LA GENETICA QUANTITATIVA
|
|
|
- Rossana Arena
- 7 anni fa
- Visualizzazioni
Transcript
1 Genetica 5
2 LA GENETICA QUANTITATIVA
3 Caratteri quantitativi vs. Caratteri qualitativi
4 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

5 controversia Mendel/cararatteri continui Mendel aveva lavorato su caratteri discontinui, per cui molti dubitavano che le sue leggi valessero anche per i caratteri quantitativi, quali l altezza, che si sapevano essere ereditari ma non se ne conoscevano i meccanismi di trasmissione. Nel 1918 Fisher provò che l ereditarietà delle caratteristiche quantitative poteva realmente venire spiegata attraverso gli effetti cumulativi di molti geni, ciascuno dei quali seguiva le leggi di Mendel

6 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
7 Ereditarietà e variabilità dei caratteri quantitativi > caratteri che variano in modo continuo nella popolazione e che per tale motivo possono essere misurati es. lunghezza della spiga, peso del seme, dimensione corporea di un animale, altezza di un individuo, ecc. > caratteri poligenici poichè sono controllati da molti geni con effetto limitato ma aventi azione uguale e cumulativa sul valore fenotipico; > caratteri la cui manifestazione fenotipica può essere influenzata da fattori ambientali.
8 Caratteri multifattoriali: poligeni e ambiente a) Carattere Poligenico: per l espressione di un determinato carattere continuo (varietà di fenotipi) concorrono soltanto più geni Ma non solo genotipi multipli. b) Carattere Multifattoriale: per l espressione di un determinato carattere continuo concorrono più geni e fattori ambientali
 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005")
9 Fattori multigenici + norma di reazione (NdR: gamma di fenotipi prodotti da un genotipo in vari ambienti) Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
10 Analisi dei caratteri quantitativi a) Dividere le variazioni osservate nel carattere nelle due componenti genetica ambientale b) Utilizzare la componente genetica per la previsione dei fenotipi della progenie
11 Campioni e popolazioni Affinchè il campione descriva la popolazione, esso deve essere: Rappresentativo Sufficientemente ampio Rappresentativo: una squadra di pallacanestro non rappresenta tutte le altezze di una popolazione Ampio: se misurassimo altezza di soli 3 studenti dell ateneo di VR e questi causalmente fossero tutti e 3 bassi, sottostimeremmo l altezza media della popolazione studentesca di VR
12 Metodi statistici per l analisi delle caratteristiche quantitative All interno di un gruppo, la variabilità fenotipica può essere rappresentata per mezzo di una distribuzione di frequenza: grafico di frequenze relative ai diversi fenotipi Asse X riporta le classi fenotipiche. L asse Y il numero o la percentuale. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
 asimmetrica, C) bimodale Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005")
13 Distribuzione normale, asimmetrica e bimodale Quando la curva è simmetrica, la distribuzione è normale. B) asimmetrica, C) bimodale Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
14 La media La media fornisce informazioni sul centro di una distribuzione Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 x = n x i Misuriamo altezza ragazzi di 10 e di 18 anni. Entrambe le distribuzioni sono normali, ma centrate su valori di statura differenti. FORMULA: la media si calcola sommando le singole misurazioni e dividendo per il numero di misurazioni

15 La varianza (s 2 ) La varianza è definita come la deviazione quadratica media dalla media. Essa indica il grado di variabilità di un gruppo di fenotipi (misurazioni). Quanto maggiore è la varianza, tanto più dispersi sono i valori di una distribuzione intorno alla media Per calcolare la varianza si sottrae la media da ogni misurazione, portando al quadrato il valore ottenuto. Si sommano poi i valori e il risultato lo si divide per il numero di misurazioni di partenza meno 1 S 2 = (x i x) 2 n - 1

16 La deviazione standard (s) La deviazione standard è la radice quadrata della varianza. Essa misura la dispersione dei dati intorno al valore atteso Si esprime nelle stesse unità della misura originale, pertanto descrive la variabilità di una misura s = s 2 La media più o meno una deviazione standard comprende il 66% circa delle misurazioni relative ad una distribuzione normale. Di conseguenza, solo l 1% di una popolazione con distribuzione normale rimane fuori dall intervallo media +- 3s Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
17 Coefficiente di correlazione (r) Il coefficiente di correlazione stabilisce la forza dell associazione fra due caratteristiche. Es. altezza (x) e peso di una persona (y) Accade sovente che due o più caratteri varino contemporaneamente. Per esempio, negli allevamenti di polli sono importanti sia il numero sia la dimensione delle uova prodotte. Queste due caratteristiche non sono tra di loro indipendenti, bensi legate da una correlazione inversa
18 Coefficiente di correlazione (r) r = cov xy s x s y Il coeff. di correlazione si ottiene dividendo la covarianza di x e y per il prodotto delle deviazioni standard di x e y. Esso spazia da +1 a -1. Un valore positivo indica l esistenza di correlazione positiva fra le due variabili cov xy = (x i x) (y i y) n - 1 La covarianza è un indice che misura la "contemporaneità" della variazione (in termini lineari) di due variabili casuali. In pratica, la covarianza di due variabili aleatorie X e Y è il valore atteso dei prodotti delle loro distanze dalla media Essa può assumere sia valori positivi che negativi. Nel caso di valori positivi indica che al crescere di una caratteristica statisticamente cresce anche l'altra, nel caso di valori negativi accade il contrario
19 altezza e peso numero uova e peso uova altezza e intelligenza Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
20 r puo essere calcolato anche per una sola variabile misurata per coppie di individui Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
 a = intercetta di y, ovvero il valore di y quando")
21 Coefficiente di regressione (b) Indica l entità dell aumento di y all aumentare di x. Pertanto, b consente di prevedere il valore di una variabile, dato il valore dell altra b = cov xy s x 2 la curva di regressione è ( y = a + bx ) a = intercetta di y, ovvero il valore di y quando x=0, b = pendenza della curva, ovvero l aumento medio di y all aumentare di x Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
22 Il coefficiente di regressione consente di prevedere determinate caratteristiche della progenie generata da una data unione anche senza conoscere i genotipi che codificano per tale carattere Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
23 Influenza dell ambiente sulla variabilità dei caratteri quantitativi Wilhelm Johannsen ( ): primo studioso a mettere in evidenza l azione congiunta dei fattori genetici e dei fattori ambientali nell eredità dei caratteri quantitativi egli realizzò una serie di esperimenti utilizzando semi di fagiolo della varietà commerciale Princess, sapendo che la specie scelta è strettamente autogama e che ciascun seme era da ritenersi omozigote a tutti i loci
24 Influenza dell ambiente sulla variabilità dei caratteri quantitativi la variabilità fenotipica di una carattere quantitativo può avere due componenti: una genetica ed un altra ambientale; la selezione è efficace solo in presenza di variabilità genetica; la variabilità che si osserva entro linee pure è dovuta unicamente all ambiente; la selezione entro linea pura è del tutto inefficace.
25 Esperimenti di W. Johannsen in fagiolo: Influenza dell ambiente sull espressione dei caratteri quantitativi Johannsen valutando il peso dei semi di fagiolo è stato il primo a distinguere la variabilità genetica dalla variabilità ambientale e a dimostrare che la selezione può operare solo sulla variabilità genetica. Le 19 linee estratte dalla varietà commerciale erano infatti differenti: le differenze tra le medie delle linee in termini di peso del seme dovevano essere di natura genetica. La selezione entro linee risultava invece inefficace dimostrando che la variabilita del peso del seme entro linea dipendeva soltanto da fattori ambientali.
26 Influenza dei fattori genetici sulla variabilità dei caratteri quantitativi Herman Nilsson-Ehle (1908): è stato il primo ricercatore a trovare un modello naturale in grado di spiegare l eredità dei caratteri quantitativi. egli studiò il colore della cariosside in frumento e formulò l ipotesi che più coppie alleliche segreganti in maniera indipendente, ereditate in assenza di dominanza ed aventi azione uguale e cumulativa (additiva) potessero spiegare i risultati relativi al grado di espressione del carattere nella generazione F 2.
27 Eredità del colore della cariosside in frumento P Colore delle cariossidi A A B B x A 1 A 1 B 1 B1 Bianco Rosso molto scuro Grado di colore 0 4 F 1 AB (¼) A 1 B (¼) AB 1 (¼) A 1 B 1 (¼) A A 1 B B1 Rosso intermedio 2 AB (¼) A 1 B (¼) AB 1 (¼) A 1 B 1 (¼) AABB AA 1 BB AABB 1 AA 1 BB 1 AA 1 BB A 1 A 1 BB AA 1 BB 1 A 1 A 1 BB 1 AABB 1 AA 1 BB 1 AAB 1 B 1 AA 1 B 1 B 1 AA 1 BB 1 A 1 A 1 BB 1 AA 1 B 1 B 1 A 1 A 1 B 1 B 1 F 2 Rapporto 15:1 piante con cariossidi colorate (ma di intensità variabile: chiaro, medio, scuro e molto scuro) e piante con cariossidi bianche
28
29 Influenza dei fattori genetici sulla variabilità dei caratteri quantitativi Se due coppie alleliche vengano ereditate in maniera indipendente, in assenza di dominanza e gli alleli plus hanno azione uguale e cumulativa nel dare la colorazione: l intensità del colore è data dal numero di alleli A 1 e B 1 che nel genotipo delle diverse piante F 2 agiscono nello stesso senso in maniera cumulativa AB 1 A 1 B AB A 1 B 1 AB A 1 B AB 1 A 1 B 1 Azione di fattori A 1 B 1 per il colore e di alleli A e B per il non colorato geni plus geni minus Alleli minus Alleli che esercitano un effetto minimo o che sono ininfluenti sulla manifestazione fenotipica di un carattere quantitativo. Alleli plus Alleli in grado di contribuire alla manifestazione di un carattere quantitativo agendo nello stesso senso in maniera cumulativa e con effetto sul fenotipo superiore alla media.
30 Assunzioni dell ipotesi multigenica di Nilsson-Ehle: in nessuno dei loci uno degli alleli presenta dominanza sull altro, piuttosto risulta coinvolta una serie di alleli con effetto additivo; ogni allele plus agisce nello stesso senso in maniera cumulativa ed ha uguale effetto sul fenotipo; gli alleli minus non contribuiscono (o contribuiscono in maniera minima) al fenotipo; non esiste interazione genica (epistasi) tra loci differenti di una serie; i loci non sono associati, cioè gli alleli segregano in maniera indipendente. Questo lavoro ha dimostrato che la differenza fra eredità di caratteri continui e discontinui risiede nel numero di loci che determinano quel carattere
31 Determinazione numero di geni per caratteristica poligenica Vengono incrociati due individui omozigoti per alleli differenti (A1 e A) di un singolo locus (A) A 1 A 1 x AA A 1 A Gli individui della risultante F1 sono incrociati fra di loro A 1 A x A 1 A ¼ della F2 risulterà omozigote come i parentali. Se ci sono 2 loci coinvolti, ¼ x ¼ = 1/16 A 1 A 1 A 1 A AA 1 AA ¼ 2/4 ¼ Pertanto, in F2 il numero di individui simili a ciascuno dei parentali (n indica numero di loci) e : 1 4 n se trovo: 1/4 individui, significa che c e un locus 1/16 individui, significa che ci sono 2 loci 1/256 individui, significa che ci sono 4 loci Questo metodo assume che tutti i geni che influenzano la caratteristica siano additivi, presentino effetti equivalenti e non siano associati.

32 Determinazione numero di geni per caratteristica poligenica Edward East accoppiò due varietà di tabacco omozigoti per la lunghezza del fiore: 40,5 mm e 93,3 mm, rispettivamente. Egli trovò che nella F 1 la lunghezza del fiore era intermedia, con una varianza simile a quella riscontrata nei parentali. East accoppiò poi fra di loro gli individui F 1. Nella F 2 la lunghezza media del fiore era simile a quella riscontrata nella F 1, ma la varianza era molto superiore, ad indicare la presenza di differenti genotipi. Dato che nessuna delle 444 piante F 2 mostrava lunghezza del fiore analoga ai parentali, il carattere doveva essere codificato da più di 4 loci Successivamente, East selezionò alcune piante F 2 e le incrociò: nella F 3 l estensione del fiore dipendeva dalla lunghezza del fiore nei parentali. Questo indicava l esistenza di differenze sul piano genetico nell ambito della F 2. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

33 Localizzazione dei geni che influiscono sulle caratteristiche quantitative QTL (quantitative trait loci) sono i geni che controllano le caratteristiche poligeniche Si identificano marcatori molecolari che segregano nella popolazione assieme ad una porzione del carattere quantitativo Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

34 Quantitative Trait Locus (QTL) Un QTL è una regione di DNA associata ad un particolare carattere quantitativo. Il QTL è strettamente associato ai geni che determinano il carattere fenotipico in questione o partecipa nella sua determinazione. Normalmente infatti un carattere quantitativo è determinato dalla somma dell'azione di più geni (un fenomeno detto additività). Di conseguenza più QTL, che possono trovarsi anche su diversi cromosomi, sono associati ad un singolo carattere. Il numero di QTL coinvolti in un carattere fornisce informazioni sull'architettura genetica del carattere; per esempio indica se l'altezza di una pianta è determinata da molti geni, l'effetto di ognuno dei quali è di portata limitata, oppure invece da pochi geni con un effetto più marcato.
35 Ereditabilità di un carattere (H 2 ) Frazione della variazione fenotipica causata dalle differenze genetiche Osservabile a livello di popolazione, non di singolo individuo! Oltre ad essere poligeniche, le caratteristiche quantitative sono spesso influenzate da fattori ambientali. E quindi utile sapere quanto la variabilità di un carattere quantitativo sia dovuta a differenze genetiche e quanto a differenze ambientali. La quota di variazione fenotipica complessiva derivante da fattori genetici è nota come ereditabilità
36 Ereditabilità di un carattere (H 2 ) Esprime la proporzione della variabilità fenotipica (V P ) imputabile a differenze genetiche (V G ) Il contributo relativo dei fattori genetici e dei fattori ambientali nella determinazione della variabilità di un carattere quantitativo è misurato da un parametro che prende il nome di ereditabilità. Essa consente di prevedere il fenotipo della progenie in base al fenotipo parentale EREDITABILITA : H 2 = V G V P frazione ereditabile della variazione di un carattere quantitativo H 2 = 0-1 Per determinare l ereditabilità di un carattere quantitativo occorre per prima cosa misurarne la variabilità - varianza (misura la fluttuazione dei dati individuali intorno alla media del campione ed equivale allo scarto quadratico medio) - e quindi suddividere questa nelle sue componenti.
37 COMPONENTI DELLA VARIANZA FENOTIPICA (V P ) Alcune differenze fenotipiche sono dovute a fattori genetici e a fattori ambientali. VARIANZA GENETICA = V G (differenze genetiche tra gli individui) VARIANZA AMBIENTALE = V E (influenza delle condizioni ambientali) V P =V G +V E
38 Esempio di interazione genotipo ambiente. Il genotipo influisce sul peso della pianta, ma sono le condizioni climatiche a determinare quale genotipo produca l esemplare piu pesante! Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
39 Una certa quota di variazione genetica è ereditabile poiché passa dai genitori ai figli. Un altra quota di variazione è genetica ma non strettamente ereditabile a causa di fenomeni quali la dominanza e le interazioni epistatiche che non passano direttamente dai genitori ai figli. Per esempio, se un allele è dominante su un altro, il fenotipo di un genitore eterozigote è determinato in parte dalla interazione di dominanza fra i due alleli. Un genitore che si riproduce sessualmente passerà alla sua prole soltanto un allele. La prole, infatti, eredita il genotipo da entrambi i genitori e quindi non erediterà l interazione di dominanza, ma soltanto l effetto di un singolo allele
40 COMPONENTI DELLA VARIANZA GENETICA (V G ) La variazione genetica può quindi essere ulteriormente suddivisa in tre componenti: 1. variazione genetica additiva: alcuni alleli possono contribuire con un valore fisso al valore metrico di un carattere quantitativo. Tali geni si definiscono additivi e contribuiscono alla varianza genetica additiva (V A ). 2. variazione genetica dominante: alcuni alleli sono dominanti su altri e mascherano il contributo degli alleli recessivi in quel locus. Questa fonte di variabilità contribuisce alla varianza genetica da dominanza (V D ). 3. variazione genetica causata dalle interazioni fra geni diversi, dovuta fondamentalmente a fenomeni di epistasi. Tale variazione contribuisce alla varianza genetica da interazione (V I ) V G = V A + V D + V I
41 COMPONENTI DELLA VARIANZA GENETICA (V G ) VARIANZA ADDITIVA = V A VARIANZA dovuta alla DOMINANZA = V D VARIANZA EPISTATICA = V I V G = V A + V D + V I e la varianza fenotipica totale può essere riscritta come: V P = V E + V A + V D + V I
42 EREDITABILITA Il termine generale che descrive la proporzione della varianza genetica alla varianza totale è l ereditabilità. Si possono stimare due tipi specifici di ereditabilità. In senso lato l ereditabilità è il rapporto fra la varianza genetica totale (incluse dominanza ed epistasi) e la varianza fenotipica totale, cioè: H 2 = V G /V P In senso stretto l ereditabilità è il rapporto fra la varianza genetica additiva e la varianza fenotipica totale, cioè: h 2 varia tra 0 (V A =0) e 1(V A =V P ) h 2 = V A /V P Per predire il fenotipo della prole proveniente da un particolare accoppiamento è importante conoscere la porzione di varianza genetica additiva, poiché in questo modo sarà possibile conoscere il contributo relativo dei genitori alla costituzione della F1.
43 EREDITABILITA IN SENSO LATO E la frazione di varianza totale che rimane dopo esclusione della frazione dovuta agli effetti ambientali. Una misura del grado di determinazione genetica di un carattere. H 2 = V G V P = V G V G + V E
44 EREDITABILITA IN SENSO STRETTO E il rapporto fra varianza genetica additiva e varianza totale. Una stima dell'efficienza della selezione. h 2 = V A V P = V A V G + V E Solo le differenze genetiche attribuibili alle azioni geniche additive possono essere fissate con la selezione: Essendo legate all effetto medio dei geni, le azioni additive rimangono inalterate nelle generazioni successive; quando invece le differenze tra i materiali selezionati sono dovute a specifiche interazioni geniche, come dominanza ed epistasi, queste non possono essere fissate con la selezione, poichè per effetto della segregazione e della ricombinazione possono ottenersi combinazioni di geni diverse da quelle della generazione precedente.
45 Calcolo dell ereditabilità in senso lato mediante eliminazione di componenti della varianza V P =V G +V E possiamo annullare V E facendo sviluppare gli individui nello stesso ambiente -> impossibile possiamo annullare V G facendo sviluppare individui geneticamente identici se V E = 0, V P = V G se V G = 0, V P = V E V G (degli individui variabili geneticamente) = V p (variabili) V E (identici) H 2 = V G V p Facciamo un esperimento in cui alleviamo individui geneticamente identici in un determinato ambiente e calcoliamo la V P, che in questo caso corrisponde alla V E. Poi prendiamo individui geneticamente diversi e calcoliamo la V P. La V G ora la posso calcolare perche ho la p e la e
46 Calcolo dell ereditabilità in senso lato mediante eliminazione di componenti della varianza: esempio Ereditabilità della chiazzatura bianca nelle cavie nella popolazione variabile geneticamente V P =573 nella popolazione dopo numerosi incroci (omozigote) V P =340 Poichè in quest ultimo gruppo V G =0, V P = V E. Questo valore di V E puo essere assegnato anche alla popolazione di partenza (geneticamente variabile) V P V E = V G = 233 H 2 = V G V p 233 H 2 = 0, % della variazione nella chiazzatura delle cavie era dovuto a differenze riguardanti il genotipo
47 Problemi nel calcolo dell ereditabilità mediante eliminazione di componenti della varianza Si assume che la varianza ambientale degli individui geneticamente identici sia equivalente a quella degli individui variabili geneticamente, e ciò può non essere vero Approccio applicabile solo quando è possibile ottenere individui geneticamente identici
48 Ereditabilità mediante regressione tra individui parentali e prole Si confrontano fenotipi di individui che presentano gradi di parentela diversi. Se la varianza fenotipica è determinata da differenze genetiche: i gemelli monozigotici devono essere simili per un determinato carattere più dei gemelli dizigotici la progenie deve risultare simile ai genitori in misura maggiore a quanto avviene negli individui non imparentati In una regressione del fenotipo medio della prole rispetto ai genitori, l ereditabilità in senso stretto (h 2 ) è uguale al coefficiente di regressione (b). Pertanto, il coefficiente di regressione fornisce informazioni sul grado dell ereditablità
 in (b) il fenotipo della prole è equivalente a quelli parentali in (c) sia i geni sia l ambiente influiscono sul fenotipo Ogni punto rappresenta un")
49 Ereditabilità mediante regressione tra individui parentali e prole Il coefficiente di regressione fornisce informazioni sul grado dell ereditablità in (a) non c e alcuna relazione fra fenotipo parentale e della prole (raggruppati in famiglie) in (b) il fenotipo della prole è equivalente a quelli parentali in (c) sia i geni sia l ambiente influiscono sul fenotipo Ogni punto rappresenta un nucleo famigliare, dove sull asse delle X c è il valore fenotipico medio dei parentali di quella famiglia, e sull asse delle y il valore fenotipico medio della prole di quella famiglia. Se per un dato tratto non c è ereditabilità in senso stretto (h2=0) i punti si disperderanno in maniera casuale e il conseguente coefficiente di regressione sarà = 0 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
50 Limitazioni dell ereditabilita Non dice quanto l espressione di un carattere dipenda da fattori genetici, ma esprime solo la proporzione della varianza fenotipica tra gli individui di una popolazione che è dovuta a differenze genetiche. L ereditabilità pertanto indica il grado a cui i geni stabiliscono la variazione di un tratto; Non si riferisce ad un individuo. E una caratteristica di una popolazione (un individuo non presenta ereditabilità!); Non esiste ereditabilità universale per un dato carattere. L ereditabilità è specifica per una data popolazione in uno specifico ambiente; Non può venire usata per trarre conclusioni riguardo la natura di differenze genetiche tra popolazioni, appunto perché è specifica per un data popolazione in un dato ambiente
51 Riepilogo La varianza fenotipica è uguale alla somma della varianza dovuta a differenze genetiche e ambientali Ereditabilità in senso lato è la proporzione di varianza fenotipica dovuta alle differenze genetiche nella popolazione Ereditabilità in senso stretto è la proporzione di varianza fenotipica dovuta agli effetti additivi degli alleli nella popolazione
52 LA GENETICA EVOLUZIONISTICA
53 Evoluzione Alla fine del 1700 la teoria predominante era quella di Linneo, detta della fissità, che definiva le varie specie come entità create una volta per tutte e incapaci di modificarsi o capaci entro ben determinati limiti. All'inizio del XIX secolo iniziarono a sorgere, negli studiosi di Scienze Naturali i primi dubbi concreti: negli strati rocciosi più antichi infatti mancano totalmente tracce (fossili) degli esseri attualmente viventi e se ne rinvengono altre appartenenti ad organismi attualmente non esistenti. Nel 1809, il naturalista Lamarck presentò per primo una teoria evoluzionista (detta lamarckismo) secondo cui gli organismi viventi si modificherebbero gradualmente nel tempo adattandosi all'ambiente: l'uso o il non uso di determinati organi porterebbe con il tempo ad un loro potenziamento o ad un'atrofia. Tale ipotesi implica quello che oggi viene considerato l'errore di fondo: l'ereditabilità dei caratteri acquisiti (esempio: un culturista non avrà necessariamente figli muscolosi; la muscolosità del culturista è infatti una manifestazione fenotipica, cioè morfologica, derivante dall'interazione dello sportivo con l'ambiente, il continuo sollevare pesi; ma il particolare sviluppo muscolare non è dettato dal suo patrimonio genetico, il genotipo).
54 Lamarck trovò opposizione in Georges L. Chretien Cuvier, il quale aveva elaborato la 'teoria delle catastrofi naturali' secondo la quale la maggior parte degli organismi viventi nel passato sarebbero stati spazzati via da numerosi cataclismi e il mondo infatti sarebbe stato ripopolato dalle specie sopravvissute. Dopo cinquant'anni Darwin formulò una nuova teoria evoluzionista; il noto naturalista, durante il suo viaggio giovanile sul brigantino Beagle, fu colpito dalla variabilità delle forme viventi che aveva avuto modo di osservare nei loro ambienti naturali intorno al mondo. Riflettendo sugli appunti di viaggio e traendo spunto dagli scritti dell'economista Thomas Malthus, Darwin si convinse che la lotta per la vita fosse uno dei motori principali dell'evoluzione intuendo il ruolo selettivo dell'ambiente sulle specie viventi. L'ambiente, infatti, non può essere la causa primaria nel processo di evoluzione (come invece sostenuto nella teoria di Lamarck) in quanto tale ruolo è giocato dalle mutazioni genetiche, in gran parte casuali. L'ambiente entra in azione in un secondo momento, nella determinazione del vantaggio o svantaggio riproduttivo che quelle mutazioni danno alla specie mutata, in poche parole, al loro migliore o peggiore adattamento (fitness in inglese). I principali meccanismi che partecipano in queste situazioni sono: * meccanismi genetici * meccanismi ecologici


55 Specie Un insieme di popolazioni formate da individui interfecondi, la cui prole a sua volta è interfeconda: pool genico chiuso
56 Evoluzione E il cambiamento della variabilità in una popolazione valutato nel tempo Le forze dell Evoluzione: Mutazione Selezione Deriva genetica: Isolamento geografico e migrazioni L evoluzione di una nuova specie da una specie ancestrale avviene sulla base della selezione naturale. Le variazioni casuali favorevoli tendono ad essere conservate, quelle non favorevoli tendono ad essere eliminate dalla pressione ambientale: adattamento all ambiente che cambia (modificazione evolutiva che aumenta la probabilità di sopravivenza di un organismo e il suo successo evolutivo viene selezionata).
57 Filogenesi E il percorso evolutivo che ha portato alla differenziazione delle specie a partire da un origine comune. La sua ricostruzione è fondamentale per la sistematica che si occupa di ricostruire le relazioni di parentela evolutiva, di gruppi tassonomici di organismi a qualunque livello sistematico La filogenetica studia l'origine e l'evoluzione di un insieme di organismi, solitamente di una specie. Un compito essenziale della sistematica è di determinare le relazioni ancestrali fra specie note (vive ed estinte). Rapporti filogenetici: i rapporti di discendenza comune tra specie o gruppi di ordine superiore
58 La selezione naturale Insieme dei fattori che determinano la sopravvivenza di alcuni organismi rispetto ad altri, all interno di una data popolazione. Essa determina l evoluzione della specie, favorendo gli organismi dotati dei caratteri più adatti in un determinato ambiente, ossia quelli che hanno la fitness maggiore In una popolazione compaiono continuamente nuovi caratteri, e nuove combinazioni di geni, sia per effetto dei meccanismi di mutazione spontanea, sia per la variabilità genetica assicurata dal crossing-over e dalla ricombinazione dei cromosomi nei gameti durante il processo di meiosi; questi fenomeni avvengono in modo casuale, e hanno quindi la stessa probabilità di verificarsi in qualsiasi popolazione. A seconda delle caratteristiche ambientali, però, la possibilità di sopravvivere e di riprodursi di alcuni individui, caratterizzati da un particolare patrimonio genetico, sarà maggiore rispetto a quella di altri, che risultano sfavoriti. Il cambiamento delle condizioni esterne determina inevitabilmente anche un cambiamento della fitness degli individui: quelli che nelle condizioni precedenti erano favoriti, possono in seguito risultare sfavoriti.
59 La selezione naturale Ad ogni generazione viene prodotto un numero sempre maggiore di individui in grado di sopravvivere fino alla riproduzione Il grado di variabilità fenotipica è elevato Una certa quota di questa variabilità fenotipica è ereditabile pertanto Gli individui con determinati caratteri (detti adattativi) sopravvivono e si riproducono in modo più efficiente rispetto agli altri. Questi caratteri sono ereditabili, per cui nella generazione successiva aumenterà in numero di individui che evidenzieranno tali caratteri
60

61 Human population diversity is low
62
63
64 Quel nostro antenato che morì di freddo Un teschio di uomo di Neanderthal Arcata dentaria Che fine fecero i nostri cugini neandertaliani vissuti a lungo in Europa prima di noi? Ora scrutando nel Dna estratto nei resti fossilizzati di 13 uomini vissuti tra l Europa e l Asia in un periodo tra 100 mila e 35 mila anni fa, ricercatori svedesi e spagnoli sono riusciti a precisare che cosa accadde ai lontani parenti. Esattamente 50 mila anni fa la maggior parte di loro si estinse dalla scena europea, quindi migliaia di anni prima che i nostri antenati Homo sapiens arrivassero dall Africa. Un piccolo gruppo, però, riuscì a sopravvivere per altri 10 mila anni rifugiandosi verso l ovest europeo. Poi la specie scomparve definitivamente. La causa prevalente pare legata al cambiamento climatico, come spiegano i ricercatori sulla rivista Molecular Biology and Evolution. Neanderthal lavorava le pelli e le usava per vestirsi unendole con fermagli d osso: una prova, questa, di una certa abilità tecnica.

65
66 La melanina contenuta nelle cellule dell epidermide protegge l individuo dagli effetti dannosi dei raggi UV che possono per esempio provocare gravi tumori della pelle come il melanoma. Ma la melanina impedisce la sintesi della vitamina D fondamentale per il corretto sviluppo osseo nell organismo, sintesi che necessita di un corretto apporto di raggi UVB. Popolazioni europee: plausibile che pelle chiara sia legata ad alimentazione a base di cereali e dunque povera di vitamina D3. Nel frumento presente precursore: ergosterolo che si converte in vitamina D sotto l azione dei raggi solari che attraversano la pelle e raggiungono il sangue tanto più facilmente quanto più essa è chiara Per questo motivo a nord le pressioni selettive avrebbero finito per favorire gli individui con la pelle più chiara. Questa teoria è stata per prima proposta quando alcuni studi all inizio del secolo scorso hanno dimostrato che le persone di pelle scura che vivono nell emisfero nord hanno il doppio di possibilità di sviluppare forme di rachitismo.
67 Extreme Adaptation The "Barreleye Fish" is an extreme example of nature's adaptive capability. This aquatic curiosity lives at depths (About 5,000 feet) where light cannot penetrate. This is how nature makes up for that deficit.
68 Apporto di vitamina D3 garantito dall alimentazione
69 Le dimensioni corporee delle popolazioni che vivono in regioni molto umide, come la foresta tropicale (pigmei, indios amazzonici) sono ridotte rispetto alla media umana. Infatti in un clima caldo e umido è conveniente essere piccoli per aumentare la superficie rispetto al volume, poiché è alla superficie che avviene l evaporazione del sudore, che consente al corpo di raffreddarsi. Inoltre un corpo piccolo necessita di un minore impiego di energia nel muoversi e quindi di una minore produzione di calore interno. Allo stesso modo, i capelli crespi, tipici dei pigmei della foresta dell Ituri (Congo) e dei neri in generale, trattengono maggiormente il sudore, prolungando l effetto di raffreddamento dovuto alla traspirazione.
70 Per contro, nei popoli che abitano regioni torride e desertiche (come i gruppi nilotici dell Africa orientale o i tuareg del Sahara) si nota una tendenza verso una figura allungata e sottile, che consente una dispersione del calore ottimale e un migliore raffreddamento.
71 Nelle popolazioni che vivono in climi freddi sono caratteristici i tratti somatici di tipo mongolico: il naso piccolo riduce il pericolo di congelamento e le narici affilate fanno in modo che l aria, arrivando più lentamente ai polmoni, si riscaldi e acquisti umidità; l accumulo di grasso sotto le palpebre protegge gli occhi dal freddo e lascia un apertura molto sottile, che riduce l esposizione dell occhio ai venti artici
72
73 Il linguaggio
74
75


76 The Red Queen complained that in Wonderland "it takes all the running you can do, to keep in the same place"

77 La selezione artificiale Selezione intenzionale e programmata delle caratteristiche biologiche di una specie o popolazione che non risponde ai criteri di autoorganizzazione propri dell ambiente naturale. La selezione artificiale si attua senza competizione per la sopravvivenza tra individui che mutino in modo casuale. In essa, infatti, i mutanti dotati di una qualsiasi proprietà desiderata possono essere favoriti dall intervento umano mirato alla produzione di particolari razze animali o vegetali mediante incroci selettivi, per scopi zootecnici o agronomici. Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
78

79 Risposta alla selezione in mais Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
80 La risposta alla selezione (R) Entità di variazione di un carattere sottoposto a selezione, nel corso di una generazione La risposta alla selezione si calcola sottraendo il fenotipo medio della popolazione originale da quello medio della prole. Es. mandria produce 80 l latte a settimana, la progenie ne produce 100. Risposta alla selezione= 20 l a settimana
81 La risposta alla selezione (R) Viene determinata prevalentemente da due fattori: ereditabilità in senso stretto grado di selezione degli individui da incrociare: differenza fenotipica di questi individui, che si misura con il differenziale di selezione (S). R = h 2 x S Il differenziale di selezione (S) è definito come la differenza fra il fenotipo medio degli individui parentali scelti, e quello medio della popolazione originale. Es. mandria produce 80 l latte a settimana, i parentali scelti per l incrocio ne producono 120. Differenziale di selezione = 40 l
82 Prevedere la risposta alla selezione (R) R = h 2 x S h 2 = V A V P Numero setole addominali in Drosophila Ereditabilità in senso stretto = 0,52 Numero medio setole in popolazione di partenza = 35,3 Numero medio setole in moscerini selezionati per l incrocio = 40,6 Pertanto, differenziale di selezione = 5,3 R = 0,52 x 5,3 = 2,8 R rappresenta l aumento atteso nella prole rispetto alla popolazione originale
83 L ereditabilità realizzata La risposta alla selezione può essere utilizzata per stimare l ereditabilità Quando G. Clayton osservò il numero medio di setole (37.9), scoprì che questo differiva dal numero stimato: 38.1 ( ) L ereditabilità in senso stretto puo pertanto essere calcolata rielaborando l equazione R = h 2 x S Ereditabilità realizzata h 2 = R S S: differenziale di selezione R: risposta alla selezione L ereditabilità realizzata è definita per mezzo di un esperimento di risposta alla selezione. Calcolare S: media parentali scelti media popolazione di partenza Calcolare R: media progenie media popolazione di partenza

84 Limiti della risposta alla selezione Quando un carattere è selezionato per molte generazioni non risponde più alla selezione, si stabilizza. Questo avviene perché si esaurisce la variabilità genetica della popolazione o perché la selezione naturale si oppone a uteriori cambiamenti: se cerchi di rimpicciolire i topi questi ad un certo punto diventano sterili per cui non posso trasmettere alla progenie i loro geni responsabili della ridotta dimensione Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
85 LA GENETICA DI POPOLAZIONE La genetica di popolazione riguarda la costituzione genetica di un gruppo di individui, e il modo in cui essa muta nel tempo

86 La genetica di popolazione Gli esseri viventi presentano un certo grado di variabilità genetica Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
87 Genetica di popolazione Nasce all inizio del 900 come logica conseguenza dello sviluppo della genetica mendeliana Studia geni e alleli non più nella singola famiglia, o nel singolo incrocio, ma in popolazioni di individui interfecondi Ricerca strumenti matematici per descrivere la variabilità genetica entro una popolazione. La struttura genetica di una popolazione considera i tipi e le frequenze dei genotipi e degli alleli di quella popolazione (frequenze geniche e genotipiche) Sviluppa modelli matematici che spieghino come tale variabilità si mantenga di generazione in generazione ovvero come e perché si verificano cambiamenti nelle frequenze geniche La genetica di popolazione riguarda cioè la costituzione genetica di un gruppo di individui e il modo in cui essa muta nel tempo
88 Popolazioni mendeliane e pool genico Si definisce popolazione mendeliana un gruppo di individui interfecondi che risiede all interno di confini geografici definiti. Se tutti i gameti prodotti da una popolazione mendeliana sono considerati come un ipotetico miscuglio di unità genetiche dalle quali nascerà la generazione successiva, ne emerge il concetto di pool genico. Il pool genico (o pool genetico) di una specie o di una popolazione è il set completo di alleli unici che verrebbero trovati esaminando il materiale genetico di ogni membro vivente di quella specie o popolazione, cioè l'insieme di tutti gli alleli di tutti i geni di tutti gli individui di una popolazione
89 Genetica di popolazione Parametri: Frequenza fenotipica: la proporzione di individui in una popolazione con un dato fenotipo Frequenza genotipica: la proporzione di uno specifico genotipo ad un dato locus, considerando che sono possibili molti genotipi diversi Frequenza allelica: la proporzione di uno specifico allele in un dato locus, considerando che la popolazione può avere da uno a più alleli a quel locus
90 Le frequenze genotipiche Frequenza genotipica: proporzione di uno specifico genotipo ad un dato locus, considerando che sono possibili molti genotipi diversi Si calcola sommando gli individui che possiedono lo specifico genotipo e poi dividendo per l ammontare complessivo di individui del campione frequenza (AA) = # individui AA N (# individui campione) frequenza (Aa) = # individui Aa N (# individui campione) frequenza (aa) = # individui aa N (# individui campione) La somma delle frequenze genotipiche è sempre uguale a 1
91 Le frequenze alleliche Il pool genetico di una popolazione può essere descritto anche in forma di frequenze alleliche: la proporzione di uno specifico allele in un dato locus. Il numero di alleli è sempre inferiore al numero genotipi! Se f(aa), f(aa) e f(aa) sono le frequenze di tre genotipi in un locus con due alleli, allora la frequenza p dell' allele A e la frequenza q dell'allele a si ottengono contando gli alleli. Poiché ogni omozigote AA consiste solo di alleli A, e dato che metà degli alleli di ogni eterozigote Aa sono alleli A, la frequenza totale p di alleli A nella popolazione si calcola come: frequenza di A In modo analogo, la frequenza q dell'allele a è data da frequenza di a Ci si aspetta che la somma di p e q sia 1, dato che essi sono le frequenze degli unici due alleli presenti. Infatti otteniamo: e da quest'ultima: q = 1 p e p = 1 q Se ci sono più di due diverse forme alleliche, la frequenza per ogni allele è semplicemente la frequenza del suo omozigote più la metà della somma delle frequenze per tutti gli altri eterozigoti in cui appare quel determinato allele.
92 Calcolo delle frequenze alleliche Somma delle copie di un particolare allele presente in un campione diviso l ammontare complessivo degli alleli del campione Si consideri una popolazione di dieci individui e un dato locus con due possibili alleli, A e a. Si supponga che i genotipi degli individui siano i seguenti: AA, Aa, AA, aa, Aa, AA, AA, Aa, Aa, e AA p = frequenza (A) = 2n AA + n Aa 2N (ogni individuo diploide possiede due alleli in un locus) q = frequenza (a) = 2n aa + n Aa 2N Si noti che p + q = 1, cioè la percentuale dei gameti A e a deve dare il 100%, se tiene conto di tutti i gameti nel pool genico.
93 Esempio di calcolo delle frequenze alleliche Si consideri una popolazione di dieci individui e un dato locus con due possibili alleli, A e a. Si supponga che i genotipi degli individui siano i seguenti: AA, Aa, AA, aa, Aa, AA, AA, Aa, Aa, e AA le frequenze alleliche dell'allele A e dell'allele a sono: ovvero c'è un 70% di probabilità che la popolazione riceva questo allele, e c'è un 30% di probabilità che la popolazione ottenga questo.
94 LEGGE DI HARDY-WEINBERG (1908) Il fine primario della genetica di popolazione consiste nel comprendere i processi che modellano il pool genico di un gruppo di individui. Bisogna pertanto comprendere gli effetti della riproduzione e delle leggi di Mendel sulle frequenze genotipiche e alleliche. Cioè: in che modo la segregazione degli alleli durante la formazione dei gameti e il loro combinarsi nel corso della fecondazione influenzano il pool di geni? La risposta a questa domanda risiede nella legge formulata indipendentemente da Hardy e da Weinberg nel 1908: modello matematico che valuta l effetto della riproduzione sulle frequenze genotipiche e alleliche di una popolazione
95 LEGGE DI HARDY-WEINBERG (1908) Modello matematico che valuta l effetto della riproduzione sulle frequenze genotipiche e alleliche di una popolazione In una popolazione, le frequenze alleliche e genotipiche si mantengono costanti di generazione in generazione Le frequenze genotipiche sono correlate alle frequenze alleliche da una semplice formula: A con frequenza p a con frequenza q Le frequenze dei tre possibili genotipi sono date da: p 2 + 2pq + q 2 = (p + q) 2 = 1 AA Aa aa p + q = 1
96 Equazione di HARDY-WEINBERG Dal momento che ciascun soggetto ha due alleli per un gene, la distribuzione dei genotipi alla generazione successiva può essere riassunta come segue: (p + q) 2 p 2 + 2pq + q 2 = 1 AA Aa aa Ogni soggetto della popolazione deriva infatti dall unione di due gameti, nei quali i due alleli A ed a possono presentarsi con probabilità p e q rispettivamente. Questa legge indica che la sola riproduzione non altera le frequenze alleliche o genotipiche, e che le frequenze alleliche determinano le frequenze genotipiche Quando i genotipi sono nelle proporzioni attese di p 2, 2pq e q 2, si dice che la popolazione è in equilibrio di Hardy-Weinberg
97 Quando una popolazione è in equilibrio di Hardy-Weinberg, le proporzioni dei genotipi sono stabilite dalle frequenze degli alleli Per un locus con 2 alleli, la frequenza dell eterozigote è più alta delle frequenze degli omozigoti quando le frequenze dei due alleli sono comprese fra 0,33 e 0,66. E massima (0,5) quando entrambe le frequenze sono = 0,5 Frequenza allele bassa: omozigoti rari Frequenza allele alta: omozigoti numerosi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
98 Condizioni necessarie per soddisfare la legge di Hardy e Weinberg 1. La popolazione è infinitamente estesa e si incrocia a caso (PANMISSIA) 2. Non c è selezione in atto a favore o contro un determinato fenotipo, tutti gli individui portatori dell uno o dell altro allele ad un dato locus sono ugualmente vitali e fertili 3. La popolazione è chiusa, cioè non vi sono movimenti migratori in atto 4. Non c è mutazione da uno stato allelico ad un altro, oppure il tasso di mutazione A -> a è uguale ad a -> A Questa legge si applica AD UN SINGOLO LOCUS. Quindi non è proprio necessario l accoppiamento casuale e la mancanza di selezione, migrazione e mutazione per tutte le caratteristiche, bensì solo per quel locus
99 Sommario condizioni necessarie per soddisfare la legge di Hardy e Weinberg 1. La popolazione è sufficientemente ampia 2. Gli individui della popolazione si accoppiano in maniera casuale 3. Le frequenze alleliche della popolazione non sono influenzate da selezione naturale, migrazioni, mutazioni
100 Implicazioni della legge di Hardy e Weinberg Se le condizioni elencate sopra fossero sempre rispettate, cioè se la popolazione fosse in equilibrio: L accoppiamento casuale genererebbe le frequenze di equilibrio p 2, 2pq, q 2 le frequenze genotipiche nelle popolazioni sarebbero sempre in equilibrio e IMMUTABILI nel tempo, cioè non ci sarebbe alcuna EVOLUZIONE Le frequenze genotipiche sarebbero stabilite dalle frequenze alleliche Questa legge afferma che la riproduzione, da sola, non potrà determinare l evoluzione
101 Condizioni necessarie per soddisfare la legge di Hardy e Weinberg Le condizioni elencate in precedenza sono teoriche, e non vengono mai rispettate in natura in modo assoluto Di fatto la legge di H.W. è applicabile a molti studi di popolazione ed è molto utile Bisogna comunque verificare con test statistici (es: test χ2) la sua applicabilità al campione in esame
102 Calcolo delle frequenze alleliche 1. E semplice per un locus con due alleli codominanti (p.es. locus gruppo sanguigno MN) Fenotipi: M MN N Genotipi: MM MN NN In un campione di 100 individui si contano 52 MM 36 MN 12 NN Frequenza dell allele M? n copie M presenti / n totale degli alleli ((52x2) + 36) / 200 = 0,7
103 Calcolo delle frequenze alleliche 2. Frequenza dell allele N? n copie N presenti/ n totale degli alleli ((12x2) + 36) / 200 = 0,3 Se la popolazione campionata rispetta equilibrio di H.W. le distribuzioni genotipiche attese sono: p 2 (MM)= 0,7x0,7= pq (MN)= 2x0,7x0,3=0,42 q 2 (NN)= 0,3x0,3=0,09 Il test del χ2 [Σ (oss.-att.) 2 /att.] mi dice che gli scostamenti fra osservati e attesi NON sono statisticamente significativi, quindi H.W. è rispettato: la popolazione è in equilibrio
104 Utilizzo pratico della legge di Hardy-Weinberg La fibrosi cistica è una malattia recessiva, rara, che colpisce in Italia 1/2000 neonati. Qual e la frequenza dei portatori sani? q 2 = 1/2000 (frequenza di aa) = q = = p = 1-q = ƒ Aa = 2pq = cioè 1/23 La probabilità che due Aa si incrocino è 1/23 x 1/23 = 1/529 Se entrambi i genitori sono Aa, ¼ dei figli sarà aa (malato). Quindi, il rischio che due italiani non imparentati e senza casi di malattia nelle rispettive famiglie abbiano un figlio malato è: 1/4 x 1/23 x 1/23 = 1/2116 cioè molto simile all 1/2000 osservato
105

106
107 L analisi dei profili del DNA si basa sulle assunzioni di Hardy-Weinberg Si studiano quelle parti del genoma che non influiscono sul fenotipo e che quindi non sono soggette a selezione naturale/artificiale, quali le sequenze corte ripetute che ricadono nelle regioni intergeniche
108 Ogni individuo può essere classificato come omozigote o eterozigote in base al numero di copie di una particolare ripetizione presenti nello stesso locus dei due cromosomi omologhi Individuo 1: eterozigote Individuo 2: omozigote Individuo 3: eterozigote
109
110 A ciascun numero di copie è assegnata una frequenza in base alla frequenza osservata in una particolare popolazione Se si considerano ripetizioni su cromosomi diversi, è possibile usare la regola del prodotto per calcolare la probabilità che una particolare combinazione di ripetizioni venga trovata in una popolazione. Si applica poi la legge di Hardy- Weinberg: la disposizione dei frammenti indica se un individuo è omozigote (p 2 o q 2 ) o eterozigote (2pq) per ogni ripetizione. Queste frequenze vengono poi moltiplicate.
111
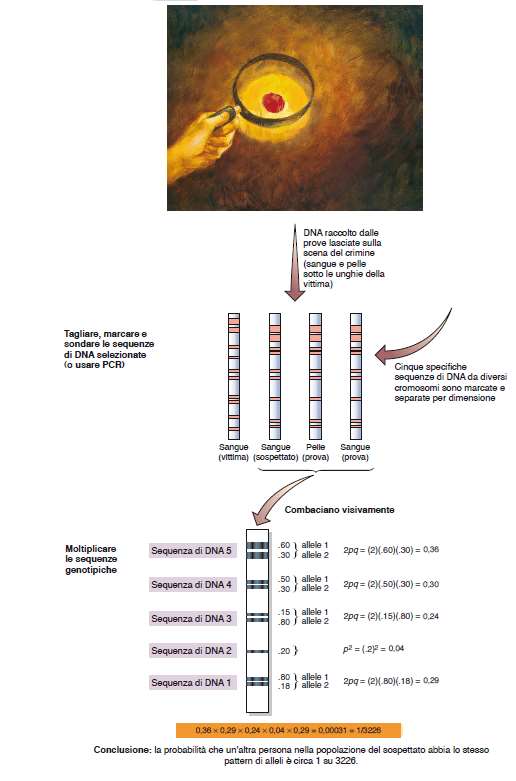

112
113 Dito medio più corto del secondo e del quarto: Carattere autosomico recessivo Se 9 individui su 100 hanno il dito medio più corto (dd), la frequenza è 9/100 o 0,09 Dato che dd corrisponde a q 2, q = 0,3 Dato che p + q = 1, p= 0,7 Quindi la proporzione dei tre genotipi generati quando i gameti si combinano casualmente, nella generazione 1 è: DD = p 2 = (0,7) 2 = 0,49 = 49% Dd = 2pq = 0,7 x 0,3 = 0,42 = 42% dd = q 2 = (0,3) 2 = 0,09 = 9% Quindi 9 individui su 100 avranno il dito medio più corto (dd), mantenendo la frequenza di 9/100 o 0,09 Pertanto la proporzione dei tre genotipi generati quando i gameti si combinano casualmente, nella generazione 2 sarà: DD = p 2 = (0,7) 2 = 0,49 = 49% Dd = 2pq = 0,7 x 0,3 = 0,42 = 42% dd = q 2 = (0,3) 2 = 0,09 = 9%
114 Quando i test statistici mostrano discrepanze significative fra le frequenze genotipiche osservate e le frequenze attese in base alla formula p2, 2pq, q2.. dobbiamo pensare che una o piu delle condizioni di validità della legge siano state violate Per esempio, nel caso del dito medio più corto del secondo e del quarto possiamo ipotizzare che sia errata l ipotesi che le persone scelgano il proprio compagno senza considerare la lunghezza delle dita (accoppiamento quindi non casuale)
115 Fattori che possono disturbare l equilibrio di Hardy-Weinberg Vari fattori possono disturbare l equilibrio di H. W.: 1. Accoppiamento non casuale 2. Mutazione 3. Migrazione 4. Inincrocio 5. Selezione 6. Dimensioni ridotte della popolazione Uno o piu fattori che violino le condizioni di validità della legge di H.W. possono promuovere cambiamenti nelle frequenze alleliche e genotipiche -> EVOLUZIONE
116 Fattori che cambiano frequenze geniche Mutazione Selezione Migrazione Introducono variabilità nel pool genico Dimensioni limitate di una popolazione (<1000 ii) Unioni non casuali (p.es. inincrocio) Riducono la variabilità del pool genico L'inincrocio è l'incrocio fra individui strettamente imparentali o consanguinei
117 Fattori che cambiano frequenze geniche. 1 Accoppiamento non casuale
118 Fattori che cambiano frequenze geniche. 2 Mutazione/Selezione La mutazione - evento raro e casuale - crea nuove varianti alleliche La selezione agisce sugli individui mutanti: se la mutazione è vantaggiosa verrà selezionata positivamente -> aumenta la frequenza della nuova variante allelica
119 Fattori che cambiano frequenze geniche. 3 Migrazione o flusso genico Influsso di geni provenienti da altre popolazioni Previene la differenza genetica fra popolazioni Aumenta la variabilità genetica entro popolazioni Effetti della migrazione sulle frequenze geniche (q): Δq = variazione frequenza genica i = frazione di immigrati qn = frequenza nei nativi qi = frequenza negli immigrati Δq= i(qn-qi)

120 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
121 Fattori che cambiano frequenze geniche. 4 L inincrocio (inbreeding) Accoppiamento fra individui che condividono una certa quota di alleli identici per discendenza Può essere la conseguenza di matrimoni fra membri di comunità ristrette e geograficamente isolate Può essere la conseguenza di matrimoni combinati fra consanguinei (p.es. primi cugini) per ragioni religiose, culturali, patrimoniali etc. Aumenta la probabilità di omozigosi nella prole per alleli recessivi rari Comparsa di malattie AR rare

122 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

123 Per la maggior parte delle specie a fecondazione incrociata, l inincrocio è dannoso in quanto aumenta la proporzione degli omozigoti favorendo quindi la comparsa di omozigoti recessivi deleteri o addirittura letali Depressione da inincrocio Riduzione della fitness conseguente alla pratica sistematica dell inbreeding Dovuta all omozigosi per alleli recessivi con effetto deleterio Si osservano ad es. riduzione di fertilità, ridotto tasso di sopravvivenza, comparsa di malformazioni, aumentata suscettibilità alle infezioni
 dell ibrido L'eterosi si riferisce all incrocio tra due linee pure che porta alla generazione di ibridi caratterizzati genotipicamente da un incremento di eterozigosità.")
124 Eteròsi (o vigore) dell ibrido L'eterosi si riferisce all incrocio tra due linee pure che porta alla generazione di ibridi caratterizzati genotipicamente da un incremento di eterozigosità. Il termine è l'opposto di inbreeding che si riferisce all'incrocio tra consanguinei aumentando l omozigosi cioè la presenza di alleli identici nello stesso locus. All'eterosi si associa il fenomeno osservato detto vigore dell'ibrido, in cui l'individuo prodotto dell'accoppiamento risulta con caratteristiche di fenotipo particolarmente vigorose
125 Vantaggio dell eterozigote si è verificato in africa occ. in ambiente malarico. gli eterozigoti β a β s hanno fitness maggiore degli omozigoti β a β a, perché risultano resistenti alla malaria (i loro globuli rossi sono inospitali al protozoo plasmodium) e anche degli omozigoti β s β s, gravemente colpiti dalla anemia falciforme. Quindi l allele dannoso viene mantenuto nella popolazione perché porta un vantaggio: polimorfismo bilanciato
126 Polimorfismo bilanciato In una popolazione, è il mantenimento di più di un allele in conseguenza della superiorità adattativa degli eterozigoti rispetto agli omozigoti. Rappresenta il risultato della pressione selettiva su una mutazione sfavorevole, che ne permette la persistenza in una popolazione. Per esempio, la persistenza del gene per l'anemia falciforme nelle popolazioni africane grazie alla protezione contro la malaria manifestata dagli eterozigoti. La diffusione della malaria rende la situazione eterozigote molto favorevole dal punto di vista evolutivo. In questo modo le frequenze dei fenotipi omozigoti ed eterozigoti si mantengono relativamente stabili.
127
128

129 Fattori che cambiano le frequenze geniche. 5 La selezione (naturale e artificiale)
130 Fattori che cambiano le frequenze geniche. 6 La deriva genetica La deriva genetica (Drift) è la variazione, dovuta al caso, delle frequenze alleliche in una piccola popolazione La deriva genetica è principalmente causata dall effetto del fondatore e dal collo di bottiglia
131 La deriva genetica fa variare le frequenze alleliche all interno delle popolazioni, portando a una riduzione di variabilità genetica attraverso la fissazione e la divergenza tra le popolazioni Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005

132 In questo esperimento sono prese in esame le frequenze di due alleli (bw75 e bw) che influiscono sul colore dell occhio in drosofila, in 107 popolazioni replicate, costituite da 8 maschi e 8 femmine, in cui la frequenza iniziale di bw75 era 0,5 Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005
133 Cause della deriva genetica EFFETTO DEL FONDATORE Si ha quando una nuova popolazione viene stabilita da un numero limitato di individui. Es. Situazioni di isolamento ecologico
134
135

136 Cause della deriva genetica EFFETTO COLLO DI BOTTIGLIA (Bottleneck) Si ha quando una popolazione si trova in condizioni sfavorevoli che riducono drasticamente il numero di individui Determina variazioni casuali nelle frequenze alleliche simili a quelle dovute all effetto del fondatore
GENETICA QUANTITATIVA
 GENETICA QUANTITATIVA Caratteri quantitativi e qualitativi I caratteri discontinui o qualitativi esibiscono un numero ridotto di fenotipi e mostrano una relazione genotipo-fenotipo semplice I caratteri
GENETICA QUANTITATIVA Caratteri quantitativi e qualitativi I caratteri discontinui o qualitativi esibiscono un numero ridotto di fenotipi e mostrano una relazione genotipo-fenotipo semplice I caratteri
Genetica quantitativa Evoluzione dei caratteri fenotipici
 Genetica quantitativa Evoluzione dei caratteri fenotipici I caratteri che sono determinati da molti geni mostrano una variazione continua Esempi nell uomo sono l altezza, l intelligenza, l abilità nello
Genetica quantitativa Evoluzione dei caratteri fenotipici I caratteri che sono determinati da molti geni mostrano una variazione continua Esempi nell uomo sono l altezza, l intelligenza, l abilità nello
Genetica dei caratteri quantitativi
 PAS Percorsi Abilitanti Speciali Classe di abilitazione A057 Scienza degli alimenti Tracciabilità genetica degli alimenti Genetica dei caratteri quantitativi 1 Concetti di base in genetica L informazione
PAS Percorsi Abilitanti Speciali Classe di abilitazione A057 Scienza degli alimenti Tracciabilità genetica degli alimenti Genetica dei caratteri quantitativi 1 Concetti di base in genetica L informazione
POPOLAZIONE. Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente
 POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l unita di base del cambiamento evolutivo, perchè permette
POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l unita di base del cambiamento evolutivo, perchè permette
Il legame genetico di una popolazione di n individui con la generazione precedente è rappresentato dai 2n gameti che hanno dato loro origine.
 La Genetica di popolazioni studia i fattori che influenzano la trasmissione dei caratteri ereditari a livello di popolazioni e i processi evolutivi che ne modificano la composizione genetica nello spazio
La Genetica di popolazioni studia i fattori che influenzano la trasmissione dei caratteri ereditari a livello di popolazioni e i processi evolutivi che ne modificano la composizione genetica nello spazio
La genetica dei caratteri quantitativi
 Studia l ereditarietà nelle popolazioni dei caratteri quantitativi Molti caratteri sono quantitativi, cioè hanno molte classi fenotipiche Il modo in cui si esprimono e vengono trasmessi i caratteri quantitativi
Studia l ereditarietà nelle popolazioni dei caratteri quantitativi Molti caratteri sono quantitativi, cioè hanno molte classi fenotipiche Il modo in cui si esprimono e vengono trasmessi i caratteri quantitativi
LA GENETICA QUANTITATIVA
 Genetica 5 LA GENETICA QUANTITATIVA Caratteri quantitativi vs. Caratteri qualitativi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 1910-1930 controversia Mendel/cararatteri continui Mendel
Genetica 5 LA GENETICA QUANTITATIVA Caratteri quantitativi vs. Caratteri qualitativi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 1910-1930 controversia Mendel/cararatteri continui Mendel
LA GENETICA QUANTITATIVA
 Genetica 5 LA GENETICA QUANTITATIVA Caratteri quantitativi vs. Caratteri qualitativi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 1910-1930 controversia Mendel/cararatteri continui Mendel
Genetica 5 LA GENETICA QUANTITATIVA Caratteri quantitativi vs. Caratteri qualitativi Pierce, GENETICA, Zanichelli editore S.p.A. Copyright 2005 1910-1930 controversia Mendel/cararatteri continui Mendel
GENETICA E PROBABILITA
 GENETICA E PROBABILITA Cromosoma:corpo cellulare contenete i geni disposti in ordine lineare, appare nel corso della mitosi (o della meiosi) come un filamento o un bastoncello di cromatina Gene:Unità di
GENETICA E PROBABILITA Cromosoma:corpo cellulare contenete i geni disposti in ordine lineare, appare nel corso della mitosi (o della meiosi) come un filamento o un bastoncello di cromatina Gene:Unità di
Mendel, il padre della genetica
 Mendel, il padre della genetica Gregor Mendel (1822-1894), monaco e naturalista, condusse esperimenti di genetica in un orto del monastero di Brno, nell odierna Repubblica Ceca. La genetica dell Ottocento
Mendel, il padre della genetica Gregor Mendel (1822-1894), monaco e naturalista, condusse esperimenti di genetica in un orto del monastero di Brno, nell odierna Repubblica Ceca. La genetica dell Ottocento
La nuova biologia.blu
 David Sadava, David M. Hillis, H. Craig Heller, May R. Berenbaum La nuova biologia.blu Genetica, DNA ed evoluzione PLUS 2 Capitolo B1 Da Mendel ai modelli di ereditarietà 3 Mendel, il padre della genetica
David Sadava, David M. Hillis, H. Craig Heller, May R. Berenbaum La nuova biologia.blu Genetica, DNA ed evoluzione PLUS 2 Capitolo B1 Da Mendel ai modelli di ereditarietà 3 Mendel, il padre della genetica
Lezione 3. I geni nelle popolazioni. Lezione modificata da Rosenblum
 Lezione 3 I geni nelle popolazioni Lezione modificata da Rosenblum http://people.ibest.uidaho.edu/~bree/ Graur and Li: Capitolo 2 L evoluzione è Il processo che risulta dal cambiamento delle caratteristiche
Lezione 3 I geni nelle popolazioni Lezione modificata da Rosenblum http://people.ibest.uidaho.edu/~bree/ Graur and Li: Capitolo 2 L evoluzione è Il processo che risulta dal cambiamento delle caratteristiche
POPOLAZIONE. Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente
 POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l'unità di base del cambiamento evolutivo, perché permette
POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l'unità di base del cambiamento evolutivo, perché permette
Legge di Hardy-Weinberg
 Legge di Hardy-Weinberg Popolazione in equilibrio Popolazione infinitamente grande No mutazioni No migrazione (geni non sono introdotti o persi) No selezione Incroci casuali Unione fra genotipi è casuale
Legge di Hardy-Weinberg Popolazione in equilibrio Popolazione infinitamente grande No mutazioni No migrazione (geni non sono introdotti o persi) No selezione Incroci casuali Unione fra genotipi è casuale
Struttura genetica delle popolazioni. Piante a propagazione vegetativa Piante autogame Piante allogame
 Struttura genetica delle popolazioni Piante a propagazione vegetativa Piante autogame Piante allogame PIANTE A PROPAGAZIONE VEGETATIVA La popolazione è costituita da un numero infinitamente grande di individui
Struttura genetica delle popolazioni Piante a propagazione vegetativa Piante autogame Piante allogame PIANTE A PROPAGAZIONE VEGETATIVA La popolazione è costituita da un numero infinitamente grande di individui
I paleontologi utilizzano numerosi metodi per la datazione dei fossili. Datazione relativa Scala temporale geologica
 I paleontologi utilizzano numerosi metodi per la datazione dei fossili Datazione relativa Scala temporale geologica Gli strati di rocce sedimentarie permettono di attribuire un età relativa ai fossili
I paleontologi utilizzano numerosi metodi per la datazione dei fossili Datazione relativa Scala temporale geologica Gli strati di rocce sedimentarie permettono di attribuire un età relativa ai fossili
Legge di Hardy-Weinberg
 Legge di Hardy-Weinberg Popolazione in equilibrio Popolazione infinitamente grande No mutazioni No migrazione (geni non sono introdotti o persi) No selezione Incroci casuali Unione fra genotipi è casuale
Legge di Hardy-Weinberg Popolazione in equilibrio Popolazione infinitamente grande No mutazioni No migrazione (geni non sono introdotti o persi) No selezione Incroci casuali Unione fra genotipi è casuale
Genetica della trasmissione dei caratteri. Genetica molecolare. Genetica di popolazione
 Genetica della trasmissione dei caratteri Genetica molecolare Genetica di popolazione Individuò regole attraverso cui i caratteri dei genitori si ripresentano nella progenie caratteristiche della prole
Genetica della trasmissione dei caratteri Genetica molecolare Genetica di popolazione Individuò regole attraverso cui i caratteri dei genitori si ripresentano nella progenie caratteristiche della prole
Vale a dire le proporzioni di ciascun allele sul totale degli alleli di quel locus nella popolazione
 Vale a dire le proporzioni di ciascun allele sul totale degli alleli di quel locus nella popolazione Gli organismi geneticamente sono strutturati nel seguente livello gerarchico Geni Genotipi (individui)
Vale a dire le proporzioni di ciascun allele sul totale degli alleli di quel locus nella popolazione Gli organismi geneticamente sono strutturati nel seguente livello gerarchico Geni Genotipi (individui)
Equilibrio di Hardy-Weinberg
 Equilibrio di Hardy-Weinberg L'equilibrio di Hardy-Weinberg, o legge di Hardy-Weinberg [1][2] è un modello della genetica delle popolazioni che postula che all'interno di una popolazione (ideale), vi è
Equilibrio di Hardy-Weinberg L'equilibrio di Hardy-Weinberg, o legge di Hardy-Weinberg [1][2] è un modello della genetica delle popolazioni che postula che all'interno di una popolazione (ideale), vi è
ciascun gamete riceve con la stessa probabilità l uno o l altro di essi
 LE LEGGI DI MENDEL I primi tre postulati di Mendel: 1.I caratteri genetici sono controllati da fattori che esistono in coppie nei singoli organismi 2.Quando due fattori diversi, responsabili di un unico
LE LEGGI DI MENDEL I primi tre postulati di Mendel: 1.I caratteri genetici sono controllati da fattori che esistono in coppie nei singoli organismi 2.Quando due fattori diversi, responsabili di un unico
ciascun gamete riceve con la stessa probabilità l uno o l altro di essi
 LE LEGGI DI MENDEL I primi tre postulati di Mendel: 1.I caratteri genetici sono controllati da fattori che esistono in coppie nei singoli organismi 2.Quando due fattori diversi, responsabili di un unico
LE LEGGI DI MENDEL I primi tre postulati di Mendel: 1.I caratteri genetici sono controllati da fattori che esistono in coppie nei singoli organismi 2.Quando due fattori diversi, responsabili di un unico
8. La genetica studia la trasmissione dei caratteri ereditari
 8. La genetica studia la trasmissione dei caratteri ereditari La genetica studia le modalità di trasmissione dei caratteri ereditari e il modo in cui questi si manifestano. Ogni genitore trasmette ai figli
8. La genetica studia la trasmissione dei caratteri ereditari La genetica studia le modalità di trasmissione dei caratteri ereditari e il modo in cui questi si manifestano. Ogni genitore trasmette ai figli
Ampliamenti delle leggi di Mendel
 Ampliamenti delle leggi di Mendel 1 La dominanza incompleta Nella dominanza incompleta gli eterozigoti mostrano un fenotipo intermedio a quello dei genitori, in accordo con le leggi di Mendel. Entrambi
Ampliamenti delle leggi di Mendel 1 La dominanza incompleta Nella dominanza incompleta gli eterozigoti mostrano un fenotipo intermedio a quello dei genitori, in accordo con le leggi di Mendel. Entrambi
Mendel e l ereditarietà. Anna Onofri
 Mendel e l ereditarietà Anna Onofri Mendel, il padre della genetica Gregor Mendel (1822-1894) era un monaco e naturalista che condusse i suoi studi nel giardino del monastero di Brno, nell odierna Repubblica
Mendel e l ereditarietà Anna Onofri Mendel, il padre della genetica Gregor Mendel (1822-1894) era un monaco e naturalista che condusse i suoi studi nel giardino del monastero di Brno, nell odierna Repubblica
David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis. Biologia.blu B - Le basi molecolari della vita e dell evoluzione
 David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis Biologia.blu B - Le basi molecolari della vita e dell evoluzione 1 L evoluzione e i suoi meccanismi 2 Il viaggio di Charles
David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis Biologia.blu B - Le basi molecolari della vita e dell evoluzione 1 L evoluzione e i suoi meccanismi 2 Il viaggio di Charles
La nuova biologia.blu
 David Sadava, David M. Hillis, H. Craig Heller, May R. Berenbaum La nuova biologia.blu Genetica, DNA ed evoluzione PLUS 2 Capitolo B5 L evoluzione e l origine delle specie viventi 3 La teoria di Darwin
David Sadava, David M. Hillis, H. Craig Heller, May R. Berenbaum La nuova biologia.blu Genetica, DNA ed evoluzione PLUS 2 Capitolo B5 L evoluzione e l origine delle specie viventi 3 La teoria di Darwin
GENETICA DI POPOLAZIONI. Si occupa dello studio della costituzione genetica delle popolazioni e di come cambia da generazione a generazione
 GENETICA DI POPOLAZIONI Si occupa dello studio della costituzione genetica delle popolazioni e di come cambia da generazione a generazione Legge di Hardy Weinberg (Equilibrium) Godfrey Harold Hardy Mt
GENETICA DI POPOLAZIONI Si occupa dello studio della costituzione genetica delle popolazioni e di come cambia da generazione a generazione Legge di Hardy Weinberg (Equilibrium) Godfrey Harold Hardy Mt
Riassunto della teoria di Darwin
 Riassunto della teoria di Darwin Le specie si evolvono Antenato comune Le specie si moltiplicano L evoluzione è un processo graduale L evoluzione è regolata dalla selezione naturale MICROEVOLUZIONE DISCENDENZA
Riassunto della teoria di Darwin Le specie si evolvono Antenato comune Le specie si moltiplicano L evoluzione è un processo graduale L evoluzione è regolata dalla selezione naturale MICROEVOLUZIONE DISCENDENZA
Caratteristiche di un carattere complesso (o multifattoriale) Il fenotipo è influenzato dall azione di più geni e da un elevata componente ambientale.
 Il fenotipo è influenzato dall azione di più geni e da un elevata componente ambientale.") Caratteristiche di un carattere complesso (o multifattoriale) Il fenotipo è influenzato dall azione di più geni e da un elevata componente ambientale. La Psoriasi è una malattia genetica? Se tu sei affetto
Caratteristiche di un carattere complesso (o multifattoriale) Il fenotipo è influenzato dall azione di più geni e da un elevata componente ambientale. La Psoriasi è una malattia genetica? Se tu sei affetto
Da Mendel ai modelli di ereditarietà
 Capitolo B1 Da Mendel ai modelli di ereditarietà 1 Mendel, il padre della genetica Gregor Mendel (1822-1894), monaco e naturalista, condusse esperimenti di genetica in un orto del monastero di Brno, nell
Capitolo B1 Da Mendel ai modelli di ereditarietà 1 Mendel, il padre della genetica Gregor Mendel (1822-1894), monaco e naturalista, condusse esperimenti di genetica in un orto del monastero di Brno, nell
Concetti di miglioramento genetico
 Concetti di miglioramento genetico Categories : Anno 2008, N. 68-1 novembre 2008 Alcuni concetti base per affrontare lo studio del miglioramento genetico delle specie animali in produzione zootecnica di
Concetti di miglioramento genetico Categories : Anno 2008, N. 68-1 novembre 2008 Alcuni concetti base per affrontare lo studio del miglioramento genetico delle specie animali in produzione zootecnica di
La genetica è la scienza dell ereditarietà: Studia la trasmissione delle caratteristiche ereditarie, che distinguono gli individui tra di loro
 La genetica è la scienza dell ereditarietà: Studia la trasmissione delle caratteristiche ereditarie, che distinguono gli individui tra di loro la comprensione di come i geni si trasmettono da genitori
La genetica è la scienza dell ereditarietà: Studia la trasmissione delle caratteristiche ereditarie, che distinguono gli individui tra di loro la comprensione di come i geni si trasmettono da genitori
Genetica dei caratteri complessi
 Genetica dei caratteri complessi Lezione 3 1 Definizione Eredità dei caratteri complessi Determinati caratteri vengono trovati con maggior frequenza in alcune famiglie, non mostrando però la normale segregazione
Genetica dei caratteri complessi Lezione 3 1 Definizione Eredità dei caratteri complessi Determinati caratteri vengono trovati con maggior frequenza in alcune famiglie, non mostrando però la normale segregazione
- Riproduzione riservata - 1
 Genetica mendeliana; Mendel elaborò tre leggi sull ereditarietà dei caratteri: - Legge della segregazione - Legge della dominanza - Legge dell assortimento indipendente Legge della segregazione; Per poter
Genetica mendeliana; Mendel elaborò tre leggi sull ereditarietà dei caratteri: - Legge della segregazione - Legge della dominanza - Legge dell assortimento indipendente Legge della segregazione; Per poter
Riassunto della teoria di Darwin
 Riassunto della teoria di Darwin Le specie si evolvono Antenato comune Le specie si moltiplicano L evoluzione è un processo graduale L evoluzione è regolata dalla selezione naturale MICROEVOLUZIONE DISCENDENZA
Riassunto della teoria di Darwin Le specie si evolvono Antenato comune Le specie si moltiplicano L evoluzione è un processo graduale L evoluzione è regolata dalla selezione naturale MICROEVOLUZIONE DISCENDENZA
SECONDA LEGGE DI MENDEL
 SECONDA LEGGE DI MENDEL Seconda Legge di Mendel: principio dell assortimento indipendente Il rapporto fenotipico 9:3:3:1 in incroci di diibridi Seconda Legge di Mendel: principio dell assortimento indipendente
SECONDA LEGGE DI MENDEL Seconda Legge di Mendel: principio dell assortimento indipendente Il rapporto fenotipico 9:3:3:1 in incroci di diibridi Seconda Legge di Mendel: principio dell assortimento indipendente
Jay Phelan, Maria Cristina Pignocchino. Scopriamo la biologia
 Jay Phelan, Maria Cristina Pignocchino Scopriamo la biologia Capitolo 5 La basi della genetica 3 1. La genetica e i caratteri ereditari /1 La genetica è la scienza che studia le leggi e i meccanismi di
Jay Phelan, Maria Cristina Pignocchino Scopriamo la biologia Capitolo 5 La basi della genetica 3 1. La genetica e i caratteri ereditari /1 La genetica è la scienza che studia le leggi e i meccanismi di
Chapter 21 Population Genetics
 Chapter 21 Population Genetics CORSO GENETICA LAUREA TRIENNALE ROMA TRE Prof. A. Antoccia Copyright 2010 Pearson Education Inc. Aspetti diversi dell ereditarieta La genetica classica studia i processi
Chapter 21 Population Genetics CORSO GENETICA LAUREA TRIENNALE ROMA TRE Prof. A. Antoccia Copyright 2010 Pearson Education Inc. Aspetti diversi dell ereditarieta La genetica classica studia i processi
GENETICA DELLE POPOLAZIONI
 GENETICA DELLE POPOLAZIONI La teoria dell evoluzione evoluzione di Darwin aveva un grande difetto: non spiegava l origine l della variazione e come particolari varianti venissero ereditate. Nel 1900, la
GENETICA DELLE POPOLAZIONI La teoria dell evoluzione evoluzione di Darwin aveva un grande difetto: non spiegava l origine l della variazione e come particolari varianti venissero ereditate. Nel 1900, la
LE LEGGI DI MENDEL SPIEGANO LA TRASMISSIONE DEI CARATTERI MONOFATTORIALI!!
 CARATTERE = una qualsiasi caratteristica di un organismo FENOTIPO (INSIEME DEI CARATTERI OSSERVABILI DELL ORGANISMO) GENOTIPO (COSTITUZIONE GENETICA DELL'INDIVIDUO, PATRIMONIO EREDITARIO) EREDITARIO: carattere
CARATTERE = una qualsiasi caratteristica di un organismo FENOTIPO (INSIEME DEI CARATTERI OSSERVABILI DELL ORGANISMO) GENOTIPO (COSTITUZIONE GENETICA DELL'INDIVIDUO, PATRIMONIO EREDITARIO) EREDITARIO: carattere
GENOTIPO: costituzione genetica di un individuo (sia riferito ad un singolo gene, sia all insieme dei suoi geni).
.") DNA e geni Cosa sono i geni? Sono tratti di DNA ben delimitati Sono sequenze codificanti: tramite le istruzioni contenute in uno specifico gene viene prodotta una caratteristica fenotipica (carattere).
DNA e geni Cosa sono i geni? Sono tratti di DNA ben delimitati Sono sequenze codificanti: tramite le istruzioni contenute in uno specifico gene viene prodotta una caratteristica fenotipica (carattere).
!In una popolazione infinitamente grande, in cui gli
 Cosa vuol dire equilibrio!in una popolazione infinitamente grande, in cui gli incroci avvengono casualmente, in cui non vi siano mutazioni, ne migrazioni, ne selezione le frequenze alleliche in locus con
Cosa vuol dire equilibrio!in una popolazione infinitamente grande, in cui gli incroci avvengono casualmente, in cui non vi siano mutazioni, ne migrazioni, ne selezione le frequenze alleliche in locus con
Le proprietà del materiale gene0co possono essere studiate a tre livelli: Individuale:presenza o assenza di un certo cara3ere;
 Aspetti diversi dello studio dell ereditarieta Le proprietà del materiale gene0co possono essere studiate a tre livelli: Individuale:presenza o assenza di un certo cara3ere; Famiglia:dominanza o recessività;
Aspetti diversi dello studio dell ereditarieta Le proprietà del materiale gene0co possono essere studiate a tre livelli: Individuale:presenza o assenza di un certo cara3ere; Famiglia:dominanza o recessività;
Come interagiscono i geni?
 Come interagiscono i geni? I primi genetisti, Mendel compreso, lavorarono dando per scontato che ogni gene influenzasse un solo carattere, indipendentemente dall azione degli altri geni. Col procedere
Come interagiscono i geni? I primi genetisti, Mendel compreso, lavorarono dando per scontato che ogni gene influenzasse un solo carattere, indipendentemente dall azione degli altri geni. Col procedere
On the Origin of Species by Means of Natural Selection, 1859
 On the Origin of Species by Means of Natural Selection, 1859 Darwin Reperti fossili Biogeografia Distribuzione geografica delle specie Es: marsupiali in Australia Anatomia comparata Omologia Analogia Embriologia
On the Origin of Species by Means of Natural Selection, 1859 Darwin Reperti fossili Biogeografia Distribuzione geografica delle specie Es: marsupiali in Australia Anatomia comparata Omologia Analogia Embriologia
Gli eventi caratteristici della meiosi sono:
 Gli eventi caratteristici della meiosi sono: 1. la segregazione (=separazione) dei cromosomi omologhi; 2. l assortimento indipendente dei cromosomi; 3. il crossing-over LA COMPRENSIONE DI COME I GENI SI
Gli eventi caratteristici della meiosi sono: 1. la segregazione (=separazione) dei cromosomi omologhi; 2. l assortimento indipendente dei cromosomi; 3. il crossing-over LA COMPRENSIONE DI COME I GENI SI
Mendel e i princìpi dell ereditarietà
 Mendel e i princìpi dell ereditarietà Gregor Mendel Gregor Mendel formulò le leggi di base dell ereditarietà Prima di Mendel si credeva all ereditarietà per mescolanza dei caratteri. I risultati degli
Mendel e i princìpi dell ereditarietà Gregor Mendel Gregor Mendel formulò le leggi di base dell ereditarietà Prima di Mendel si credeva all ereditarietà per mescolanza dei caratteri. I risultati degli
LA TEORIA DI DARWIN SULLA SELEZIONE NATURALE SOSTIENE: L. Scientifico "G. Rummo"-BN - Prof.ssa R. Spagnuolo
 LA TEORIA DI DARWIN SULLA SELEZIONE NATURALE SOSTIENE: Il NEO-DARWINISMO SUCCESSIVAMENTE: Incorpora le nuove conoscenze dovute alla genetica Vede l evoluzione attraverso la genetica di popolazioni (frequenza
LA TEORIA DI DARWIN SULLA SELEZIONE NATURALE SOSTIENE: Il NEO-DARWINISMO SUCCESSIVAMENTE: Incorpora le nuove conoscenze dovute alla genetica Vede l evoluzione attraverso la genetica di popolazioni (frequenza
Cromosomi sessuali. Le cellule maschili e femminili differiscono per i cromosomi sessuali o
 Cromosomi sessuali Le cellule maschili e femminili differiscono per i cromosomi sessuali o cromosomi del sesso o eterosomi (cosiddetti perché hanno forma diversa). Nell uomo e in molte altre specie (ma
Cromosomi sessuali Le cellule maschili e femminili differiscono per i cromosomi sessuali o cromosomi del sesso o eterosomi (cosiddetti perché hanno forma diversa). Nell uomo e in molte altre specie (ma
CARATTERI QUANTITATIVI misurabili su una scala metrica variabilità continua
 CARATTERI DI INTERESSE ZOOTECNICO CARATTERI QUANTITATIVI misurabili su una scala metrica variabilità continua CARATTERI QUALITATI VI classificazione delle unità sperimentali variabilità discontinua CARATTERI
CARATTERI DI INTERESSE ZOOTECNICO CARATTERI QUANTITATIVI misurabili su una scala metrica variabilità continua CARATTERI QUALITATI VI classificazione delle unità sperimentali variabilità discontinua CARATTERI
Frequenze alleliche. Esempio: Locus biallelico: A, a. Freq allele A (p) = N A /N tot. p + q = 1. Freq allele a (q) = N a /N tot
 = N A /N tot. p + q = 1. Freq allele a (q) = N a /N tot") Frequenze alleliche Esempio: Locus biallelico: A, a Freq allele A (p) = N A /N tot Freq allele a (q) = N a /N tot p + q = 1 N tot = numero totale di alleli nella popolazione= doppio numero totale individui
Frequenze alleliche Esempio: Locus biallelico: A, a Freq allele A (p) = N A /N tot Freq allele a (q) = N a /N tot p + q = 1 N tot = numero totale di alleli nella popolazione= doppio numero totale individui
Hardy-Weinberg LEZIONE 4. By NA 1
 Hardy-Weinberg LEZIONE 4 By NA 1 La Genetica di popolazione La genetica di popolazione studia l ereditarieta di caratteri determinati da uno o pochi geni in gruppi di individui By NA 2 La genetica di popolazione
Hardy-Weinberg LEZIONE 4 By NA 1 La Genetica di popolazione La genetica di popolazione studia l ereditarieta di caratteri determinati da uno o pochi geni in gruppi di individui By NA 2 La genetica di popolazione
Collega ciascun termine con la sua definizione A. Fenotipo B. B. Genotipo C. C. Carattere D. D. Omozigote E. E. Eterozigote 1.
 Collega ciascun termine con la sua definizione A. Fenotipo B. B. Genotipo C. C. Carattere D. D. Omozigote E. E. Eterozigote 1. Insieme delle caratteristiche contenute nei geni, sia quelle manifeste, sia
Collega ciascun termine con la sua definizione A. Fenotipo B. B. Genotipo C. C. Carattere D. D. Omozigote E. E. Eterozigote 1. Insieme delle caratteristiche contenute nei geni, sia quelle manifeste, sia
La teoria cromosomica dell'ereditarietà mette in relazione il comportamento dei cromosomi con l'eredità dei caratteri secondo Mendel
 9 3 3 1 La teoria cromosomica dell'ereditarietà mette in relazione il comportamento dei cromosomi con l'eredità dei caratteri secondo Mendel Il meccanismo con cui si ereditano i caratteri è spiegabile
9 3 3 1 La teoria cromosomica dell'ereditarietà mette in relazione il comportamento dei cromosomi con l'eredità dei caratteri secondo Mendel Il meccanismo con cui si ereditano i caratteri è spiegabile
CORSO DI GENETICA. Roberto Piergentili. Università di Urbino Carlo Bo LE LEGGI DI MENDEL
 CORSO DI GENETICA LE LEGGI DI MENDEL Genotipo e Fenotipo (Johannsen 1909) Fenotipo: indica il complesso delle caratteristiche manifeste di un individuo, sia morfologiche che funzionali, prodotte dall interazione
CORSO DI GENETICA LE LEGGI DI MENDEL Genotipo e Fenotipo (Johannsen 1909) Fenotipo: indica il complesso delle caratteristiche manifeste di un individuo, sia morfologiche che funzionali, prodotte dall interazione
EPI-GENETICA: EREDITARIETA E AMBIENTE
 EPI-GENETICA: EREDITARIETA E AMBIENTE EREDITARIETA L'ereditarietà genetica è la trasmissione, da una generazione alle successive, dei caratteri originati dall'assetto genetico. Il patrimonio ereditario
EPI-GENETICA: EREDITARIETA E AMBIENTE EREDITARIETA L'ereditarietà genetica è la trasmissione, da una generazione alle successive, dei caratteri originati dall'assetto genetico. Il patrimonio ereditario
GENETICA. Modulo di 6 CFU. Esame integrato di BIOCHIMICA&GENETICA Secondo anno del corso di laurea triennale in SCIENZE AMBIENTALI
 GENETICA Modulo di 6 CFU Esame integrato di BIOCHIMICA&GENETICA Secondo anno del corso di laurea triennale in SCIENZE AMBIENTALI Docente: Flavia Cerrato Dip.to Scienze e Tecnologie Ambientali, Biologiche
GENETICA Modulo di 6 CFU Esame integrato di BIOCHIMICA&GENETICA Secondo anno del corso di laurea triennale in SCIENZE AMBIENTALI Docente: Flavia Cerrato Dip.to Scienze e Tecnologie Ambientali, Biologiche
L epistasi è una forma d interazione tra geni, in base alla quale un gene maschera l espressione fenotipica di un altro gene
 L epistasi è una forma d interazione tra geni, in base alla quale un gene maschera l espressione fenotipica di un altro gene L interazione genica è causa di rapporti diibridi atipici. Consideriamo un
L epistasi è una forma d interazione tra geni, in base alla quale un gene maschera l espressione fenotipica di un altro gene L interazione genica è causa di rapporti diibridi atipici. Consideriamo un
Ricombinazione di geni associati sullo stesso cromosoma e mappe genetiche
 Ricombinazione di geni associati sullo stesso cromosoma e mappe genetiche doppio eterozigote Segregazione indipendente di due caratteri: geni non associati omozigote recessivo M= foglie normali m= foglie
Ricombinazione di geni associati sullo stesso cromosoma e mappe genetiche doppio eterozigote Segregazione indipendente di due caratteri: geni non associati omozigote recessivo M= foglie normali m= foglie
Gli studi che Mendel effettuò vennero alla luce soltanto diversi anni dopo la sua morte e costituirono la base di una nuova scienza: la.
 Gli studi che Mendel effettuò vennero alla luce soltanto diversi anni dopo la sua morte e costituirono la base di una nuova scienza: la Genetica Le leggi dell ereditarietà Geni e alleli Ogni carattere
Gli studi che Mendel effettuò vennero alla luce soltanto diversi anni dopo la sua morte e costituirono la base di una nuova scienza: la Genetica Le leggi dell ereditarietà Geni e alleli Ogni carattere
CAPITOLO 11 STRUTTURA GENETICA DELLE POPOLAZIONI LIGUORI EDITORE
 CAPITOLO 11 STRUTTURA GENETICA DELLE POPOLAZIONI LIGUORI EDITORE 1 Genetics Genetica delle popolazioni 2 Cosa è una popolazione? Entità continua nello spazio e nel tempo La successione è garantita dalla
CAPITOLO 11 STRUTTURA GENETICA DELLE POPOLAZIONI LIGUORI EDITORE 1 Genetics Genetica delle popolazioni 2 Cosa è una popolazione? Entità continua nello spazio e nel tempo La successione è garantita dalla
LA GENETICA E L EREDITARIETA
 LA GENETICA E L EREDITARIETA Come si trasmettono i caratteri ereditari? Il primo studioso che osservò le caratteristiche umane e la modalità di trasmissione fu Ippocrate (460-377 a. C.) 1 Come si trasmettono
LA GENETICA E L EREDITARIETA Come si trasmettono i caratteri ereditari? Il primo studioso che osservò le caratteristiche umane e la modalità di trasmissione fu Ippocrate (460-377 a. C.) 1 Come si trasmettono
Corso di Genetica -Lezione 2- Cenci
 Corso di Genetica -Lezione 2- Cenci Gli incroci diibridi DIIBRIDO, una pianta eterozigote per due geni allo stesso tempo. I differenti tipi di cellule germinali [uova o polline] di un ibrido sono prodotti,
Corso di Genetica -Lezione 2- Cenci Gli incroci diibridi DIIBRIDO, una pianta eterozigote per due geni allo stesso tempo. I differenti tipi di cellule germinali [uova o polline] di un ibrido sono prodotti,
MENDEL E L EREDITARIETA
 MENDEL E L EREDITARIETA Gregor Mendel (1822-1884) era un monaco ceco, biologo e matematico; E considerato, per le sue osservazioni sulla trasmissione dei caratteri ereditari, il precursore della moderna
MENDEL E L EREDITARIETA Gregor Mendel (1822-1884) era un monaco ceco, biologo e matematico; E considerato, per le sue osservazioni sulla trasmissione dei caratteri ereditari, il precursore della moderna
I MOTORI DELL EVOLUZIONE PT5. POMERIGGIO DI AGGIORNAMENTO PROF. M.A. ZORDAN, Ph.D UNIVERSITÀ DEGLI STUDI DI PADOVA
 I MOTORI DELL EVOLUZIONE PT5 POMERIGGIO DI AGGIORNAMENTO 23.03.2011 PROF. M.A. ZORDAN, Ph.D UNIVERSITÀ DEGLI STUDI DI PADOVA 1 LA VARIABILITÀ GENETICA E LA SELEZIONE NATURALE 2 Popolazioni di organismi
I MOTORI DELL EVOLUZIONE PT5 POMERIGGIO DI AGGIORNAMENTO 23.03.2011 PROF. M.A. ZORDAN, Ph.D UNIVERSITÀ DEGLI STUDI DI PADOVA 1 LA VARIABILITÀ GENETICA E LA SELEZIONE NATURALE 2 Popolazioni di organismi
Cenni di. genetica di popolazioni
 Cenni di genetica di popolazioni In termini genetici una popolazione è l insieme dei geni di un dato numero di individui. La genetica di popolazione riguarda pertanto non direttamente i fenotipi degli
Cenni di genetica di popolazioni In termini genetici una popolazione è l insieme dei geni di un dato numero di individui. La genetica di popolazione riguarda pertanto non direttamente i fenotipi degli
L EVOLUZIONE COME CARATTERISTICA DELLE POPOLAZIONI
 L EVOLUZIONE COME CARATTERISTICA DELLE POPOLAZIONI Per capire come il concetto di evoluzione si leghi a quello di ereditarietà occorre prendere in considerazione l individuo non da solo ma in rapporto
L EVOLUZIONE COME CARATTERISTICA DELLE POPOLAZIONI Per capire come il concetto di evoluzione si leghi a quello di ereditarietà occorre prendere in considerazione l individuo non da solo ma in rapporto
Variabilità genetica
 Variabilità genetica La variabiltà della struttura genetica esistente nelle popolazioni è superiore a quella riconducibile alle differenze fenotipiche. Essa può essere misurata analizzando i polimorfismi
Variabilità genetica La variabiltà della struttura genetica esistente nelle popolazioni è superiore a quella riconducibile alle differenze fenotipiche. Essa può essere misurata analizzando i polimorfismi
GRUPPI SANGUIGNI. Supponendo che la popolazione italiana sia H-W, calcola la probabilità di ogni singolo allele e di ogni genotipo
 GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
Statistica Elementare
 Statistica Elementare 1. Frequenza assoluta Per popolazione si intende l insieme degli elementi che sono oggetto di una indagine statistica, ovvero l insieme delle unità, dette unità statistiche o individui
Statistica Elementare 1. Frequenza assoluta Per popolazione si intende l insieme degli elementi che sono oggetto di una indagine statistica, ovvero l insieme delle unità, dette unità statistiche o individui
Rispondere alle domande Vero/Falso e alle Domande a scelta multipla apponendo una crocetta sulla risposta giusta.
 Rispondere alle domande Vero/Falso e alle Domande a scelta multipla apponendo una crocetta sulla risposta giusta. Domande vero/ falso 1) Un individuo con genotipo AaBBcc può produrre due tipi di gameti
Rispondere alle domande Vero/Falso e alle Domande a scelta multipla apponendo una crocetta sulla risposta giusta. Domande vero/ falso 1) Un individuo con genotipo AaBBcc può produrre due tipi di gameti
Genetica dei caratteri complessi
 Genetica dei caratteri complessi Lezione 4 1 Definizione Eredità dei caratteri complessi Determinati caratteri vengono trovati con maggior frequenza in alcune famiglie, non mostrando però la normale segregazione
Genetica dei caratteri complessi Lezione 4 1 Definizione Eredità dei caratteri complessi Determinati caratteri vengono trovati con maggior frequenza in alcune famiglie, non mostrando però la normale segregazione
La trasmissione dei caratteri genetici. le leggi di Mendel
 La trasmissione dei caratteri genetici e le leggi di Mendel I figli assomigliano ai genitori perché hanno ereditato da loro alcune caratteristiche fisiche. Ma in che modo avviene questa trasmissione dei
La trasmissione dei caratteri genetici e le leggi di Mendel I figli assomigliano ai genitori perché hanno ereditato da loro alcune caratteristiche fisiche. Ma in che modo avviene questa trasmissione dei
MENDEL E LE SUE LEGGI
 MENDEL E LE SUE LEGGI Gregor Mendel (1822 1884) Era un monaco boemo considerato il padre della genetica, a cui si debbono le prime fondamentali leggi sull ereditarietà (1865). E importante ricordare un
MENDEL E LE SUE LEGGI Gregor Mendel (1822 1884) Era un monaco boemo considerato il padre della genetica, a cui si debbono le prime fondamentali leggi sull ereditarietà (1865). E importante ricordare un
LA GENETICA. La GENETICA (dal greco gennao, γενναω, dare vita, generare) è la scienza che studia le regole e i meccanismi dell ereditarieta biologica.
 è la scienza che studia le regole e i meccanismi dell ereditarieta biologica.") LA GENETICA La GENETICA (dal greco gennao, γενναω, dare vita, generare) è la scienza che studia le regole e i meccanismi dell ereditarieta biologica. L inizio della Genetica come scienza risale agli esperimenti
LA GENETICA La GENETICA (dal greco gennao, γενναω, dare vita, generare) è la scienza che studia le regole e i meccanismi dell ereditarieta biologica. L inizio della Genetica come scienza risale agli esperimenti
L EVOLUZIONE DELLA SPECIE. Le teorie di Darwin e le loro correlazioni con la genetica moderna
 L EVOLUZIONE DELLA SPECIE Le teorie di Darwin e le loro correlazioni con la genetica moderna Le Teorie di Darwin e l evoluzione della specie Charles Robert Darwin (1809 1882) è stato un naturalista britannico,
L EVOLUZIONE DELLA SPECIE Le teorie di Darwin e le loro correlazioni con la genetica moderna Le Teorie di Darwin e l evoluzione della specie Charles Robert Darwin (1809 1882) è stato un naturalista britannico,
Problemi di Genetica. Mendel, due loci. 1) La progenie di un testcross è la seguente:
 La progenie di un testcross è la seguente:") Problemi di Genetica Mendel, due loci 1) La progenie di un testcross è la seguente: 54 soggetti AaBb 57 soggetti Aabb 53 soggetti aabb 56 soggetti aabb su un totale di 220 Dimostrare che i loci presi in
Problemi di Genetica Mendel, due loci 1) La progenie di un testcross è la seguente: 54 soggetti AaBb 57 soggetti Aabb 53 soggetti aabb 56 soggetti aabb su un totale di 220 Dimostrare che i loci presi in
Università degli Studi di Firenze
 Dimensione delle popolazioni e consanguineità In popolazioni ad accoppiamento casuale incremento di consanguineità DF = 1 / (2N) Per generazione L equazione indica: incremento di consanguineità è inversamente
Dimensione delle popolazioni e consanguineità In popolazioni ad accoppiamento casuale incremento di consanguineità DF = 1 / (2N) Per generazione L equazione indica: incremento di consanguineità è inversamente
IL MIGLIORAMENTO GENETICO IN ZOOTECNIA
 IL MIGLIORAMENTO GENETICO IN ZOOTECNIA COS E Il miglioramento genetico (MG) degli animali zootecnici è la tecnica che consente l'aumento delle prestazioni produttive e riproduttive degli allevamenti attraverso
IL MIGLIORAMENTO GENETICO IN ZOOTECNIA COS E Il miglioramento genetico (MG) degli animali zootecnici è la tecnica che consente l'aumento delle prestazioni produttive e riproduttive degli allevamenti attraverso
Indice Prefazione Capitolo 1 Principi di zootecnica generale 1.1 Animali domestici e tassonomia 1.2 Produzione e riproduzione
 Indice Prefazione XI Capitolo 1 Principi di zootecnica generale (Pierlorenzo Secchiari) 1 1.1 Animali domestici e tassonomia 1 1.2 Produzione e riproduzione 3 1.2.1 Riproduzione naturale 6 1.2.2 Tecniche
Indice Prefazione XI Capitolo 1 Principi di zootecnica generale (Pierlorenzo Secchiari) 1 1.1 Animali domestici e tassonomia 1 1.2 Produzione e riproduzione 3 1.2.1 Riproduzione naturale 6 1.2.2 Tecniche
Analisi QTL. Include materiale dal capitolo 20.6 (pag ), vol III, Barcaccia e Falcinelli
, vol III, Barcaccia e Falcinelli") Analisi QTL Include materiale dal capitolo 20.6 (pag 984-1000), vol III, Barcaccia e Falcinelli Interpretazione della distribuzione continua dei caratteri nelle popolazioni naturali e sperimentali Tre
Analisi QTL Include materiale dal capitolo 20.6 (pag 984-1000), vol III, Barcaccia e Falcinelli Interpretazione della distribuzione continua dei caratteri nelle popolazioni naturali e sperimentali Tre
Corso di Genetica (canale M-Z)
") Corso di Genetica (canale M-Z) http://gentest.uniroma2.it/gentestmz/ per registrarsi username: password: studente studente-mz Corso di Laurea in Scienze Biologiche Genetica 1 Corso di Genetica (canale
Corso di Genetica (canale M-Z) http://gentest.uniroma2.it/gentestmz/ per registrarsi username: password: studente studente-mz Corso di Laurea in Scienze Biologiche Genetica 1 Corso di Genetica (canale
Matematicandoinsieme. Mendel e l ereditarietà
 Matematicandoinsieme Mendel e l ereditarietà 1 Gregor Mendel: il padre della genetica Gregor Mendel(1822-1894) Monaco e naturalista austriaco. Condusse i suoi studi sulle piante del pisello odoroso nel
Matematicandoinsieme Mendel e l ereditarietà 1 Gregor Mendel: il padre della genetica Gregor Mendel(1822-1894) Monaco e naturalista austriaco. Condusse i suoi studi sulle piante del pisello odoroso nel
MITOSI E MEIOSI. Esercizio n.1
 MITOSI E MEIOSI Esercizio n.1 Disegna in modo schematico i cromosomi nei diversi stadi di mitosi di una cellula diploide con un numero di cromosomi n = 1. Indica il cromosoma o il cromatidio con una linea,
MITOSI E MEIOSI Esercizio n.1 Disegna in modo schematico i cromosomi nei diversi stadi di mitosi di una cellula diploide con un numero di cromosomi n = 1. Indica il cromosoma o il cromatidio con una linea,
Caratteri: tra Geni e Ambiente
 Caratteri: tra Geni e Ambiente Perturbazioni casuali Fattori genetici Fattori ambientali ESPRESSIONE DEL CARATTERE 1 Caratteri: tra Geni e Ambiente Ambiente Genoma Ambiente Genoma Ambiente Genoma Ambiente
Caratteri: tra Geni e Ambiente Perturbazioni casuali Fattori genetici Fattori ambientali ESPRESSIONE DEL CARATTERE 1 Caratteri: tra Geni e Ambiente Ambiente Genoma Ambiente Genoma Ambiente Genoma Ambiente
La frequenza di crossing-over varia normalmente nei diversi segmenti del cromosoma, ma e un evento che si puo' facilmente prevedere fra due qualunque
 La frequenza di crossing-over varia normalmente nei diversi segmenti del cromosoma, ma e un evento che si puo' facilmente prevedere fra due qualunque loci genici. Quindi le effettive distanze fisiche fra
La frequenza di crossing-over varia normalmente nei diversi segmenti del cromosoma, ma e un evento che si puo' facilmente prevedere fra due qualunque loci genici. Quindi le effettive distanze fisiche fra
GRUPPI SANGUIGNI. Supponendo che la popolazione italiana sia H-W, calcola la probabilità di ogni singolo allele e di ogni genotipo
 GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
GRUPPI SANGUIGNI La distribuzione dei gruppi sanguigni nella popolazione italiana è: gruppo A 36%, gruppo B 17%, gruppo AB 7%, gruppo 0 40%. Il gruppo sanguigno è determinato da un locus genetico con tre
BIOTECNOLOGIE ANIMALI: LINKAGE E MARCATORI
 BIOTECNOLOGIE ANIMALI: LINKAGE E MARCATORI Associazione o Linkage Marcatori genetici Su ciascun cromosoma sono presenti più geni, ognuno ad un locus specifico Se i cromosomi venissero trasmessi da una
BIOTECNOLOGIE ANIMALI: LINKAGE E MARCATORI Associazione o Linkage Marcatori genetici Su ciascun cromosoma sono presenti più geni, ognuno ad un locus specifico Se i cromosomi venissero trasmessi da una
La L G e G n e et e ica Le leggi di Mendel
 La Genetica Le leggi di Mendel Il monaco Gregor Mendel (1822-1884) fu il primo a studiare in modo rigoroso il fenomeno della trasmissione dei caratteri ereditari. Per questo, pur non avendo nessuna conoscenza
La Genetica Le leggi di Mendel Il monaco Gregor Mendel (1822-1884) fu il primo a studiare in modo rigoroso il fenomeno della trasmissione dei caratteri ereditari. Per questo, pur non avendo nessuna conoscenza
ANALISI MENDELIANA. GENOTIPO: costituzione genetica di un individuo, sia riferito ad un singolo gene, sia all insieme dei suoi geni.
 ANALISI MENDELIANA ANALISI MENDELIANA GENOTIPO: costituzione genetica di un individuo, sia riferito ad un singolo gene, sia all insieme dei suoi geni. FENOTIPO: manifestazione fisica di un carattere genetico,
ANALISI MENDELIANA ANALISI MENDELIANA GENOTIPO: costituzione genetica di un individuo, sia riferito ad un singolo gene, sia all insieme dei suoi geni. FENOTIPO: manifestazione fisica di un carattere genetico,
I gameti prodotti saranno:
 GENI CONCATENATI Con i principi di Mendel e con lo studio della dinamica della meiosi due geni si trasmettono ciascuno in modo indipendente rispetto all altro se sono localizzati su paia di cromosomi diversi
GENI CONCATENATI Con i principi di Mendel e con lo studio della dinamica della meiosi due geni si trasmettono ciascuno in modo indipendente rispetto all altro se sono localizzati su paia di cromosomi diversi
Matematica Lezione 22
 Università di Cagliari Corso di Laurea in Farmacia Matematica Lezione 22 Sonia Cannas 14/12/2018 Indici di posizione Indici di posizione Gli indici di posizione, detti anche misure di tendenza centrale,
Università di Cagliari Corso di Laurea in Farmacia Matematica Lezione 22 Sonia Cannas 14/12/2018 Indici di posizione Indici di posizione Gli indici di posizione, detti anche misure di tendenza centrale,
GENETICA DEI CARATTERI COMPLESSI
 GENETICA DEI CARATTERI COMPLESSI 1 EPISTASI In genetica l'epistasi è il fenomeno per cui l'espressione fenotipica di un gene è soppressa o inibita da un secondo carattere ereditario. Il termine deriva
GENETICA DEI CARATTERI COMPLESSI 1 EPISTASI In genetica l'epistasi è il fenomeno per cui l'espressione fenotipica di un gene è soppressa o inibita da un secondo carattere ereditario. Il termine deriva
I ESERCITAZIONE GENETICA
 2-04-2008 _ Dott.Baratta I ESERCITAZIONE GENETICA 1) Definizione di Probabilità (eventi elementari) 2) Calcolo della Probabilità di eventi composti (Regola del prodotto e della somma) 3) Predizione dei
2-04-2008 _ Dott.Baratta I ESERCITAZIONE GENETICA 1) Definizione di Probabilità (eventi elementari) 2) Calcolo della Probabilità di eventi composti (Regola del prodotto e della somma) 3) Predizione dei
TAPPE IMPORTANTI DELLA GENETICA (1)
") TAPPE IMPORTANTI DELLA GENETICA (1) 1859 DARWIN Natura ereditaria della variabilità delle specie 1865 MENDEL Leggi che regolano la trasmissione dei caratteri 1902 GARROD Primo esempio umano di ereditarietà
TAPPE IMPORTANTI DELLA GENETICA (1) 1859 DARWIN Natura ereditaria della variabilità delle specie 1865 MENDEL Leggi che regolano la trasmissione dei caratteri 1902 GARROD Primo esempio umano di ereditarietà
I fondamenti genetici del lavoro di selezione
 I fondamenti genetici del lavoro di selezione Per un programma di miglioramento genetico occorrono: 1. Un idea precisa su quello che vogliamo ottenere: Definizione dell IDEOTIPO. I materiali idonei, che
I fondamenti genetici del lavoro di selezione Per un programma di miglioramento genetico occorrono: 1. Un idea precisa su quello che vogliamo ottenere: Definizione dell IDEOTIPO. I materiali idonei, che
GENETICA. Dott.ssa Priscilla Bettini http://www.dbag.unifi.it. http://www3.unifi.it/clscna/index.php
 GENETICA Dott.ssa Priscilla Bettini http://www.dbag.unifi.it http://www3.unifi.it/clscna/index.php INNOVAZIONE DI MENDEL Scelta del materiale: Pisum sativum pianta annuale si autofeconda caratteri ben
GENETICA Dott.ssa Priscilla Bettini http://www.dbag.unifi.it http://www3.unifi.it/clscna/index.php INNOVAZIONE DI MENDEL Scelta del materiale: Pisum sativum pianta annuale si autofeconda caratteri ben
Correzione primo compitino, testo A
 Correzione primo compitino, testo A gennaio 0 Parte Esercizio Facciamo riferimento alle pagine e del libro di testo Quando si ha a che fare con la moltiplicazione o la divisione di misure bisogna fare
Correzione primo compitino, testo A gennaio 0 Parte Esercizio Facciamo riferimento alle pagine e del libro di testo Quando si ha a che fare con la moltiplicazione o la divisione di misure bisogna fare
La genetica quantitativa e i caratteri multifattoriali. Prof. Alberto Pallavicini
 La genetica quantitativa e i caratteri multifattoriali Prof. Alberto Pallavicini pallavic@units.it Ereditarietà dei caratteri complessi Questa famosa immagine mostra l incredibile varietà in tratti come
La genetica quantitativa e i caratteri multifattoriali Prof. Alberto Pallavicini pallavic@units.it Ereditarietà dei caratteri complessi Questa famosa immagine mostra l incredibile varietà in tratti come