO GENITALE maschile
 APPARATO GENITALE maschile Proliferazione dell epitelio celomatico Parziale degenerazione del mesonefro Gli elementi epiteliali vanno a formare a formare i cordoni sessuali primitivi Gonadi Vie genitali
APPARATO GENITALE maschile Proliferazione dell epitelio celomatico Parziale degenerazione del mesonefro Gli elementi epiteliali vanno a formare a formare i cordoni sessuali primitivi Gonadi Vie genitali
www.slidetube.it Il nefrone è costituito dal corpuscolo renale e dal tubulo
 Il nefrone è costituito dal corpuscolo renale e dal tubulo Il corpuscolo renale è formato da : a) il glomerulo arterioso b) la capsula di Bowman www.slidetube.it Zona corticale Zona midollare Dall arteria
Il nefrone è costituito dal corpuscolo renale e dal tubulo Il corpuscolo renale è formato da : a) il glomerulo arterioso b) la capsula di Bowman www.slidetube.it Zona corticale Zona midollare Dall arteria
Lo sviluppo embrionale.
 Lo sviluppo embrionale. La riproduzione umana In biologia si definisce riproduzione il processo attraverso il quale vengono generati nuovi individui della stessa specie. La riproduzione umana è caratterizzata
Lo sviluppo embrionale. La riproduzione umana In biologia si definisce riproduzione il processo attraverso il quale vengono generati nuovi individui della stessa specie. La riproduzione umana è caratterizzata
Epitelio di origine endodermica (tranne i tratti ectodermici iniziali e terminali del proctodeo e dello stomodeo)
") L APPARATO DIGERENTE Tubo digerente (dalla bocca all ano) e ghiandole annesse; epitelio di origine endodermica (tranne i tratti ectodermici iniziali e terminali del proctodeo e dello stomodeo) Epitelio
L APPARATO DIGERENTE Tubo digerente (dalla bocca all ano) e ghiandole annesse; epitelio di origine endodermica (tranne i tratti ectodermici iniziali e terminali del proctodeo e dello stomodeo) Epitelio
Ruolo del calcio nella cellula
 Ruolo del calcio nella cellula I meccanismi evolutivi hanno attribuito agli ioni fosfato un importanza fondamentale poiché sono necessari per sintetizzare ATP. Per questo motivo la loro concentrazione
Ruolo del calcio nella cellula I meccanismi evolutivi hanno attribuito agli ioni fosfato un importanza fondamentale poiché sono necessari per sintetizzare ATP. Per questo motivo la loro concentrazione
PERCHÉ RESPIRIAMO PROF.SSA PORTAS NERINA ISTITUTO AMSICORA I.P.I.A. OLBIA
 PERCHÉ RESPIRIAMO Respiriamo per portare ossigeno alle cellule dove avviene la respirazione cellulare. La respirazione cellulare è il meccanismo attraverso cui la cellula, in presenza di ossigeno, è in
PERCHÉ RESPIRIAMO Respiriamo per portare ossigeno alle cellule dove avviene la respirazione cellulare. La respirazione cellulare è il meccanismo attraverso cui la cellula, in presenza di ossigeno, è in
FUSIONE MATERIALE GENETICO APLOIDE 300 MIO 200 ovocita 12-24 ore dopo l ovulazione Capacitazione Completamento Meiosi II La cellula uovo, ancora
 FECONDAZIONE FUSIONE MATERIALE GENETICO APLOIDE 300 MIO 200 ovocita 12-24 ore dopo l ovulazione Capacitazione Completamento Meiosi II La cellula uovo, ancora circondata dalle più piccole cellule follicolari
FECONDAZIONE FUSIONE MATERIALE GENETICO APLOIDE 300 MIO 200 ovocita 12-24 ore dopo l ovulazione Capacitazione Completamento Meiosi II La cellula uovo, ancora circondata dalle più piccole cellule follicolari
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da
. La stabilità del ph è insidiata da") Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni
 5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
Caratteristiche fisiologiche del neonato
 Caratteristiche fisiologiche del neonato CARATTERISTICHE FISIOLOGICHE DEL NEONATO I primi due giorni di vita: è Bellissimo!! Unico, Perfetto!!! CARATTERISTICHE FISIOLOGICHE DEL NEONATO Dopo due giorni:
Caratteristiche fisiologiche del neonato CARATTERISTICHE FISIOLOGICHE DEL NEONATO I primi due giorni di vita: è Bellissimo!! Unico, Perfetto!!! CARATTERISTICHE FISIOLOGICHE DEL NEONATO Dopo due giorni:
Apparato scheletrico
 Apparato scheletrico L'apparato scheletrico ha la funzione di sostegno e di protezione degli organi interni ed è costituita dalle ossa. Le ossa sono formate dal tessuto osseo, composto da una sostanza
Apparato scheletrico L'apparato scheletrico ha la funzione di sostegno e di protezione degli organi interni ed è costituita dalle ossa. Le ossa sono formate dal tessuto osseo, composto da una sostanza
Lo stato liquido. Un liquido non ha una forma propria, ma ha la forma del recipiente che lo contiene; ha però volume proprio e non è comprimibile.
 I liquidi Lo stato liquido Lo stato liquido rappresenta una condizione intermedia tra stato aeriforme e stato solido, tra lo stato di massimo disordine e quello di perfetto ordine Un liquido non ha una
I liquidi Lo stato liquido Lo stato liquido rappresenta una condizione intermedia tra stato aeriforme e stato solido, tra lo stato di massimo disordine e quello di perfetto ordine Un liquido non ha una
E caratterizzata da 2 processi fondamentali:
 LA QUARTA SETTIMANA EVENTI IV a SETTIMANA E caratterizzata da 2 processi fondamentali: 1) la crescita e l organizzazione dei derivati dei tre foglietti embrionali, che portano alla formazione degli abbozzi
LA QUARTA SETTIMANA EVENTI IV a SETTIMANA E caratterizzata da 2 processi fondamentali: 1) la crescita e l organizzazione dei derivati dei tre foglietti embrionali, che portano alla formazione degli abbozzi
gonadi, gonodotti ed eventuali organi copulatori
 L apparato genitale comprende: gonadi, gonodotti ed eventuali organi copulatori Le gonadi maschili, testicoli e quelle femminili, ovari sono il luogo di produzione dei gameti, spernmatozoi o ovociti e
L apparato genitale comprende: gonadi, gonodotti ed eventuali organi copulatori Le gonadi maschili, testicoli e quelle femminili, ovari sono il luogo di produzione dei gameti, spernmatozoi o ovociti e
2) Tessuto Connettivo t. c. propriamente detti t. Cartilagineo t. Osseo Sangue. 3) Tessuto muscolare striato scheletrico cardiaco liscio
 Tessuto Connettivo t. c. propriamente detti t. Cartilagineo t. Osseo Sangue. 3) Tessuto muscolare striato scheletrico cardiaco liscio") La cellula è la più piccola unità di sostanza vivente di un organismo pluricellulare. E delimitata dalla membrana plasmatica e contiene il citoplasma, gli organuli e gli inclusi citoplasmatici. Anche se
La cellula è la più piccola unità di sostanza vivente di un organismo pluricellulare. E delimitata dalla membrana plasmatica e contiene il citoplasma, gli organuli e gli inclusi citoplasmatici. Anche se
4 Richiesta di radiografie - osservazione
 A questo punto del percorso abbiamo rilevato che il corpo umano affascina e incuriosisce i bambini anche se la comprensione di questa problematica risulta complessa, per alunni della scuola elementare,
A questo punto del percorso abbiamo rilevato che il corpo umano affascina e incuriosisce i bambini anche se la comprensione di questa problematica risulta complessa, per alunni della scuola elementare,
I LIPIDI (GRASSI) Cosa sono
 Cosa sono") I LIPIDI (GRASSI) Cosa sono Comunemente chiamati "grassi", i lipidi comprendono una grande varietà di molecole, accomunate dalla caratteristica di essere insolubili in acqua. I lipidi più importanti dal
I LIPIDI (GRASSI) Cosa sono Comunemente chiamati "grassi", i lipidi comprendono una grande varietà di molecole, accomunate dalla caratteristica di essere insolubili in acqua. I lipidi più importanti dal
Eucarioti e Procarioti. Prof.ssa Ilaria Godoli
 Eucarioti e Procarioti Prof.ssa Ilaria Godoli Per studiare i viventi......nell'antichità studiare le caratteristiche di esseri viventi molto piccoli è stato pressoché impossibile perché NON c'erano gli
Eucarioti e Procarioti Prof.ssa Ilaria Godoli Per studiare i viventi......nell'antichità studiare le caratteristiche di esseri viventi molto piccoli è stato pressoché impossibile perché NON c'erano gli
Fabio Fichera Italo Paolini Area Ecografica SIMG
 Fabio Fichera Italo Paolini Area Ecografica SIMG Bolzano, 11 13 Febbraio 2016 DIAMETRO TRASVERSO: 3-4 CM. DIAMETRO A-P: 2-3 CM. DIAMETRO LONGITUDINALE: >3,5 CM. NORMALE: UNIFORMEMENTE IPOECOGENA L
Fabio Fichera Italo Paolini Area Ecografica SIMG Bolzano, 11 13 Febbraio 2016 DIAMETRO TRASVERSO: 3-4 CM. DIAMETRO A-P: 2-3 CM. DIAMETRO LONGITUDINALE: >3,5 CM. NORMALE: UNIFORMEMENTE IPOECOGENA L
Il sistema di sigillatura "ISOLBLOCK" nelle murature refrattarie di sicurezza
 Il sistema di sigillatura "ISOLBLOCK" nelle murature refrattarie di sicurezza La società REFRASUD, nel campo delle esperienze maturate sui rivestimenti per siviere acciaio, ha formulato una particolare
Il sistema di sigillatura "ISOLBLOCK" nelle murature refrattarie di sicurezza La società REFRASUD, nel campo delle esperienze maturate sui rivestimenti per siviere acciaio, ha formulato una particolare
LE PARTI DELLA PIANTA
 LE PARTI DELLA PIANTA Esistono vari tipi di vegetali, ma quasi tutti sono formati da tre parti fondamentali: FOGLIE Le radici FUSTO Il fusto Le foglie RADICI Le radici hanno tre compiti principali: 1.
LE PARTI DELLA PIANTA Esistono vari tipi di vegetali, ma quasi tutti sono formati da tre parti fondamentali: FOGLIE Le radici FUSTO Il fusto Le foglie RADICI Le radici hanno tre compiti principali: 1.
Esempi di attribuzione dei seggi
 Esempi di attribuzione dei seggi Al fine di chiarire il funzionamento dei meccanismi previsti per l attribuzione dei seggi e l ordine delle relative operazioni, vengono presentati due esempi di attribuzione
Esempi di attribuzione dei seggi Al fine di chiarire il funzionamento dei meccanismi previsti per l attribuzione dei seggi e l ordine delle relative operazioni, vengono presentati due esempi di attribuzione
Arintha biotech Dicembre 2005
 GENETICA MOLECOLARE Arintha biotech Dicembre 2005 GENETICA MOLECOLARE Il genoma è costituito dall insieme dei geni, localizzati all interno dei cromosomi Ogni gene codifica una proteina. Ogni proteina
GENETICA MOLECOLARE Arintha biotech Dicembre 2005 GENETICA MOLECOLARE Il genoma è costituito dall insieme dei geni, localizzati all interno dei cromosomi Ogni gene codifica una proteina. Ogni proteina
Gli alimenti e la piramide alimentare
 Ausilioteca del Comune di Firenze - Piattaforma didattica "Facilefacile" - www.ausiliotecafirenze.org/facilefacile Gli alimenti e la piramide alimentare 1. COSA SONO GLI ALIMENTI Si definisce alimento
Ausilioteca del Comune di Firenze - Piattaforma didattica "Facilefacile" - www.ausiliotecafirenze.org/facilefacile Gli alimenti e la piramide alimentare 1. COSA SONO GLI ALIMENTI Si definisce alimento
Le analisi chimiche dell acqua del fiume Padrongianus Scienze Integrate Chimica
 Le analisi chimiche dell acqua del fiume Padrongianus Scienze Integrate Chimica Nell ambito del Progetto relativo all esame dell ecosistema costituito dal Parco Fluviale del Padrongianus sono stati coinvolti
Le analisi chimiche dell acqua del fiume Padrongianus Scienze Integrate Chimica Nell ambito del Progetto relativo all esame dell ecosistema costituito dal Parco Fluviale del Padrongianus sono stati coinvolti
Glossario. Intero quaderno di lavoro. Riferimento. Gli alunni scrivono come titolo i concetti che si riferiscono alle brevi spiegazioni.
 Livello 2 06 / Il sangue Informazione per gli insegnanti 1/7 Riferimento Intero quaderno di lavoro Compito Gli alunni scrivono come titolo i concetti che si riferiscono alle brevi spiegazioni. Materiale
Livello 2 06 / Il sangue Informazione per gli insegnanti 1/7 Riferimento Intero quaderno di lavoro Compito Gli alunni scrivono come titolo i concetti che si riferiscono alle brevi spiegazioni. Materiale
Aspetti nutrizionali
 Aspetti nutrizionali Qualità tangibile: Caratteristiche organolettiche: - consistenza (caratt. meccaniche, geometriche, chimiche) - aspetto (forma, dimensione, colore) - aroma (sapore, odore) Caratteristiche
Aspetti nutrizionali Qualità tangibile: Caratteristiche organolettiche: - consistenza (caratt. meccaniche, geometriche, chimiche) - aspetto (forma, dimensione, colore) - aroma (sapore, odore) Caratteristiche
I differenti materiali differiscono per le caratteristiche meccaniche e fisiche.
 MATERIALI COMPOSITI I differenti materiali differiscono per le caratteristiche meccaniche e fisiche. I METALLI hanno forma cristallina e forti legami molecolari (legame metallico), che danno loro resistenza
MATERIALI COMPOSITI I differenti materiali differiscono per le caratteristiche meccaniche e fisiche. I METALLI hanno forma cristallina e forti legami molecolari (legame metallico), che danno loro resistenza
FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua
 FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua Fisiologia Generale e dell Esercizio Carlo Capelli Facoltà di Scienze Motorie Università degli Studi Verona Obiettivi Funzioni
FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua Fisiologia Generale e dell Esercizio Carlo Capelli Facoltà di Scienze Motorie Università degli Studi Verona Obiettivi Funzioni
Barghini G. L età evolutiva termina a 14 anni? No continua dopo i 14 anni.
 Barghini G. L età evolutiva termina a 14 anni? No continua dopo i 14 anni. Il D.P.R. 28.7.2000 n. 272 con l art. 1 stabilì I medici specialisti in pediatria.sono parte attiva.nel settore preposto alla
Barghini G. L età evolutiva termina a 14 anni? No continua dopo i 14 anni. Il D.P.R. 28.7.2000 n. 272 con l art. 1 stabilì I medici specialisti in pediatria.sono parte attiva.nel settore preposto alla
SISTEMA SCHELETRICO. Il Sistema scheletrico è fatto delle ossa che formano e sostengono il nostro corpo.
 2 SISTEMA SCHELETRICO Il Sistema scheletrico è fatto delle ossa che formano e sostengono il nostro corpo. Lo scheletro è molto importante e ci serve per tre motivi principali: 1. Protegge i nostri organi
2 SISTEMA SCHELETRICO Il Sistema scheletrico è fatto delle ossa che formano e sostengono il nostro corpo. Lo scheletro è molto importante e ci serve per tre motivi principali: 1. Protegge i nostri organi
Cinque prominenze (gruppi di cellule) sono legate allo sviluppo facciale: una frontonasale, due mascellari, due mandibolari. Fig.1
 sono legate allo sviluppo facciale: una frontonasale, due mascellari, due mandibolari. Fig.1") SVILUPPO FACCIALE Lo studio dello sviluppo della faccia, del labbro e del palato durante l accrescimento intrauterino, è importante per capire la tempistica, la complessità e la multifattorialità dei processi
SVILUPPO FACCIALE Lo studio dello sviluppo della faccia, del labbro e del palato durante l accrescimento intrauterino, è importante per capire la tempistica, la complessità e la multifattorialità dei processi
Università degli Studi di Firenze
 La storia delle RGA è cominciata 12 000-14 000 anni fa, durante la rivoluzione agricola del primo Neolitico con la domesticazione delle principali colture e animali. Dopo gli eventi di domesticazione iniziale,
La storia delle RGA è cominciata 12 000-14 000 anni fa, durante la rivoluzione agricola del primo Neolitico con la domesticazione delle principali colture e animali. Dopo gli eventi di domesticazione iniziale,
E COME FACCIAMO A DISTINGUERE UNA VERDURA DALL ALTRA? E SOPRATTUTTO COSA E UN FRUTTO?
 MA COSA CI PERMETTE DI RICONOSCERE I VARI TIPI DI FRUTTA? E COME FACCIAMO A DISTINGUERE UNA VERDURA DALL ALTRA? E SOPRATTUTTO COSA E UN FRUTTO? E UNA VERDURA? PRIMA DI TUTTO OCCORRE PRECISARE CHE IL FRUTTO
MA COSA CI PERMETTE DI RICONOSCERE I VARI TIPI DI FRUTTA? E COME FACCIAMO A DISTINGUERE UNA VERDURA DALL ALTRA? E SOPRATTUTTO COSA E UN FRUTTO? E UNA VERDURA? PRIMA DI TUTTO OCCORRE PRECISARE CHE IL FRUTTO
Introduzione alle macchine a stati (non definitivo)
") Introduzione alle macchine a stati (non definitivo) - Introduzione Il modo migliore per affrontare un problema di automazione industriale (anche non particolarmente complesso) consiste nel dividerlo in
Introduzione alle macchine a stati (non definitivo) - Introduzione Il modo migliore per affrontare un problema di automazione industriale (anche non particolarmente complesso) consiste nel dividerlo in
SCUOLA PRIMARIA DI MAGGIATE Classe terza
 SCUOLA PRIMARIA DI MAGGIATE Classe terza Anno scolastico 2012-2013 TERRE ACQUE Osservando il nostro pianeta possiamo notare che la maggior parte della superficie è occupata dalle acque. Infatti esse occupano
SCUOLA PRIMARIA DI MAGGIATE Classe terza Anno scolastico 2012-2013 TERRE ACQUE Osservando il nostro pianeta possiamo notare che la maggior parte della superficie è occupata dalle acque. Infatti esse occupano