Potenziale di membrana
|
|
|
- Letizia Blasi
- 5 anni fa
- Visualizzazioni
Transcript

1 Potenziale di membrana

2 Il potenziale di membrana può essere misurato introducendo nella cellula un microelettrodo costituito da una sottile pipetta di vetro riempita con una soluzione di elevata conduttività (3 M KCl).
3 Il potenziale di membrana delle cellule
4 Si distinguono due tipi di potenziali di membrana: i potenziali di equilibrio ed i potenziali di diffusione; il meccanismo ionico che li genera non è sostanzialmente diverso, tanto che i primi possono essere considerati un caso particolare dei secondi. un potenziale d'equilibrio si genera quando la membrana è impermeabile ad almeno una delle specie ioniche presenti e pemeabile ad almeno un altra, che sia in concentrazione ineguale ai due lati della membrana. Una volta stabilitosi, si mantiene indefinitamente; un potenziale di diffusione, invece, si genera quando la membrana è permeabile in misura diversa a più specie ioniche inegualmente concentrate ai suoi due lati. Non si mantiene indefinitamente, ma, in assenza di altri fattori, decade progressivamente in modo spontaneo e tende lentamente ad estinguersi.

5 Genesi di un potenziale di equilibrio
6 Che valore assumerà la differenza di potenziale a cavallo di una membrana (V1-V2 = Vm) quando un generico ione (i) si trova alle concentrazioni [i]1 ed [i]2 ai suoi due lati ed abbia raggiunto l'equilibrio elettrochimico? Poiché si tratta di un equilibrio in cui si stabiliscono, attraverso la membrana, due flussi unidirezionali dello ione (i) di uguale intensità e di opposta direzione, mossi rispettivamente dal gradiente di concentrazione e dal gradiente elettrico, il lavoro compiuto a spese del gradiente elettrico (We) per trasferire una Mole dello ione (i) in una direzione sarà uguale e contrario al lavoro (Wc) compiuto a spese del gradiente di concentrazione per muovere la Mole in senso opposto. Perciò:
7 We = -Wc e quindi We + Wc = 0 D'altro canto il lavoro We, per la definizione di potenziale elettrico, sarà We=zi F Vm ove zi è la valenza dello ione (i), F la costante di Faraday Wc invece sarà: Wc = RT {ln[i]1 - ln[i]2} ove R è la costante dei gas, T la temperatura assoluta, e ln[i]1 e ln[i]2 sono i logaritmi naturali delle concentrazioni dello ione (i) ai due lati della membrana. Per lo ione (i) all'equilibrio, quindi, poiché We = -Wc, si avrà: e quindi: Zi F Vm = RT {ln[i]2 - ln[i]1} Vm = Ei = RT zif [i]2 ln (1) [i]1
8 La (1) è l equazione di Nernst ed esprime il concetto che all equilibrio, il flusso netto transmembranario di uno ione diffusibile è nullo. Attribuendo nella 1) alle diverse costanti il loro valore numerico e trasformando i logaritmi naturali in decimali, si ha a 18 C: Ei = Vm = 0,058 {Log [i]2 /[i]1} Ne deriva che a 18 C, la differenza di potenziale varia di 0,058 Volt (58 mv) quando il rapporto tra le concentrazioni dello ione diffusibile ai lati della membrana varia di 10 volte.
9 POTENZIALE DI DIFFUSIONE Un potenziale di diffusione si genera quando la membrana è permeabile in misura diversa a più specie ioniche inegualmente concentrate ai suoi due lati. Un potenziale di diffusione si osserva in una situazione di squilibrio elettrochimico, in cui per due o più specie ioniche vi è un passaggio transmembranario netto di ioni nell'una o nell' altra direzione, secondo il loro gradiente di concentrazione. Esso non si mantiene indefinitamente, ma, in assenza di altri fattori, decade progressivamente in modo spontaneo e tende lentamente ad estinguersi.
10 Genesi di un potenziale di diffusione
11 Si possono allora così riassumere le due condizioni necessarie perché si generi tra i due lati di una membrana un potenziale di diffusione: 1) che esistano differenze di concentrazione (quindi gradienti chimici) che spingano ioni permeanti di tipo diverso a diffondere attraverso la membrana; 2) che la membrana sia diversamente permeabile alle specie ioniche permeanti.
12 Riferendoci all esempio A visto prima, nel quale viene ipotizzato un potenziale di diffusione generato dall ineguale concentrazione di NaCl ai due lati della membrana diversamente permeabile al Na + e al Cl -, all equilibrio elettrico, quando la corrente attraverso la membrana globale dei due ioni è nulla, cioè quando si ha l uguaglianza dei due flussi ionici, FNa = FCl, il potenziale di membrana Vm può essere descritto dall equazione: Vm RT ln PNa[Na+ ]1 + PCl [Cl - ]2 = F PNa[Na + ]2 + PCl [Cl - ]1 che viene detta equazione di Goldman per il potenziale o equazione di Goldman, Hodgkin e Katz.
13 Non potrà certo sfuggire che l'equazione di Goldman per il potenziale richiama quella di Nernst 1) poiché le concentrazioni degli ioni delle diverse specie sono moltiplicate per i rispettivi coefficienti di permeabilità membranale (Pi) che non figurano nell' equazione di Nernst, la differenza di concentrazione tra i due lati della membrana di ogni ione permeante avrà un "peso" sul valore di Vm tanto maggiore quanto maggiore è il suo coefficiente di permeabilità; in altri termini, Vm assumerà un valore intermedio tra i potenziali di equilibrio voluti dalle diverse specie ioniche che sarà sempre più vicino al potenziale di equilibrio dello ione con coefficiente P più elevato e più lontano da quello dello ione con coefficiente P più basso;
14 2) al limite, se la permeabilità per una delle specie ioniche fosse nulla, Vm dipenderebbe solo dalle differenze di concentrazione delle altre specie; se P fosse diverso da zero solo per una delle specie ioniche, Vm coinciderebbe con il potenziale di equilibrio di questa specie e l'equazione di GHK si ridurrebbe all'equazione di Nernst.
15 Natura del potenziale di membrana Il potenziale di membrana origina dalla ineguale distribuzione degli ioni inorganici tra il liquido intra- ed extracellulare. In generale si ha: 1) che nel liquido extra-cellulare si trovano in maggiore concentrazione il Na+ e il Cl-; 2) che nel liquido intra-cellulare sia molto più concentrato il K+, gli anioni sono in prevalenza Pr-
16 Sebbene in misura diversa, la membrana cellulare è permeabile a tutte e tre le specie di ioni inorganici che si trovano inegualmente concentrate ai suoi due lati; d'altro canto è facile dimostrare che il potenziale di membrana non ha il valore calcolabile con la equazione di Nernst per le concentrazioni in tra- ed extra-cellulari dei tre ioni (Na+, K + e Cl-). Infatti, riferendoci per semplicità al caso della fibra muscolare di anfibio e calcolando i potenziali di equilibrio dei tre ioni (ENa, EK ed ECl) in base ai valori delle loro concentrazioni esterne ed interne, si ha ENa = + 45 mv, EK = = -102 mv, ECl = -88 mv mentre il potenziale di membrana (Vm), misurato con derivazione intracellulare nella fibra muscolare, è risultato di -90 mv (interno negativo).
17 Il potenziale di membrana della fibra muscolare (come quello di tutte le cellule), poiché è generato dalla ineguale concentrazione di specie ioniche diffusibili che non si trovano all'equilibrio elettrochimico, ha i caratteri di un potenziale di diffusione; gli ioni permeanti si trovano in una condizione di equilibrio elettrico ma non di equilibrio chimico. Il suo valore quindi deve essere calcolabile con l'equazione di GHK per il potenziale applicata alle tre specie ioniche permeanti la membrana cellulare: Vm = RT ln [K+]e PK+ [Na+]e PNa+ [Cl-]i PCl F [K+]i PK+ [Na+]i PNa+ [Cl-]e PCl
18 Potenziale di membrana e correnti ioniche Un approccio diverso è quello di considerare la membrana come un conduttore elettrico, capace di consentire il passaggio, per ciascuna specie ionica permeante (i), di una corrente ionica specifica (Ii); allora, se si accetta per semplicità che la corrente transmembranale di ogni specie ionica ubbidisca alla legge di Ohm, la sua intensità sarà pari alla differenza di potenziale che muove lo ione, moltiplicata per la conduttanza della membrana (gi) nei riguardi dello ione stesso: Ii = V gi
19 D'altro canto la differenza di potenziale che costituisce la "driving force" dello ione i non è semplicemente la differenza di potenziale effettivamente presente ai due lati della membrana (Vm), ma è pari a Vm - Ei, cioè alla differenza tra il potenziale di membrana ed il potenziale di equilibrio voluto dall' equazione di Nernst per quello ione; a questo potenziale di equilibrio infatti la corrente netta dello ione i sarebbe nulla. Si avrà allora che: Ii = (Vm- Ei) gi = Vm gi - Ei gi Partendo da questa relazione è possibile calcolare il valore del potenziale generato a cavallo della membrana da qualunque numero di specie ioniche permeanti, quando siano noti i potenziali di equilibrio degli ioni delle diverse specie e le rispettive conduttanze membranarie.
20 Riferendoci alle fibre muscolari scheletriche dei vertebrati ed escludendo il cloro, avremo -una corrente di potassio, diretta dall interno verso l esterno spinta dal gradiente di concentrazione IK = (Vm EK) gk = [-90 (-102)] gk =12 gk -una corrente di sodio diretta invece dall esterno verso l interno, spinta sia dal gradiente di concentrazione che da quello elettrico INa = (Vm ENa) gna = [-90 (+45)] gna = -135 gna E poiché al potenziale di riposo INa = -IK 12 gk = 135 gna Da ciò deriva che la conduttanza per il potassio deve essere circa 11 volte maggiore di quella del sodio.
21 Da questa semplice analisi risulta che: Vm = ENa gna + EK gk gna + + gk + la quale esprime che la differenza di concentrazione tra i due lati della membrana di ogni ione permeante avrà un peso sul valore finale di Vm tanto maggiore quanto maggiore sarà la conduttanza della membrana nei suoi confronti.
22 Genesi e mantenimento del potenziale di membrana Nella genesi e nel mantenimento del potenziale di membrana intervengono: 1) processi ionici passivi, consistenti nella diffusione secondo gradiente attraverso la membrana cellulare degli ioni (fondamentalmente K + e Na+) che genera direttamente la differenza di potenziale ai due lati della membrana stessa; 2) processi ionici attivi, che consistono nel trasporto contro gradiente attraverso al membrana dei due ioni Na+ e K+; questo trasporto, di direzione opposta alla diffusione dei due ioni, crea e mantiene i gradienti di concentrazione dei due ioni ai due lati della membraqa. Questo trasporto attivo è operato dalla pompa ionica di scambio Na+ /K+.

23 Flussi ionici attivi e passivi di Na e K
24 La membrana ed i segnali elettrici Il potenziale di membrana è essenziale per molte funzioni cellulari. Il potenziale di membrana ha un ruolo fondamentale nella comunicazione di "informazione" contenuta in segnali elettrici, sia tra parti diverse di una stessa cellula, sia tra cellule vicine. La più vistosa di queste manifestazioni elettriche "attive" è indubbiamente il potenziale d'azione, quel brevissimo impulso che si propaga, ripetuto con varia frequenza, nelle fibre nervose e nelle fibre muscolari; anche i potenziali sinaptici, che si osservano nelle giunzioni interneuroniche e neuromuscolari, nonché i potenziali recettoriali degli organi di senso, hanno il carattere di segnali ricchi di informazione.
25 Una spiegazione del modo con cui si generano i diversi tipi di "segnali" elettrici nelle cellule eccitabili (e della loro trasmissione) richiede tuttavia la preliminare conoscenza del comportamento di una membrana cellulare quando essa sia sede di variazioni di potenziale in funzione del tempo. Occorre in altri termini studiare il potenziale di membrana non più nelle condizioni stazionarie fin qui considerate, ma in condizioni dinamiche.
26 Ciò è realizzabile sperimentalmente quando si fa passare attraverso la membrana cellulare una corrente elettrica opportunamente regolata.
 ed una capacità (Cm).")
27 Questo tipo di analisi parte dall' assunto che la membrana di una cellula sia semplicemente assimilabile ad un conduttore elettrico che presenti una resistenza elettrica (Rm), o meglio una conduttanza elettrica (gm = 1/Rm) ed una capacità (Cm). Sia l'una che l'altra di queste due caratteristiche corrispondono ad elementi strutturali distinti della membrana plasmatica:
28 1) la resistenza dipende infatti dal numero e dalla permeabilità dei diversi canali ionici che attraversano la membrana e consentono il passaggio delle particelle cariche (gli ioni); 2) la capacità dipende invece dal potere che possiede il doppio strato fosfolipidico di accumulare ai due versanti membranali due strati di ioni di segno opposto. Infatti le catene idrocarburiche dei fosfolipidi, disposte nello spessore della membrana, essendo "apolari", costituiscono uno strato isolante analogo al dielettrico di un condensatore, mentre i gruppi fosfatidici "polari" che si affacciano alle due superfici membranali (l'interna e l'esterna) possono accumulare o cedere, grazie alla loro dissociazione, cariche elettriche in modo analogo alle armature di un condensatore.
29 Il metodo del current clamp Le proprietà elettriche passive della membrana plasmatica si possono misurare utilizzando un procedimento di tipo "ohmmetrico" noto come metodo del "blocco della corrente" (o "current clamp"), che consiste nel misurare la differenza di potenziale generata a cavallo della membrana dal passaggio di una corrente di intensità nota. A tal fine è necessario infiggere nella cellula due microelettrodi: un elettrodo di stimolazione e un elettrodo di derivazione.
30 Schema del metodo del current-clamp (blocco della corrente)
31 Con questo procedimento è possibile studiare le caratteristiche elettriche passive della membrana cellulare sia quando la corrente che l'attraversa fluisca nel senso di depolarizzarla (cioè quando l'elettrodo interno positivo rispetto all eletrodo esterno), sia quando fluisca nel senso di iperpolarizzarla (cioè quando l'elettrodo interno venga reso negativo rispetto all altro).
32 Si utilizzano di solito brevi correnti (di durata da 10 a 100 msec) di intensità regolabile, cioè "impulsi di corrente" nei quali la corrente passa istantaneamente dal valore zero a quello che è stato prescelto ed altrettanto istantaneamente viene interrotta. Impulsi di questo tipo sono detti "rettangolari" per l'aspetto del diagramma che ne rappresenta l'andamento temporale. L inizio e la fine di ciascun impulso sono definiti da due "gradini di corrente" (o "steps") di uguale ampiezza e di direzione opposta, separati da un "pianerottolo" in cui la corrente fluisce con intensità costante.
33 Un esempio dei risultati ottenibili con la tecnica del "current clamp" è riportato nella figura stessa, ove sono illustrate in funzione del tempo le variazioni del potenziale di membrana (il Vm) durante il passaggio di due " impulsi rettangolari" di corrente di uguale intensità (Im), uno in senso depolarizzante (A) ed uno in senso iperpolarizzante (B); si noterà subito che, in corrispondenza del "gradino" iniziale di ogni impulso, le variazioni del potenziale di membrana (C e D) non s'instaurano istantaneamente, ma raggiungono il loro valore definitivo con legge esponenziale crescente; analogamente, in corrispondenza del "gradino" finale dell'impulso, il potenziale di membrana ritorna al suo valore di riposo con un andamento esponenziale decrescente, speculare al primo.

34
35 Questo comportamento della membrana cellulare di fronte al passaggio di un "gradino di corrente" è coerente con quello di un circuito equivalente in cui ad una resistenza (Rm) è collegato in parallelo un condensatore (Cm). Ai terminali (A e B) comuni di Rm e Cm (corrispondenti all'esterno ed all'interno della membrana) sono idealmente collegati: da un lato lo strumento che misura le differenze di potenziale a cavallo della membrana (millivoltmetro), e dall' altro il generatore degli impulsi di corrente.
36 Come avverrebbe in un circuito di questo genere, anche nella membrana cellulare la corrente erogata dal generatore (Im) si divide in due componenti: una componente resistiva (IR = Vm/Rm, che percorre la resistenza membranale (Rm) e determina una variazione del potenziale ai suoi capi, ed una componente capacitiva (IC = Cm dvm/dt), che va a caricare la capacità membranale (Cm).
 finché, quando Cm sarà completamente caricato")
37 Al "gradino" iniziale di un impulso di corrente tutte le cariche portate dalla corrente costante andranno a caricare la capacità Cm; IC sarà massima ed IR minima. In seguito, man mano che Cm si carica, IC si riduce ed aumenta IR; di conseguenza aumenta la differenza di potenziale ai due lati della membrana, ( Vm) finché, quando Cm sarà completamente caricato al valore corrispondente al "pianerottolo" dell'impulso, tutta la corrente (Im) passerà attraverso Rm. IR allora sarà massima ed IC nulla, e Vm raggiungerà un valore finale stazionario (Vf) che verrà mantenuto finché dura il passaggio della corrente.
38 La legge temporale con cui Vm si modifica, partendo dal valore iniziale (V0) per raggiungere il valore finale (Vf), è definita dalla seguente relazione esponenziale: t - Rm Cm) Vm=V0 + (Vf V0) (1- e Al "gradino" finale dell'impulso, il generatore cessa di erogare corrente e Vm si estingue secondo la seguente relazione temporale t - Rm Cm) Vm=V0 + (Vf V0) (e
39 Il prodotto Rm Cm che figura all' esponente delle equazioni prende il nome di "costante di tempo" ( ) della membrana, ed il suo valore esprime la maggiore o minore velocità con cui Vm raggiunge il valore finale Vf al "gradino" iniziale dell'impulso di corrente, e ritorna al valore V0 al "gradino" finale. (Rm Cm) ha le dimensioni di un tempo ed è numericamente pari al tempo necessario affinché, al gradino iniziale dell impulso, Vm aumenti di un valore pari al 63% di (Vf-V0) oppure, al gradino finale, discenda al 37% di (Vf-V0).
40 La resistenza membranale (Rm) e la capacità membranale (Cm), possono essere dedotti dalla registrazione di Vm durante l'erogazione anche di un solo impulso di corrente di cui sia nota l'intensità (Im). Infatti se anche per la membrana cellulare vale la legge di Ohm come per il circuito equivalente, il valore di Rm sarà uguale al rapporto (Vf V0)/Im o) quando Vm abbia raggiunto il suo valore finale (Vf) al "pianerottolo" dell'impulso di corrente. Nota Rm, potrà essere calcolata anche Cm deducendola dal prodotto Rm Cm, cioè ad esempio dal tempo necessario perché Vm raggiunga il 63% di (Vf-V0) al "gradino" iniziale dell'impulso di corrente.
41 Utilizzando il metodo del "current clamp" è possibile determinare, anche con un solo impulso di corrente, il valore della conduttanza membranale (gm = l/rm) nei limiti in cui la membrana cellulare come conduttore ubbidisce alla legge di Ohm; ciò significa assumere che il valore di gm sia costante ed indipendente dalla intensità della corrente Im che attraversa la membrana. Ciò purtroppo non si realizza nel caso di molte cellule.
42 Proprietà elettriche passive e attive della membrana
43 I potenziali di azione Nel potenziale d azione si distinguono due fasi: -fase di DEPOLARIZZAZIONE -fase di RIPOLARIZZAZIONE
44 La legge del tutto o nulla
45 Genesi ionica del potenziale di azione Gli studi di Hodgkin ed Huxley con il metodo del voltage clamp sull assone gigante di calamaro hanno permesso di capire quali sono gli ioni coinvolti nella genesi del potenziale di azione.
46 Il potenziale d azione si genera in seguito a una depolarizzazione grazie alla presenza di : - canali ionici voltaggio dipendenti nella membrana - gradienti elettrochimici di ioni tra cellula e ambiente extracellulare


47 Canale per il K + Canale con una porta o cancello

48 Canale per il Na + Canale con due porte o cancelli Il canale del Na voltaggio dipendente ha due porte: di attivazione e di inattivazione.

49 SEQUENZA DI EVENTI DURANTE IL POTENZIALE DI AZIONE Al potenziale di riposo il cancello di attivazione è chiuso

50 In seguito ad una depolarizzazione liminare la porta di attivazione si apre velocemente, il sodio entra nella cellula e depolarizza fino a invertire la polarità della membrana, il potenziale tende al valore del potenziale di equilibrio del Na

51 La depolarizzazione della membrana fa chiudere la porta di inattivazione si chiude. Un canale inattivato non può aprirsi se la membrana rimane depolarizzata.


52 + L apertura di canali per il K determina fuoriuscita di potassio e la ripolarizzazione della membrana K K + Durante la ripolarizzazione le due porte del sodio ritornano alle posizioni originali.


53 Quando il potenziale di membrana è ritornato al valore del potenziale di riposo anche la porta del canale del K si chiude. K +
54 RIASSUMENDO La depolarizzazione della membrana provoca 1. Un rapido aumento della gna (attivazione), seguito da 2. un suo declino dovuto ad un secondo processo chiamato inattivazione della gna 3. un lento aumento della conduttanza al K (attivazione)
55
56 Il potenziale d azione E la risposta ad uno stimolo depolarizzante che possono dare cellule elettricamente eccitabili, cioè provviste di un corredo di canali ionici voltaggio-dipendenti per il Na + e per il K + Significato funzionale: Nei neuroni segnale elettrico che propagandosi lungo la fibra nervosa consente la trasmissione di messaggi elettrici Nelle fibrocellule muscolari innesca il processo della contrazione
57 Caratteristiche generali del potenziale d azione La soglia Lo stimolo soglia è lo stimolo depolarizzante di intensità minima in grado di generare un potenziale d azione in un neurone La legge del tutto o nulla In un neurone un potenziale d azione è generaro e si sviluppa in tutta la sua ampiezza se lo stimolo raggiunge o supera la soglia, oppure non è generato affatto, se l ampiezza dello stimolo è inferiore alla soglia. La refrattarietà Un neurone, una volta generato un potenziale d azione viene a trovarsi in uno stato di refrattarietà - periodo di refrattarietà assoluta: nessuno stimolo per quanto intenso è in grado di generare un secondo potenziale d azione - periodo di refrattarietà relativa: un secondo stimolo, a condizione che sia sufficientemente più intenso di quello soglia, è in grado di generare un secondo potenziale d azione
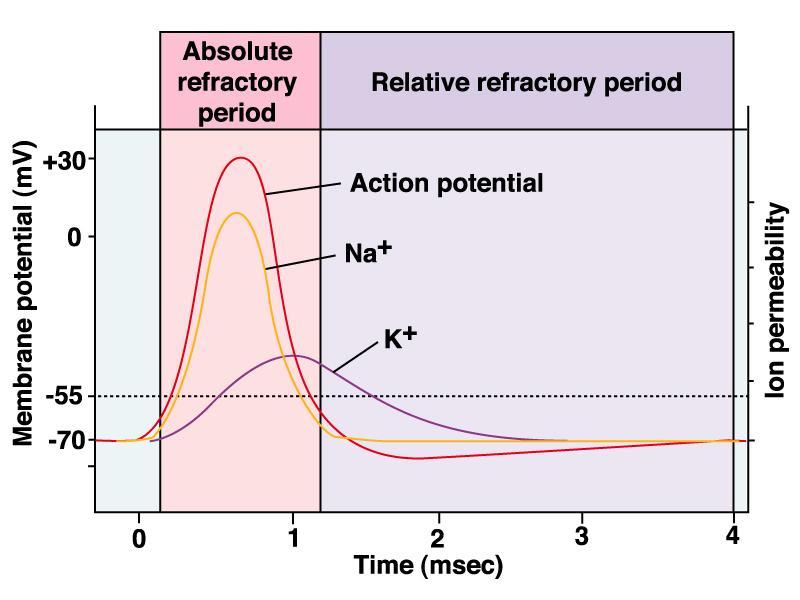
58
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.
 Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
POTENZIALE DI MEMBRANA
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Che ruolo ha la membrana plasmatica nei potenziali di diffusione?
 Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
Il potenziale di membrana è la differenza di potenziale elettrico a cavallo della membrana cellulare (negatività interna), determinato da una diversa
, determinato da una diversa") Il potenziale di membrana è la differenza di potenziale elettrico a cavallo della membrana cellulare (negatività interna), determinato da una diversa distribuzione ionica ai due lati della membrana. Nelle
Il potenziale di membrana è la differenza di potenziale elettrico a cavallo della membrana cellulare (negatività interna), determinato da una diversa distribuzione ionica ai due lati della membrana. Nelle
4. IL SISTEMA ENDOCRINO: GENERALITA
 4. IL SISTEMA ENDOCRINO: GENERALITA 1. Ghiandole endocrine e ormoni 2. Il tessuto bersaglio 3. Rilascio degli ormoni 4. Il concetto della regolazione retroattiva negativa 5. Un esempio di regolazione ormonale:
4. IL SISTEMA ENDOCRINO: GENERALITA 1. Ghiandole endocrine e ormoni 2. Il tessuto bersaglio 3. Rilascio degli ormoni 4. Il concetto della regolazione retroattiva negativa 5. Un esempio di regolazione ormonale:
Potenziale di membrana (I)
") Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Potenziale di membrana (I) Misura della differenza di potenziale tra i due lati della membrana Scaricato da www.sunhope.it 1
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Potenziale di membrana (I) Misura della differenza di potenziale tra i due lati della membrana Scaricato da www.sunhope.it 1
Genesi del Potenziale di Membrana. In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma
 Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma Forze agenti sugli ioni Gradiente di Concentrazione Campo Elettrico Energia
Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma Forze agenti sugli ioni Gradiente di Concentrazione Campo Elettrico Energia
otenziali graduati (elettrotonici) potenziale d azione
 potenziale d azione") I segnali elettrici che si generano nei neuroni: potenziali graduati (elettrotonici) potenziale d azione sono modificazioni del potenziale di riposo determinate dall apertura o chiusura di canali ionici.
I segnali elettrici che si generano nei neuroni: potenziali graduati (elettrotonici) potenziale d azione sono modificazioni del potenziale di riposo determinate dall apertura o chiusura di canali ionici.
POTENZIALE DI EQUILIBRIO DEL K + la differenza di potenziale ai due lati della membrana utile alla formazione dell equilibrio elettrochimico del K +
 POTENZIALE DI EQUILIBRIO DEL K + la differenza di potenziale ai due lati della membrana utile alla formazione dell equilibrio elettrochimico del K + P - K + P - P - P - P - P - Na + Cl - -80 mv Voltometro
POTENZIALE DI EQUILIBRIO DEL K + la differenza di potenziale ai due lati della membrana utile alla formazione dell equilibrio elettrochimico del K + P - K + P - P - P - P - P - Na + Cl - -80 mv Voltometro
Confronto della voltaggio-dipendenza di g K e g Na
 Confronto della voltaggio-dipendenza di g K e g Na 1.0 0.8 ttanza g Condut 0.6 0.4 0.2 0.0-60 -40-20 0 20 40 60 V m (mv) Il potenziale d azione E la risposta ad uno stimolo depolarizzante che possono dare
Confronto della voltaggio-dipendenza di g K e g Na 1.0 0.8 ttanza g Condut 0.6 0.4 0.2 0.0-60 -40-20 0 20 40 60 V m (mv) Il potenziale d azione E la risposta ad uno stimolo depolarizzante che possono dare
Potenziale di diffusione
 Potenziale di diffusione La diffusione di più specie ioniche attraverso la membrana secondo il loro gradiente di concentrazione può creare essa stessa differenze di potenziale elettrico a cavallo di una
Potenziale di diffusione La diffusione di più specie ioniche attraverso la membrana secondo il loro gradiente di concentrazione può creare essa stessa differenze di potenziale elettrico a cavallo di una
POTENZIALE DI MEMBRANA FGE AA
 POTENZIALE DI MEMBRANA FGE AA.2016-17 OBIETTIVI GENESI E MANTENIMENTO DEL POTENZIALE DI MEMBRANA A RIPOSO POTENZIALE DI EQUILIBRIO ED EQUAZIONE DI NERNST EQUAZIONE DI GOLDMAN E PMR DEL NEURONE CANALI IONICI
POTENZIALE DI MEMBRANA FGE AA.2016-17 OBIETTIVI GENESI E MANTENIMENTO DEL POTENZIALE DI MEMBRANA A RIPOSO POTENZIALE DI EQUILIBRIO ED EQUAZIONE DI NERNST EQUAZIONE DI GOLDMAN E PMR DEL NEURONE CANALI IONICI
Il potenziale di membrana a riposo
 Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
Diffusione Equazione di Nernst e Potenziale di Membrana. scaricatoda
 Diffusione Equazione di Nernst e Potenziale di Membrana Perché parlare del concetto di diffusione? E una proprietà fisica fondamentale di tutti i processi biologici e costituisce il motore tramite il quale
Diffusione Equazione di Nernst e Potenziale di Membrana Perché parlare del concetto di diffusione? E una proprietà fisica fondamentale di tutti i processi biologici e costituisce il motore tramite il quale
La pompa sodio-potassio
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA
 FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
Genesi del Potenziale di Membrana. In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma
 Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma Forze agenti sugli ioni Gradiente di Concentrazione Campo Elettrico Energia
Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo del plasmalemma Forze agenti sugli ioni Gradiente di Concentrazione Campo Elettrico Energia
POTENZIALE DI MEMBRANA
 POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
L informazione nervosa si basa sulla capacità dei neuroni di generare correnti elettriche, in seguito a modificazioni del potenziale di riposo che
 L informazione nervosa si basa sulla capacità dei neuroni di generare correnti elettriche, in seguito a modificazioni del potenziale di riposo che risultano dall apertura o chiusura di canali ionici. I
L informazione nervosa si basa sulla capacità dei neuroni di generare correnti elettriche, in seguito a modificazioni del potenziale di riposo che risultano dall apertura o chiusura di canali ionici. I
Ioni fosfato e proteine (anioni) sono prevalenti nella cellula
 sono prevalenti nella cellula") Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Per poter misurare una ddp ai capi di una membrana cellulare occorrono due elettrodi conduttori (uno di misura e uno di riferimento) che permettono
 che permettono") 1 Per poter misurare una ddp ai capi di una membrana cellulare occorrono due elettrodi conduttori (uno di misura e uno di riferimento) che permettono di misurare separatamente il potenziale intracellulare
1 Per poter misurare una ddp ai capi di una membrana cellulare occorrono due elettrodi conduttori (uno di misura e uno di riferimento) che permettono di misurare separatamente il potenziale intracellulare
Corso di Laurea Magistrale in Medicina e Chirurgia. Biofisica e Fisiologia I A.A ESERCITAZIONE. Prof. Clara Iannuzzi
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 ESERCITAZIONE Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 ESERCITAZIONE Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Prof. Clara Iannuzzi
 Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica I A.A. 2015/2016 Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale Via L. De Crecchio 7, Napoli Seconda
Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica I A.A. 2015/2016 Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale Via L. De Crecchio 7, Napoli Seconda
Membrane Biologiche. Caratteristiche delle membrane biologiche
 Membrane Biologiche Caratteristiche delle membrane biologiche 1. Le membrane sono strutture a foglio che formano setti di separazione tra compartimenti con composizione diversa tra loro. 2. Sono costituite
Membrane Biologiche Caratteristiche delle membrane biologiche 1. Le membrane sono strutture a foglio che formano setti di separazione tra compartimenti con composizione diversa tra loro. 2. Sono costituite
 Uno stimolo esterno (es. aumento della temperatura) o interno (es. variazione della osmolarità) viene recepito da recettori e integrato con altri stimoli. In seguito all integrazione viene attivato un
Uno stimolo esterno (es. aumento della temperatura) o interno (es. variazione della osmolarità) viene recepito da recettori e integrato con altri stimoli. In seguito all integrazione viene attivato un
Equilibrio di Gibbs-Donnan
 Equilibrio di GibbsDonnan Tra due soluzioni acquose separate da una membrana che sia impermeabile ad uno solo dei soluti si stabilisce un equilibrio (equilibrio di Donnan) garantito da una differenza di
Equilibrio di GibbsDonnan Tra due soluzioni acquose separate da una membrana che sia impermeabile ad uno solo dei soluti si stabilisce un equilibrio (equilibrio di Donnan) garantito da una differenza di
I nostri compartimenti idrici sono in Equilibrio osmotico ma non chimico
 I nostri compartimenti idrici sono in Equilibrio osmotico ma non chimico Concentrazione dei principali ioni I compartimenti del nostro corpo non sono in equilibrio elettrico ELETTRICITA Atomi: entità elettricamente
I nostri compartimenti idrici sono in Equilibrio osmotico ma non chimico Concentrazione dei principali ioni I compartimenti del nostro corpo non sono in equilibrio elettrico ELETTRICITA Atomi: entità elettricamente
Cellula vista come un circuito resistenzacondensatore
 Segnali elettrici Cellula vista come un circuito resistenzacondensatore La membrana si comporta come un circuito costituito da un condensatore e una resistenza in parallelo. Iniettando un impulso di corrente
Segnali elettrici Cellula vista come un circuito resistenzacondensatore La membrana si comporta come un circuito costituito da un condensatore e una resistenza in parallelo. Iniettando un impulso di corrente
Relazione ohmica tra voltaggio e corrente
 Relazione ohmica tra voltaggio e corrente 6 5 Potenziale soglia Corrente (na) 4 3 2 1 Le membrane neuronali rispondono in modo passivo all iper-polarizzazione e alla depolarizzazione (sotto soglia), non
Relazione ohmica tra voltaggio e corrente 6 5 Potenziale soglia Corrente (na) 4 3 2 1 Le membrane neuronali rispondono in modo passivo all iper-polarizzazione e alla depolarizzazione (sotto soglia), non
Il potenziale di membrana
 Il potenziale di membrana Nel citoplasma della cellula, così come nel liquido extracellulare sono disciolti diversi tipi di ioni. La loro concentrazione non è però uguale dai due lati della membrana citoplasmatica
Il potenziale di membrana Nel citoplasma della cellula, così come nel liquido extracellulare sono disciolti diversi tipi di ioni. La loro concentrazione non è però uguale dai due lati della membrana citoplasmatica
Attività elettrica dei neuroni. Neuroni piramidali del SNC che stanno crescendo su una matrice di glia/fibroblasti
 Attività elettrica dei neuroni Neuroni piramidali del SNC che stanno crescendo su una matrice di glia/fibroblasti Un esperimento in current-clamp Si inietta una corrente I di intensità nota nella cellula
Attività elettrica dei neuroni Neuroni piramidali del SNC che stanno crescendo su una matrice di glia/fibroblasti Un esperimento in current-clamp Si inietta una corrente I di intensità nota nella cellula
POTENZIALE DI MEMBRANA A RIPOSO. Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI
 POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione
 3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Genesi e mantenimento
3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Genesi e mantenimento
FUNZIONE, REGOLAZIONE, NATURA CHIMICA, TIPO DI RECETTORE
 Elena, Serena, Francesca, Valentina, Elisa, Lucrezia, Elisa Lezione 7 1. Asse Ipotalamo-ipofisi-surrene: CRH ACTH - Cortisolo 2. Asse Ipotalamo-ipofisi-ghiandole sessuali: GdRH e gonadotropine ipofisarie:
Elena, Serena, Francesca, Valentina, Elisa, Lucrezia, Elisa Lezione 7 1. Asse Ipotalamo-ipofisi-surrene: CRH ACTH - Cortisolo 2. Asse Ipotalamo-ipofisi-ghiandole sessuali: GdRH e gonadotropine ipofisarie:
Stimolazione artificiale del cuore
 Stimolazione artificiale del cuore Il ciclo del cuore: Impulso elettrico di stimolazione da parte di un gruppo di fibre nervose il nodo sinoatriale, Stimolatori artificiali Pacemaker impiantati chirurgicamente
Stimolazione artificiale del cuore Il ciclo del cuore: Impulso elettrico di stimolazione da parte di un gruppo di fibre nervose il nodo sinoatriale, Stimolatori artificiali Pacemaker impiantati chirurgicamente
Richiami di Fisiologia Cellulare
 Richiami di Fisiologia Cellulare Alcuni dati fondamentali In tutte le cellule 1) vi è una membrana, di natura lipidica, che separa l ambiente interno da quello esterno; nella o sulla membrana sono abbondanti
Richiami di Fisiologia Cellulare Alcuni dati fondamentali In tutte le cellule 1) vi è una membrana, di natura lipidica, che separa l ambiente interno da quello esterno; nella o sulla membrana sono abbondanti
Trasporto selettivo di molecole (trasportatori) Riconoscimento di cellule e substrati (integrine, matrice)
 Riconoscimento di cellule e substrati (integrine, matrice)") Trasporto selettivo di molecole (trasportatori) Riconoscimento di cellule e substrati (integrine, matrice) Comunicazione intercellulare (recettori e proteine trasduttrici) Attività enzimatica Determinazione
Trasporto selettivo di molecole (trasportatori) Riconoscimento di cellule e substrati (integrine, matrice) Comunicazione intercellulare (recettori e proteine trasduttrici) Attività enzimatica Determinazione
LEZIONE 3: POTENZIALE D AZIONE PROPRIETA PASSIVE DI MEMBRANA
 LEZIONE 3: POTENZIALE D AZIONE PROPRIETA PASSIVE DI MEMBRANA 1 I tempi di risposta delle cellule sono determinati dai valori di R e C: τ = RC = costante di tempo della membrana τ = RC = 1 (kω x cm 2 )
LEZIONE 3: POTENZIALE D AZIONE PROPRIETA PASSIVE DI MEMBRANA 1 I tempi di risposta delle cellule sono determinati dai valori di R e C: τ = RC = costante di tempo della membrana τ = RC = 1 (kω x cm 2 )
PROF. ALESSANDRO MALFATTI. Corso di Fisiologia e Benessere degli animali in produzione. STRUTTURA e FUNZIONE
 STRUTTURA e FUNZIONE Ogni neurone presenta 4 zone: DENDRITI: ricezione dell'impulso proveniente da altri neuroni SOMA (PIRENOFORO): integrazione delle numerose afferenze, spesso contrastanti sintesi del
STRUTTURA e FUNZIONE Ogni neurone presenta 4 zone: DENDRITI: ricezione dell'impulso proveniente da altri neuroni SOMA (PIRENOFORO): integrazione delle numerose afferenze, spesso contrastanti sintesi del
BIOELETTRICITÀ. Processo di diffusione passiva 2
 BIOELETTRICITÀ La membrana cellulare è costituita da un sottile strato lipoproteico (spessore 8 nm), che separa il liquido intracellulare dal liquido interstiziale (extracellulare). Il liquido intracellulare
BIOELETTRICITÀ La membrana cellulare è costituita da un sottile strato lipoproteico (spessore 8 nm), che separa il liquido intracellulare dal liquido interstiziale (extracellulare). Il liquido intracellulare
Equazione di Nerst. Equazione di Goldman
 Equazione di Nerst Equazione di Goldman 1 PROPRIETA PASSIVE DEI NEURONI I NEURONI posseggono proprietà passive importanti: Resistenza della membrana a riposo Capacità della membrana Resistenza assiale
Equazione di Nerst Equazione di Goldman 1 PROPRIETA PASSIVE DEI NEURONI I NEURONI posseggono proprietà passive importanti: Resistenza della membrana a riposo Capacità della membrana Resistenza assiale
Comunicazione tra cellule
 Comunicazione tra cellule I segnali-messaggi possono essere: elettrici o chimici Il messaggio deve avere le seguenti caratteristiche: 1. Contenere informazione 2. Essere indirizzato 3. Essere trasmesso
Comunicazione tra cellule I segnali-messaggi possono essere: elettrici o chimici Il messaggio deve avere le seguenti caratteristiche: 1. Contenere informazione 2. Essere indirizzato 3. Essere trasmesso
Fenomeni elettrici che interessano la cellula
 Fenomeni elettrici che interessano la cellula In quasi tutte le cellule dell organism o si generano potenziali elettrici a livello della m em brana cellulare Alcune cellule, come quelle nervose e muscolari
Fenomeni elettrici che interessano la cellula In quasi tutte le cellule dell organism o si generano potenziali elettrici a livello della m em brana cellulare Alcune cellule, come quelle nervose e muscolari
Equazione di Nerst. Equazione di Goldman
 Equazione di Nerst Equazione di Goldman 1 PROPRIETA PASSIVE DEI NEURONI I NEURONI posseggono proprietà passive importanti: Resistenza della membrana a riposo Capacità della membrana Resistenza assiale
Equazione di Nerst Equazione di Goldman 1 PROPRIETA PASSIVE DEI NEURONI I NEURONI posseggono proprietà passive importanti: Resistenza della membrana a riposo Capacità della membrana Resistenza assiale
Tessuto muscolare cardiaco
 Tessuto muscolare cardiaco Le cellule muscolari cardiache sono ramificate, possiedono un unico nucleo e sono unite tra loro da giunzioni specializzate dette dischi intercalari in corrispondenza dei quali
Tessuto muscolare cardiaco Le cellule muscolari cardiache sono ramificate, possiedono un unico nucleo e sono unite tra loro da giunzioni specializzate dette dischi intercalari in corrispondenza dei quali
Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati
 Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati della membrana. Il potenziale di membrana (negativo
Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati della membrana. Il potenziale di membrana (negativo
APPLICAZIONI. studio delle proprietà elettriche delle cellule eccitabili. neuroni, muscolo striato e cardiaco
 Elettrofisiologia APPLICAZIONI studio delle proprietà elettriche delle cellule eccitabili neuroni, muscolo striato e cardiaco POTENZIALE DI MEMBRANA tutte le cellule hanno un potenziale di membrana il
Elettrofisiologia APPLICAZIONI studio delle proprietà elettriche delle cellule eccitabili neuroni, muscolo striato e cardiaco POTENZIALE DI MEMBRANA tutte le cellule hanno un potenziale di membrana il
FISIOLOGIA GENERALE - ESERCITAZIONE n. 1 4 Ott 2017
 FISIOLOGIA GENERALE - ESERCITAZIONE n. 1 4 Ott 2017 Esercizio n. 1 Pressione osmotica In un litro di soluzione sono contenuti 1.5 g di NaCl, 2.9 g di Na 3 C 6 H 5 O 7, 1.5 g di KCl e 2.0 g di C 6 H 12
FISIOLOGIA GENERALE - ESERCITAZIONE n. 1 4 Ott 2017 Esercizio n. 1 Pressione osmotica In un litro di soluzione sono contenuti 1.5 g di NaCl, 2.9 g di Na 3 C 6 H 5 O 7, 1.5 g di KCl e 2.0 g di C 6 H 12
POTENZIALE D AZIONE FGE AA
 POTENZIALE D AZIONE FGE AA.2016-17 OBIETTIVI ORGANIZZAZIONE GENERALE SN POTENZIALE DI AZIONE: DESCRIZIONE DELLE FASI E DETERMINANTI FISICO-CHIMICI ANDAMENTO DELLE CONDUTTANZE DEL SODIO E DEL POTASSIO NEL
POTENZIALE D AZIONE FGE AA.2016-17 OBIETTIVI ORGANIZZAZIONE GENERALE SN POTENZIALE DI AZIONE: DESCRIZIONE DELLE FASI E DETERMINANTI FISICO-CHIMICI ANDAMENTO DELLE CONDUTTANZE DEL SODIO E DEL POTASSIO NEL
Sinapsi neuromuscolare
 Sinapsi neuromuscolare La sinapsi tra cellule nervose e cellule muscolari scheletriche è detta anche sinapsi neuromuscolare o placca motrice. A livello del muscolo l assone della cellula nervosa si suddivide
Sinapsi neuromuscolare La sinapsi tra cellule nervose e cellule muscolari scheletriche è detta anche sinapsi neuromuscolare o placca motrice. A livello del muscolo l assone della cellula nervosa si suddivide
Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata.
 Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata. Costante e stabile Negativa all interno Peculiare di ogni tipo di cellula Il Potenziale di
Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata. Costante e stabile Negativa all interno Peculiare di ogni tipo di cellula Il Potenziale di
Propagazione del potenziale d azione
 Propagazione del potenziale d azione Una volta generato, il potenziale d azione si propaga senza decremento lungo l assone, partendo dal monticolo assonico e giungendo al terminale assonico. Un potenziale
Propagazione del potenziale d azione Una volta generato, il potenziale d azione si propaga senza decremento lungo l assone, partendo dal monticolo assonico e giungendo al terminale assonico. Un potenziale
l Trasporto attraverso le membrane
 l Trasporto attraverso le membrane MOVIMENTO DI SOSTANZE ATTRAVERSO LA MEMBRANA GARANTISCE UNA SELETTIVITA ED AVVIENE PASSIVAMENTE: DIFFUSIONE ATTIVAMENTE: RICHIEDE ENERGIA SEMPLICE DELL ACQUA FACILITATA
l Trasporto attraverso le membrane MOVIMENTO DI SOSTANZE ATTRAVERSO LA MEMBRANA GARANTISCE UNA SELETTIVITA ED AVVIENE PASSIVAMENTE: DIFFUSIONE ATTIVAMENTE: RICHIEDE ENERGIA SEMPLICE DELL ACQUA FACILITATA
Fisiologia cellulare e Laboratorio di Colture cellulari
 Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
TARGETS DEI FARMACI ATTUALMENTE IN USO
 TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
La trasmissione dell Impulso Nervoso
 La trasmissione dell Impulso Nervoso Il Neurone Neuroni: cellule specializzate per la trasmissione degli impulsi. Dendriti prolungamenti riccamente ramificati e un lungo Assone fibroso. La parte terminale
La trasmissione dell Impulso Nervoso Il Neurone Neuroni: cellule specializzate per la trasmissione degli impulsi. Dendriti prolungamenti riccamente ramificati e un lungo Assone fibroso. La parte terminale
Il sistema cardiovascolare (cenni)
") Il sistema cardiovascolare (cenni) Il sistema cardiovascolare (cenni) Il movimento del sangue nei vasi è mantenuto dal cuore, la cui funzione è quella di pompare nelle arterie (ad alta pressione), dopo
Il sistema cardiovascolare (cenni) Il sistema cardiovascolare (cenni) Il movimento del sangue nei vasi è mantenuto dal cuore, la cui funzione è quella di pompare nelle arterie (ad alta pressione), dopo
4. IL SISTEMA ENDOCRINO: GENERALITA
 4. IL SISTEMA ENDOCRINO: GENERALITA 1. Ghiandole endocrine e ormoni 2. Il tessuto bersaglio 3. Rilascio degli ormoni 4. Il concefo della regolazione retroahva negajva 5. La neuroipofisi: ossitocina e ADH
4. IL SISTEMA ENDOCRINO: GENERALITA 1. Ghiandole endocrine e ormoni 2. Il tessuto bersaglio 3. Rilascio degli ormoni 4. Il concefo della regolazione retroahva negajva 5. La neuroipofisi: ossitocina e ADH
temperatura e attività delle membrane biologiche
 intracellulare MEZZO extracellulare Interstiziale plasma Angraecum sesquipedale Thouars, 1822, nota come orchidea cometa, è una pianta appartenente alla famiglia delle Orchidaceae, nativa del Madagascar.
intracellulare MEZZO extracellulare Interstiziale plasma Angraecum sesquipedale Thouars, 1822, nota come orchidea cometa, è una pianta appartenente alla famiglia delle Orchidaceae, nativa del Madagascar.
CELLULA E MEMBRANA CELLULARE
 CELLULA E MEMBRANA CELLULARE LA CELLULA Le membrane cellulari separano la regione intracellulare da quella extracellulare 15 µm 1 La membrana cellulare Funzioni 1) ISOLAMENTO FISICO 2) REGOLAZIONE DEGLI
CELLULA E MEMBRANA CELLULARE LA CELLULA Le membrane cellulari separano la regione intracellulare da quella extracellulare 15 µm 1 La membrana cellulare Funzioni 1) ISOLAMENTO FISICO 2) REGOLAZIONE DEGLI
!"#$%&'"()*+,*#&-.(/#&!"#$%&0("1(&'"()*+,*(&23(4+(& 5+",1#)+&6%7&8&--&796:;6<&
*+,*#&-.(/#&!#$%&0(1(&'()*+,*(&23(4+(& 5+,1#)+&6%7&8&--&796:;6<&") W}švÌ]o[Ì]}v!!"#$%&'"()*+,*#&-.(/#&!"#$%&0("1(&'"()*+,*(&23(4+(& 5+",1#)+&6%7&8&--&796:;6
W}švÌ]o[Ì]}v!!"#$%&'"()*+,*#&-.(/#&!"#$%&0("1(&'"()*+,*(&23(4+(& 5+",1#)+&6%7&8&--&796:;6
FISIOLOGIA Generale e Oculare File # 3 UniSalento Ottica e Optometria Fisiologia Generale e Oculare A.A S. Marsigliante
 FISIOLOGIA Generale e Oculare File # 3 POTENZIALE ELETTRICO DELLE MEMBRANE CELLULARI Ogni membrana cellulare ha un caratteristico potenziale elettrico che va da -10 mv sino a -100 mv; per esempio: cellule
FISIOLOGIA Generale e Oculare File # 3 POTENZIALE ELETTRICO DELLE MEMBRANE CELLULARI Ogni membrana cellulare ha un caratteristico potenziale elettrico che va da -10 mv sino a -100 mv; per esempio: cellule
L eccitabilità cellulare
 L eccitabilità cellulare La membrana citoplasmatica e il modello a mosaico fluido Funzioni della membrana plasmatica: Colesterolo 1 separazione dei liquidi intra/extracellulari 2 regolazione scambi con
L eccitabilità cellulare La membrana citoplasmatica e il modello a mosaico fluido Funzioni della membrana plasmatica: Colesterolo 1 separazione dei liquidi intra/extracellulari 2 regolazione scambi con
Segnali Bioelettrici. Bioingegneria. Docente Prof. Francesco Amato. Versione 1.3 AA 2014/15.
 Segnali Bioelettrici Docente Prof. Francesco Amato Versione 1.3 AA 2014/15 amato@unicz.it 1 Interazione: Misurare, elaborare, analizzare, presentare un biosegnale Controllo e Feedback Sensore Alimentazione
Segnali Bioelettrici Docente Prof. Francesco Amato Versione 1.3 AA 2014/15 amato@unicz.it 1 Interazione: Misurare, elaborare, analizzare, presentare un biosegnale Controllo e Feedback Sensore Alimentazione
Intracellulare Extracellulare Na K Mg Ca Cl Anioni non permeabili abbondanti scarsi
 Tessuto nervoso 12.8 Un esempio di neurone Concentrazioni ioniche all interno e all esterno di una tipica cellula di mammifero Concentrazioni espresse in mmoli/litro Intracellulare Extracellulare Na +
Tessuto nervoso 12.8 Un esempio di neurone Concentrazioni ioniche all interno e all esterno di una tipica cellula di mammifero Concentrazioni espresse in mmoli/litro Intracellulare Extracellulare Na +
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA)
") Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
+... La resistenza interna (R int ) dipende dalla resistività (ρ), da l e dalla sezione secondo la relazione: int. memb
 dipende dalla resistività (ρ), da l e dalla sezione secondo la relazione: int. memb") 0.05 mm λ 10 mm 1 La costante di spazio aumenta con le dimensioni cellulari Ogni assone può essere pensato come formato da N tasselli di membrana dotati della stessa resistenza unitaria R u. Dal punto
0.05 mm λ 10 mm 1 La costante di spazio aumenta con le dimensioni cellulari Ogni assone può essere pensato come formato da N tasselli di membrana dotati della stessa resistenza unitaria R u. Dal punto
V = 130 mv R = Ω = = A I = V R. Trascuriamo il canale del Cl - Voltaggio totale V. Resistenza totale R
 LA MEMBRANA I canali passivi trans-membranici selettivi per ciascuna specie di ione possono essere schematizzati come una combinazione di resistenze e batterie Trascuriamo il canale del Cl - V = 130 mv
LA MEMBRANA I canali passivi trans-membranici selettivi per ciascuna specie di ione possono essere schematizzati come una combinazione di resistenze e batterie Trascuriamo il canale del Cl - V = 130 mv
Propagazione elettrotonica
 Un potenziale sotto soglia nato in un punto diminuisce di ampiezza man mano che è condotto lungo l assone o i dendriti di un neurone (conduzione elettrotonica). La resistenza di membrana (r m ) e dell
Un potenziale sotto soglia nato in un punto diminuisce di ampiezza man mano che è condotto lungo l assone o i dendriti di un neurone (conduzione elettrotonica). La resistenza di membrana (r m ) e dell
Cos è l eccitabilità della membrana?
 Potenziale d Azione Cos è l eccitabilità della membrana? Modificazione transitoria delle proprietà della membrana (permeabilità ionica e potenziale transmembrana) in seguito all azione di uno stimolo.
Potenziale d Azione Cos è l eccitabilità della membrana? Modificazione transitoria delle proprietà della membrana (permeabilità ionica e potenziale transmembrana) in seguito all azione di uno stimolo.
COMPARTIMENTI IDRICI AMBIENTE ESTERNO AMBIENTE INTERNO
 COMPARTIMENTI IDRICI AMBIENTE ESTERNO AMBIENTE INTERNO OMEOSTASI Stato di stabilità locale che tende a mantenersi lontano dallo stato di equilibrio EQUILIBRIO Stato di stabilità energetica, massima entropia
COMPARTIMENTI IDRICI AMBIENTE ESTERNO AMBIENTE INTERNO OMEOSTASI Stato di stabilità locale che tende a mantenersi lontano dallo stato di equilibrio EQUILIBRIO Stato di stabilità energetica, massima entropia
Il neurone. I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi.
 attraverso le sinapsi.") Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
Scaricato da 1
 POTENZIALI GRADUATI Sono depolarizzazioni o iperpolarizzazioni che si verificano nei dendriti, nel soma o, meno di frequente, vicino al terminale assonale. Sono definiti graduati perché la loro ampiezza
POTENZIALI GRADUATI Sono depolarizzazioni o iperpolarizzazioni che si verificano nei dendriti, nel soma o, meno di frequente, vicino al terminale assonale. Sono definiti graduati perché la loro ampiezza
POTENZIALE D AZIONE FGE AA
 POTENZIALE D AZIONE FGE AA.2015-16 Neurone Soma Nucleo Assone Terminazioni sinaptiche Dendriti Guaina mielinica Neurone Sezione istologica Neurone 1 Il neurone 1 comunica con il neurone 2 come mostrato
POTENZIALE D AZIONE FGE AA.2015-16 Neurone Soma Nucleo Assone Terminazioni sinaptiche Dendriti Guaina mielinica Neurone Sezione istologica Neurone 1 Il neurone 1 comunica con il neurone 2 come mostrato
Compito A-7 BIOFISICA E FISIOLOGIA I Anno Accademico
 Compito A-7 1) L ampiezza del potenziale d azione che circola lungo un assone mielinico è 115 mv. Sapendo che, a riposo, il rapporto p Na /p K è 0,015, calcolare di quanto varia p Na nel corso di un potenziale
Compito A-7 1) L ampiezza del potenziale d azione che circola lungo un assone mielinico è 115 mv. Sapendo che, a riposo, il rapporto p Na /p K è 0,015, calcolare di quanto varia p Na nel corso di un potenziale
Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari
 Corso di laurea magistrale in BIOTECNOLOGIE DELLA RIPRODUZIONE UNIVERSITA DEGLI STUDI DI TERAMO Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari Prof.ssa Luisa Gioia Costituenti
Corso di laurea magistrale in BIOTECNOLOGIE DELLA RIPRODUZIONE UNIVERSITA DEGLI STUDI DI TERAMO Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari Prof.ssa Luisa Gioia Costituenti
Corso di Fisica Medica 1
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Conduzione nervosa 30/3/2006 Terminazioni pre-sinaptiche Mitocondri Sintesi dei neuro trasmettitori (NT) Attività enzimatica Trasporto
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Conduzione nervosa 30/3/2006 Terminazioni pre-sinaptiche Mitocondri Sintesi dei neuro trasmettitori (NT) Attività enzimatica Trasporto
Attivi (non equilibranti) Passivi (equilibranti) Migrazione attraverso canali membran. Diffusione semplice. Tasporto attivo secondario
 Passivi (equilibranti) Migrazione attraverso canali membran. Diffusione semplice. Tasporto attivo secondario") TRASPORTI Trasporti in forma libera Trasporti mediati Diffusione semplice Migrazione attraverso canali membran. Diffusione facilitata Trasporto attivo primario Tasporto attivo secondario Passivi (equilibranti)
TRASPORTI Trasporti in forma libera Trasporti mediati Diffusione semplice Migrazione attraverso canali membran. Diffusione facilitata Trasporto attivo primario Tasporto attivo secondario Passivi (equilibranti)
Caratteristiche delle membrane biologiche
 Membrane Biologiche Caratteristiche delle membrane biologiche 1. Le membrane sono strutture a foglio che formano setti di separazione tra compartimenti con composizione diversa tra loro. 2. Sono costituite
Membrane Biologiche Caratteristiche delle membrane biologiche 1. Le membrane sono strutture a foglio che formano setti di separazione tra compartimenti con composizione diversa tra loro. 2. Sono costituite
Elettricità cellulare
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Elettricità cellulare 30/3/2006 Sistemi biologici Essenzialmente costituiti da acqua (solvente) e da differenti soluti (molti dei
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Elettricità cellulare 30/3/2006 Sistemi biologici Essenzialmente costituiti da acqua (solvente) e da differenti soluti (molti dei
Trasporti attraverso. Proteine di trasporto
 Trasporti attraverso la membrana Proteine di trasporto Diffusione Facilitata Caratteristiche dei trasporti mediati I carriers sono dotati di specificità Sono soggetti a saturazione Possono essere bloccati
Trasporti attraverso la membrana Proteine di trasporto Diffusione Facilitata Caratteristiche dei trasporti mediati I carriers sono dotati di specificità Sono soggetti a saturazione Possono essere bloccati
5. Trasporto di membrana
 5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
Lezione 16. Elettrodinamica
 Lezione 16 Elettrodinamica Introduzione Nei conduttori solidi qualche elettrone per atomo può diventare libero di muoversi passando da un atomo all'altro. Applicando la teoria cinetica dei gas si trova
Lezione 16 Elettrodinamica Introduzione Nei conduttori solidi qualche elettrone per atomo può diventare libero di muoversi passando da un atomo all'altro. Applicando la teoria cinetica dei gas si trova
COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA
 COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA.2015-16 MECCANISMI DI TRASPORTO Trasporto transmembranario Le membrane biologiche sono barriere di permeabilità che impediscono a gran parte delle molecole
COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA.2015-16 MECCANISMI DI TRASPORTO Trasporto transmembranario Le membrane biologiche sono barriere di permeabilità che impediscono a gran parte delle molecole
dq dt Università degli Studi di Bari Aldo Moro Dip. DiSAAT - Ing. Francesco Santoro Corso di Fisica
 Corrente elettrica Consideriamo il moto non accelerato e con velocità piccole rispetto a quella della luce nel vuoto di un insieme di particelle dotate di carica elettrica: possono ritenersi valide le
Corrente elettrica Consideriamo il moto non accelerato e con velocità piccole rispetto a quella della luce nel vuoto di un insieme di particelle dotate di carica elettrica: possono ritenersi valide le
Differenti configurazioni di proteine-canale
 Proteine-canale Le proteine-canale formano nella membrana dei «pori» attraverso i quali transitano a grande velocità gli ioni inorganici e le molecole d acqua. Pertanto, il trasporto mediato da proteine
Proteine-canale Le proteine-canale formano nella membrana dei «pori» attraverso i quali transitano a grande velocità gli ioni inorganici e le molecole d acqua. Pertanto, il trasporto mediato da proteine
BIOFISICA DELLE MEMBRANE
 BIOFISICA DELLE MEMBRANE Le funzioni biologiche di tutti gli organismi viventi si svolgono mediante scambio di sostanze ed informazioni attraverso membrane. Membrana = struttura che separa due mezzi diversi
BIOFISICA DELLE MEMBRANE Le funzioni biologiche di tutti gli organismi viventi si svolgono mediante scambio di sostanze ed informazioni attraverso membrane. Membrana = struttura che separa due mezzi diversi
La membrana cellulare
 La membrana cellulare Membrane artificiali Modello di membrana cellulare Criodecappaggio e congelamento-incisione Le membrane sono fluide I lipidi si muovono lateralmente Presenza di code idrocarburiche
La membrana cellulare Membrane artificiali Modello di membrana cellulare Criodecappaggio e congelamento-incisione Le membrane sono fluide I lipidi si muovono lateralmente Presenza di code idrocarburiche
Neurone Biologico. Paolo Montalto Alessandro Rea Pietro Sassone Maurizio Triunfo
 Neurone Biologico Paolo Montalto Alessandro Rea Pietro Sassone Maurizio Triunfo Il Sistema Nervoso: concetti introduttivi Il sistema nervoso è la sede delle facoltà sensoriali e del pensiero e provvede
Neurone Biologico Paolo Montalto Alessandro Rea Pietro Sassone Maurizio Triunfo Il Sistema Nervoso: concetti introduttivi Il sistema nervoso è la sede delle facoltà sensoriali e del pensiero e provvede
Compito di prova - risolti
 Compito di prova - risolti A P B q A q P q B 1. La carica positiva mobile q P si trova tra le cariche positive fisse q A, q B dove AB = 1 m. Se q A = 2 C e all equilibrio AP = 0.333 m, la carica q B vale
Compito di prova - risolti A P B q A q P q B 1. La carica positiva mobile q P si trova tra le cariche positive fisse q A, q B dove AB = 1 m. Se q A = 2 C e all equilibrio AP = 0.333 m, la carica q B vale
Principali differenze tra sinapsi neuromuscolare e sinapsi centrali
 Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Biosensori Sensori Chimici. Alessandro Tognetti
 Biosensori Sensori Chimici Alessandro Tognetti Principali applicazioni dei Sensori chimici Ruolo fondamentale degli ioni nella maggior parte dei processi biologici Sensori elettrochimici per la misura
Biosensori Sensori Chimici Alessandro Tognetti Principali applicazioni dei Sensori chimici Ruolo fondamentale degli ioni nella maggior parte dei processi biologici Sensori elettrochimici per la misura
Potenziale di Membrana
 Potenziale di Membrana Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo della membrana plasmatica, infatti una cellula con un potenziale
Potenziale di Membrana Genesi del Potenziale di Membrana In tutte le cellule è possibile misurare una differenza di potenziale a cavallo della membrana plasmatica, infatti una cellula con un potenziale
Dati numerici: f = 200 V, R 1 = R 3 = 100 Ω, R 2 = 500 Ω, C = 1 µf.
 ESERCIZI 1) Due sfere conduttrici di raggio R 1 = 10 3 m e R 2 = 2 10 3 m sono distanti r >> R 1, R 2 e contengono rispettivamente cariche Q 1 = 10 8 C e Q 2 = 3 10 8 C. Le sfere vengono quindi poste in
ESERCIZI 1) Due sfere conduttrici di raggio R 1 = 10 3 m e R 2 = 2 10 3 m sono distanti r >> R 1, R 2 e contengono rispettivamente cariche Q 1 = 10 8 C e Q 2 = 3 10 8 C. Le sfere vengono quindi poste in
Elettrochimica. le trasformazioni redox spontanee (DG < 0) l energia elettrica in celle elettrolitiche per ottenere
 l energia elettrica in celle elettrolitiche per ottenere") Elettrochimica studia le relazioni tra processi chimici ed energia elettrica. i. e. si interessa dei processi che coinvolgono il trasferimento di elettroni (reazioni di ossido riduzione). Sfrutta: le trasformazioni
Elettrochimica studia le relazioni tra processi chimici ed energia elettrica. i. e. si interessa dei processi che coinvolgono il trasferimento di elettroni (reazioni di ossido riduzione). Sfrutta: le trasformazioni
RELAZIONE SULLO STAGE AL DIFI SEZIONE DI BIOFISICA
 Autera Simone De Martini Valentina Liggieri Giovanni RELAZIONE SULLO STAGE AL DIFI SEZIONE DI BIOFISICA TUTORS: Francesca Pellistri, Silvia Siano, Andrea Gallo 27/02-03/03/ 2006 Lo stage al DIFI (Dipartimento
Autera Simone De Martini Valentina Liggieri Giovanni RELAZIONE SULLO STAGE AL DIFI SEZIONE DI BIOFISICA TUTORS: Francesca Pellistri, Silvia Siano, Andrea Gallo 27/02-03/03/ 2006 Lo stage al DIFI (Dipartimento
giunzioni comunicanti connessone cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali precoci veloci bidirezionali
 La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
IL SISTEMA NERVOSO. Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO. Sensoriale (dall'esterno)
 Trasmissivo (Cablaggio) SISTEMA NERVOSO. Sensoriale (dall'esterno)") IL SISTEMA NERVOSO Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO Periferico Afferente (dalla periferia verso l'unità centrale) Sensoriale (dall'esterno) Autonomo
IL SISTEMA NERVOSO Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO Periferico Afferente (dalla periferia verso l'unità centrale) Sensoriale (dall'esterno) Autonomo
Per una mole di AB 1 - α α α. Per n moli di AB n (1 α) n α n α. Il numero totale delle particelle, all'equilibrio, sono:
 n α n α. Il numero totale delle particelle, all'equilibrio, sono:") Elettroliti AB A + + B - Grado di dissociazione α è la frazione di mole che all equilibrio ha subito dissociazione α : 1 = n mol dissociate : n mol iniziali AB A + + B - Per una mole di AB 1 - α α α Per
Elettroliti AB A + + B - Grado di dissociazione α è la frazione di mole che all equilibrio ha subito dissociazione α : 1 = n mol dissociate : n mol iniziali AB A + + B - Per una mole di AB 1 - α α α Per