Laila Craighero: 4 crediti = 32 ore Sezione di Fisiologia umana
|
|
|
- Eva Elisabetta Fantoni
- 6 anni fa
- Visualizzazioni
Transcript
1 Laila Craighero: 4 crediti = 32 ore crh@unife.it Sezione di Fisiologia umana

2 Studio: sezione di Fisiologia umana, via Fossato di Mortara 19 (2 piano del chiostro)

3

4

5 Fisiologia Germann Stanfield Edizione: III / 2009 Pubblicato da Edises 53 Euro Sulla pagina web di questo testo, inoltre, potete trovare le animazioni di molti concetti che trattiamo a lezione:
6
7 TUTTE LE FIGURE PRESENTATE A LEZIONE POSSONO ESSERE DOMANDA D ESAME
8
9 CAPITOLO 7 Cellule nervose e segnali elettrici
10 IL SISTEMA NERVOSO E la sede della ricezione, elaborazione, e trasmissione delle informazioni relative a tutto il corpo. fornisce sensazioni sull'ambiente interno ed esterno integra le informazioni sensoriali coordina le attività volontarie e involontarie regola e controlla le strutture e gli apparati periferici Il tessuto nervoso comprende due distinte popolazioni cellulari: le cellule nervose o neuroni e le cellule di sostegno o neuroglia. La neuroglia isola i neuroni e fornisce una rete di sostegno. I neuroni sono responsabili del trasferimento e dell'elaborazione delle informazioni nel sistema nervoso.

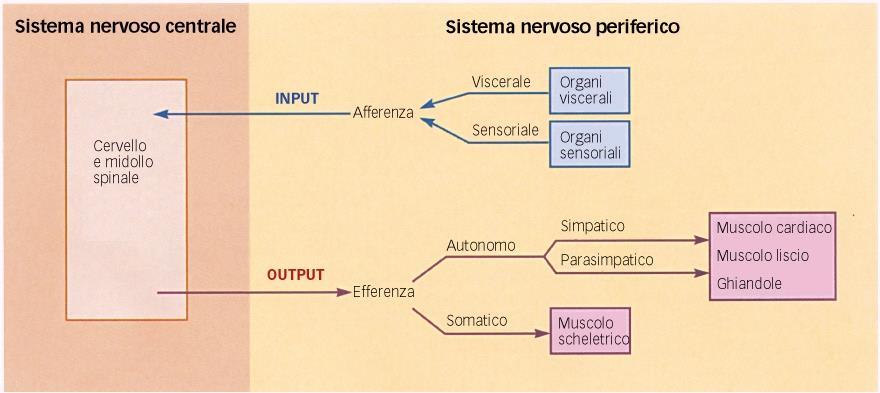
11 Il sistema nervoso viene diviso anatomicamente in: Sistema nervoso centrale (SNC): encefalo, racchiuso nella scatola cranica midollo spinale, contenuto invece nel canale vertebrale. Controlla autonomamente i riflessi. Il SNC è responsabile dell'integrazione, analisi e coordinazione dei dati sensoriali e dei comandi motori ed è la sede delle funzioni cognitive (intelligenza, memoria, apprendimento, linguaggio, ecc). A differenza del sistema nervoso periferico, il SNC non è solo in grado di raccogliere e trasmettere informazioni, ma anche di integrarle. Sistema nervoso periferico (SNP) è costituito da tutto il tessuto nervoso al di fuori del SNC. Svolge essenzialmente la funzione di trasmissione del segnale attraverso fasci di conduzione. I segnali in entrata (afferenti) da un'unità periferica (organo) o in uscita (efferenti) verso un'unità periferica, decorrono in fibre separate (assoni) che generalmente sono raggruppate in un fascio di conduzione unitario (nervo). Un nervo contiene esclusivamente assoni, cellule di Schwann e tessuto connettivo. I corpi delle cellule nervose sono raggruppati nei gangli del sistema nervoso periferico e nei nuclei del midollo spinale e del tronco encefalico.


12 Ganglio: raggruppamento di corpi cellulari, localizzato all esterno del SNC

13 NEURONI: cellule specializzate nella generazione e trasmissione di informazione sotto forma di impulsi elettrici. Tale informazione è alla base delle sensazioni, del comportamento, dei processi fisiologici e delle capacità cognitive.
14 I neuroni Un neurone tipico possiede un corpo cellulare, o soma, molte diramazioni, dendriti sensoriali, e un lungo assone che termina in una o più terminazioni sinaptiche. Ogni neurone deve adempiere cinque funzioni fondamentali: ricevere informazioni (input) dall'ambiente esterno o interno, oppure da altri neuroni; integrare le informazioni ricevute e produrre un'adeguata risposta in forma di segnale (output); condurre il segnale al suo terminale di uscita; trasmettere il segnale ad altre cellule nervose, ghiandole o muscoli; coordinare le proprie attività metaboliche, mantenendo l'integrità della cellula.
15 I dendriti Sono ramificazioni che si estendono dal corpo della cellula nervosa, specializzati nel rispondere ai segnali provenienti da altri neuroni o dall'ambiente esterno. La loro forma ramificata offre un'ampia superficie alla ricezione dei segnali. I dendriti dei neuroni sensoriali sono dotati di speciali adattamenti della membrana che consentono loro di rispondere a stimoli ambientali specifici come la pressione, gli odori, la luce o il calore. Nei neuroni del cervello e del midollo spinale, i dendriti rispondono ai neurotrasmettitori chimici liberati da altri neuroni. Essi sono dotati di recettori proteici di membrana che si legano a neurotrasmettitori specifici e inviano, come risultato di quel legame, segnali elettrici.
16 Il soma (corpo cellulare) Assicura le funzioni vitali del neurone e integra i segnali elettrici provenienti dai dendriti. Viaggiando lungo i dendriti, i segnali confluiscono al corpo cellulare del neurone che, comportandosi come un centro di integrazione, li "interpreta" e "decide" se produrre un potenziale d'azione, il segnale elettrico di uscita (output) del neurone. Provvisto dell'assortimento di organuli simile a quello di qualsiasi altra cellula, il corpo cellulare sintetizza anche proteine, lipidi e carboidrati, e coordina inoltre le attività metaboliche della cellula.
17 L'assone Trasporta a destinazione i segnali elettrici generati dal corpo cellulare. In un neurone tipico, l'assone, che è una fibra lunga e sottile, si protende dal corpo cellulare, facendo del neurone la cellula più lunga del corpo umano. Singoli assoni, per esempio, si estendono dal midollo spinale alle dita dei piedi, coprendo una distanza superiore a un metro.

18 I terminali sinaptici I terminali sinaptici comunicano con altri neuroni, muscoli e ghiandole. La maggior parte dei terminali sinaptici contiene una sostanza chimica specifica, detta neurotrasmettitore, che viene liberata in risposta a un potenziale d'azione che percorre l'assone. Il segnale in uscita (output) della prima cellula (presinaptica) diventa il segnale in entrata (input) per la seconda cellula (postsinaptica).


19 Nei neuroni del cervello e del midollo spinale, i dendriti rispondono ai neurotrasmettitori chimici liberati da altri neuroni. I dendriti dei neuroni sensoriali sono dotati di speciali adattamenti della membrana che consentono loro di rispondere a stimoli ambientali specifici come la pressione, gli odori, la luce o il calore.
20 I neuroni comunicano generando segnali elettrici sotto forma di modificazioni del potenziale di membrana Potenziale di membrana=differenza di potenziale elettrico ai due lati della membrana cellulare = le cariche elettriche sono distribuite in modo disomogeneo tra l esterno e l interno del neurone Nei sistemi biologici le cariche elettriche non sono date dagli elettroni ma dagli ioni ioni=atomi carichi elettricamente. Si possono combinare a fare dei composti es.: NaCl, cloruro di sodio, sale. Se il composto viene messo in acqua, si scinde dividendosi in ioni positivi e negativi. Il n di cariche positive è uguale a quello delle cariche negative. NaCl Le molecole non sono mai ferme (agitazione termica). Lo sarebbero solamente ad una temperatura vicina allo zero assoluto: -273 C, condizione che non si verifica mai! Non si staccano perché sono intrappolate dalla struttura cristallina. H2O NaCl Mettendo il cristallo in acqua, dove l acqua bagna la struttura reticolare, esso si modifica, le molecole di NaCl di staccano separandosi successivamente in Sodio e Cloro. Cl- Na+ NaCl Cl- Na+ Na+ Cl- Cl- Na+ Na+ Cl- Cl- Cl- Na+ Cl- Na+ Na+ Na+ Gli ioni continuano a muoversi allontanandosi, finché non sbattono contro il bordo. All inizio la concentrazione è maggiore vicino al cristallo (gradiente di concentrazione). La distribuzione è in termini di probabilità. Cl-
21 Membrana semipermeabile: passa l acqua (solvente) e non il sale (soluto). Stesso livello nei due recipienti. Più molecole d acqua Cl- NaCl Cl- Na+ Na+ Il solvente si muove dal compartimento in cui la soluzione è meno concentrata a quello dove la concentrazione è maggiore. H2O Na+ Na+ Cl- Cl- Na+ Na+ Cl- Cl- Cl- Na+ Cl- Na+ Na+ Cl- Meno molecole d acqua perché un po di spazio è occupato dal sale Le specie chimiche tendono a migrare dalla soluzione dove sono più concentrate a quella dove la loro concentrazione è minore. E più probabile che l acqua passi da più a meno, che il contrario: GRADIENTE DI CONCENTRAZIONE H2O Cl- Na+ NaCl Cl- Na+ Na+ Cl- Cl- Na+ Na+ Cl- FORZA CHIMICA Cl- Cl- Na+ Cl- Na+ Na+ Cl- Na+ Non si raggiungerà mai l equilibrio, perché il soluto c è sempre. Il livello di liquido si ferma quando il suo peso esercita una forza pari a quella che spinge l acqua a spostarsi
22 Carichiamo l acqua negativamente e con una pila mettiamo il polo positivo a sinistra e quello negativo a destra. + - Cariche dello stesso segno si respingono, mentre cariche di segno opposto si attraggono Cl- NaCl Cl- Na+ Na+ H2O- Na+ Na+ Cl- Cl- Na+ Na+ Cl- FORZA CHIMICA Cl- Cl- Na+ Cl- Na+ Cl- Na+ Se la forza chimica e la forza elettrica sono pari non c è più passaggio di ioni FORZA ELETTRICA + - H2O- NaCl Cl- Na+ Cl- Na+ Na+ Cl- Cl- Cl- Na+ Na+ Cl- Cl- Cl- Na+ FORZA CHIMICA Na+ Na+ Cl- Na+
23 I neuroni comunicano generando segnali elettrici sotto forma di modificazioni del potenziale di membrana Potenziale di membrana: differenza di potenziale elettrico ai due lati della membrana cellulare Le cariche elettriche sono distribuite in modo disomogeneo tra l esterno e l interno del neurone Nei sistemi biologici le cariche elettriche non sono date dagli elettroni ma dagli ioni (atomi carichi elettricamente). Nei neuroni gli ioni in gioco sono: Sodio (Na+) Potassio (K+) Dentro il neurone ci sono le proteine (Proteine -) che vengono sintetizzate all interno e che non possono uscire Il K+ può passare attraverso la membrana (membrana permeabile al K+). Il Na+ non può passare attraverso la membrana (membrana impermeabile al Na+)
24 CANALI IONICI La loro apertura o chiusura cambia la permeabilità della membrana cellulare per specifici ioni. Quando cambia la permeabilità, gli ioni si muovono attraverso la membrana e quindi CAMBIA LA DISTRIBUZIONE DELLE CARICHE ELETTRICHE TRA I DUE LATI DELLA MEMBRANA e quindi si modifica il potenziale di membrana. Canali ionici passivi Sono sempre aperti Canali ligando-dipendenti Si aprono o si chiudono in risposta al legame di un messaggero chimico ad uno specifico recettore presente sulla membrana plasmatica. Nei neuroni questi canali si trovano sulla membrana della cellula postsinaptica, il messaggero è detto neurotrasmettitore e viene liberato dalla cellula presinaptica. Canali voltaggio-dipendenti Si aprono o chiudono in risposta a modificazioni del potenziale di membrana. Quelli per il Na+ e il K+ si trovano prevalentemente nel monticolo assonale e sono necessari per l insorgenza e la propagazione del potenziale d azione. Quelli per il Calcio si trovano nel terminale assonico e si aprono quando il potenziale d azione arriva ad esso, permettendo l entrata del Calcio nel terminale assonico attivando il rilascio del neurotrasmettitore.
25 POTENZIALI DI EQUILIBRIO DEL SODIO E DEL POTASSIO
26 Cosa spinge uno ione a spostarsi? la sua concentrazione: tende ad andare dove ce n è di meno: FORZA CHIMICA la sua carica: tende ad andare verso la carica opposta: FORZA ELETTRICA



27 Si supponga ad esempio che inizialmente uno ione sia molto più concentrato da un lato della membrana e che non vi sia alcuna differenza di potenziale tra i due lati Inizialmente lo ione si muoverà in direzione del lato dove è meno concentrato. Tuttavia man mano che lo ione passa, andranno accumulandosi da quel lato sempre più cariche positive (che saranno invece sottratte dall altra parte) La differenza di potenziale andrà aumentando e inizierà a ri-sospingere lo ione dentro la cellula All equilibrio le due forze si equivarranno senza che si possa mai raggiungere né ugual concentrazione dai due lati, né neutralità elettrica Gradiente di concentrazione Gradiente elettrico
28 IPOTIZZIAMO UNA CELLULA PERMEABILE SOLO AL POTASSIO IN CUI, ALL INIZIO, IL POTASSIO E SOLO FUORI Proteinenon passano K+

29 Le proteine-canale gli permettono di passare attraverso la membrana Proteinenon passano K+
30 Il K+ è tutto fuori (vuole andare dentro perché dentro non ce n è) Forza chimica Ha carica positiva (vuole andare dentro perché dentro la carica è negativa) Forza elettrica La membrana gli permette di passare ENTRA Proteinenon passano K+ K+ passa Forza elettrica Forza chimica
31 Il K+ ENTRA Fino a quando entra? Fino a quando la sua differenza di concentrazione non è così forte da generare una forza chimica opposta a quella elettrica Proteinenon passano K+ Ha carica positiva (vuole andare dentro perché dentro la carica è negativa) Forza elettrica Forza chimica Dentro ce n è già molto e quindi vuole uscire Quando la forza elettrica e quella chimica sono uguali, il movimento netto di K+ = 0 Questo nel neurone avviene quando la differenza di potenziale è di -94 mv (potenziale di equilibrio per il potassio)
32 L interno della cellula rimane negativo perché la negatività delle proteine è maggiore della positività del potassio - Proteinenon passano K+
33 IPOTIZZIAMO UNA CELLULA PERMEABILE SOLO AL SODIO IN CUI, ALL INIZIO, IL SODIO E SOLO FUORI - Proteinenon passano Na+
34 Il Na+ è tutto fuori (vuole andare dentro perché dentro non ce n è) Forza chimica Ha carica positiva (vuole andare dentro perché dentro la carica è negativa) Forza elettrica La membrana gli permette di passare ENTRA - Na+ passa Na+ Forza elettrica Forza chimica
35 Il Na+ ENTRA Fino a quando entra? Fino a quando le cariche positive che porta dentro non generano una forza elettrica che spinge gli ioni sodio fuori, opponendosi alla forza chimica + Na+ Na+ Forza elettrica Forza chimica Quando la forza elettrica e quella chimica sono uguali, il movimento netto di Na+ = 0 Questo nel neurone avviene quando la differenza di potenziale è di +60 mv (potenziale di equilibrio per il sodio)
36 NEI NEURONI A RIPOSO LA MEMBRANA E PERMEABILE AL POTASSIO E QUASI TOTALMENTE IMPERMEABILE AL SODIO Il potenziale di membrana a riposo nei neuroni è -70 mv (e non -94mV,pot. d equilibrio del K+, perché un po di Na+ entra) K+ - Na+ non passa Forza elettrica Forza chimica
37 Il Na+ è tutto fuori (vuole andare dentro perché dentro non ce n è) Ha carica positiva (vuole andare dentro perché dentro la carica è negativa) La membrana NON gli permette di passare RESTA FUORI MA VORREBBE ENTRARE!! K+ - Na+ non passa Forza elettrica Forza chimica La situazione del neurone al potenziale di riposo NON E UNA SITUAZIONE DI EQUILIBRIO!

38
39 FORZA ELETTROCHIMICA: È la forza totale che agisce sugli ioni trasportati. Determina la direzione con la quale gli ioni si muoverebbero se venisse loro permesso di attraversare spontaneamente la membrana. Quando gli ioni vengono trasportati passivamente, si muovono sempre nella direzione della forza elettrochimica, cioè SI MUOVONO SECONDO IL LORO GRADIENTE ELETTROCHIMICO. Quando sono trasportati attivamente (es. pompa sodio-potassio che utilizza energia) si muovono in direzione opposta a quella della forza elettrochimica SI MUOVONO CONTRO IL LORO GRADIENTE ELETTROCHIMICO

40 A riposo, la separazione di cariche dovuta al fatto che l interno è negativo perché ci sono le proteine e l esterno è positivo perché c è il Na+, porta ad un potenziale di membrana a riposo di circa -70mV K+ - Na+ non passa
41 La pompa sodio-potassio impedisce che, a lungo andare, la permeabilità al potassio (alta) e al sodio (bassa) annulli i gradienti di concentrazione di potassio e sodio, «scaricando» il neurone. La pompa sodio-potassio è una pompa attiva perché utilizza energia (ATP) per trasportare ioni CONTRO il loro gradiente elettrochimico. Così facendo, mantiene il potenziale di membrana a riposo stabile a -70 mv.
42 THE MEMBRANE POTENTIAL permeabilità e pompa sodio-potassio
")

43 Se a un dendrita dei neuroni sensoriali arriva uno stimolo ambientale (pressione, odori, luce, calore) oppure un dendrita dei neuroni del cervello o del midollo spinale riceve un neurotrasmettitore da un altro neurone (sinapsi) cambia la permeabilità per uno ione = si aprono dei buchi che fanno passare quello ione passano ioni attraverso la membrana e quindi la separazione di cariche cambia e quindi il potenziale di membrana cambia K+ - + Entrano un po di cariche positive Na+
44
45 Lo stimolo può essere più o meno forte ossia può far passare più o meno cariche e quindi può modificare più o meno il potenziale di membrana POTENZIALE GRADUATO : la sua ampiezza dipende dall intensità dello stimolo K+ + Ne entrano - un po di più Na+
46 I neuroni comunicano tra di loro grazie a modificazioni del potenziale di membrana dovute all apertura o chiusura di canali: - elettrici (voltaggio-dipendenti) - chimici (ligando-dipendenti) - meccanici (meccano-sensitivi) Quando questi canali si aprono o si chiudono, si modifica la permeabilità di membrana per un certo ione, determinando una modifica del potenziale di membrana, che può essere di due tipi: 1) POTENZIALI GRADUATI Piccoli segnali elettrici che agiscono a breve distanza perché diminuiscono di intensità man mano che ci si allontana dal sito di stimolazione 2) POTENZIALI D AZIONE Ampi segnali elettrici che si propagano per lunghe distanze senza diminuire in ampiezza
47 POTENZIALI GRADUATI I potenziali graduati rappresentano piccole modificazioni del potenziale di membrana che si verificano quando i canali ionici si aprono o si chiudono in risposta ad uno stimolo che agisce sulla cellula. CHE STIMOLO? COS E? Lo stimolo che genera un potenziale graduato può essere un neurotrasmettitore che si lega ai recettori localizzati sulla membrana del neurone (nelle sinapsi tra neurone e neurone) oppure può essere uno stimolo sensoriale (stimolo pressorio o luminoso) che agisce su un recettore sensitivo o sulla terminazione periferica di un neurone. - l ampiezza della variazione del potenziale di membrana varia in funzione dello stimolo: è un fenomeno graduato. Uno stimolo debole produce un piccolo cambiamento del potenziale di membrana, mentre uno stimolo più intenso provoca una variazione di potenziale di maggiore ampiezza
.")
48 ESEMPIO DI GENERAZIONE DI POTENZIALE GRADUATO IN SEGUITO A STIMOLAZIONE SENSORIALE: Per esempio, basta deformare meccanicamente la membrana (fenomeno comune nei sensori della cute) e si aprono dei buchi che permettono il passaggio del Na+ (e quindi determinano una depolarizzazione della cellula perché il Na+ entra). Come un palloncino sgonfio che ha dei piccoli buchi: se lo stiriamo i buchi si allargano. Questo è un fenomeno graduato, posso avere depolarizzazioni minori o maggiori a seconda di quanta distorsione esercito sulla membrana.
49 POTENZIALI GRADUATI - Alcuni potenziali graduati generano depolarizzazione (eccitatori), altri causano iperpolarizzazione (inibitori)

50 POTENZIALI GRADUATI - I potenziali graduati generano un potenziale d azione se depolarizzano la membrana fino al valore di soglia Nei neuroni, il valore soglia è approssimativamente -55mV
51 POTENZIALI GRADUATI - Il potenziale graduato si attenua con la distanza: la variazione del potenziale di membrana genera un flusso di corrente che si propaga ad aree adiacenti della membrana (conduzione elettrotonica=movimento passivo di cariche), ma parte della corrente attraversa la membrana determinando una diminuzione della variazione del potenziale di membrana. Quando l acqua scorre attraverso un manicotto collegato a un rubinetto, se il manicotto è intatto la quantità di acqua che entra è uguale a quella che esce; se invece vi è una piccola perdita (come nella membrana che perde ioni attraverso i canali passivi, sempre aperti) la fuoriuscita di acqua risulterà minore. All allontanarsi del potenziale graduato dal sito di stimolazione, la corrente scorre (grazie alla conduzione elettrotonica) in un ampia area di membrana e una sua parte attraversa la membrana e si disperde. Quindi, l ampiezza della variazione del potenziale di membrana diminuisce all aumentare della distanza dal punto di stimolazione.

52 La corrente che esce dalla membrana viene persa. Per questo motivo la modificazione del potenziale di membrana diminuisce allontanandosi dal sito di stimolazione.
53 Legge di Ohm: I = E/R e propagazione del potenziale graduato I = quantità di cariche elettriche che passa (corrente) E = differenza di potenziale R = resistenza La quantità di cariche elettriche che passa aumenta all aumentare della differenza di potenziale e diminuisce all aumentare della resistenza La RESISTENZA dipende: dal DIAMETRO dell assone. Più è piccolo, più fa fatica a passare. Dalla RESISTENZA DI MEMBRANA: la membrana non è completamente isolata (ci sono dei buchi) e quindi un po di ioni escono e si disperdono. Il potenziale graduato può propagarsi dal punto di stimolazione per brevi distanze in quanto si attenua con la distanza dal punto in cui si genera. La distanza a cui si propaga il potenziale graduato dipende dalla Costante di spazio: aumenta all aumentare della resistenza di membrana (se ci sono pochi buchi) diminuisce all aumentare della resistenza dell assone (se è piccolo) La conduzione passiva del potenziale graduato è una conduzione elettrotonica che segue le stesse regole di qualsiasi conduttore.
54 Costante di spazio: aumenta all aumentare della resistenza di membrana (se ci sono pochi buchi) diminuisce all aumentare della resistenza dell assone (se è piccolo) a) Res. Membrana: + Res. Assone: - b) Res. Membrana: - Res. Assone: + Maggiore distanza percorsa nel caso a)

55 Un singolo potenziale graduato non è quasi mai di ampiezza sufficiente a generare un potenziale d azione. Se i singoli potenziali graduati si sovrappongono si possono sommare: SOMMAZIONE TEMPORALE: nello stesso punto vengono applicati in rapida successione più stimoli. Il tempo tra uno stimolo e l altro deve essere così breve da sovrapporre le depolarizzazioni dei diversi stimoli. SOMMAZIONE SPAZIALE: si sommano gli effetti dei potenziali graduati che nascono in diverse regioni della membrana.
56 Quando il potenziale graduato raggiunge il livello di soglia (-55mV) si aprono i CANALI VOLTAGGIO-DIPENDENTI PER IL Na+ - - K+ Na+ LIVELLO SOGLIA
57 Quando il potenziale graduato raggiunge il livello di soglia (-55mV) si aprono i CANALI VOLTAGGIO-DIPENDENTI PER IL Na+ e il Na+ ENTRA A VALANGA perché è positivo (e l interno è negativo) e il Na+ è tutto fuori - - K+ Na+ Na+ LIVELLO SOGLIA

58 Quando la depolarizzazione determinata da molti potenziali graduati raggiunge il monticolo assonale il potenziale d azione viene generato Il potenziale d azione viene generato nel monticolo assonale perché in esso è presente un gran numero di canali voltaggio dipendenti per il Na+


59 Il processo di depolarizzazione è rigenerativo
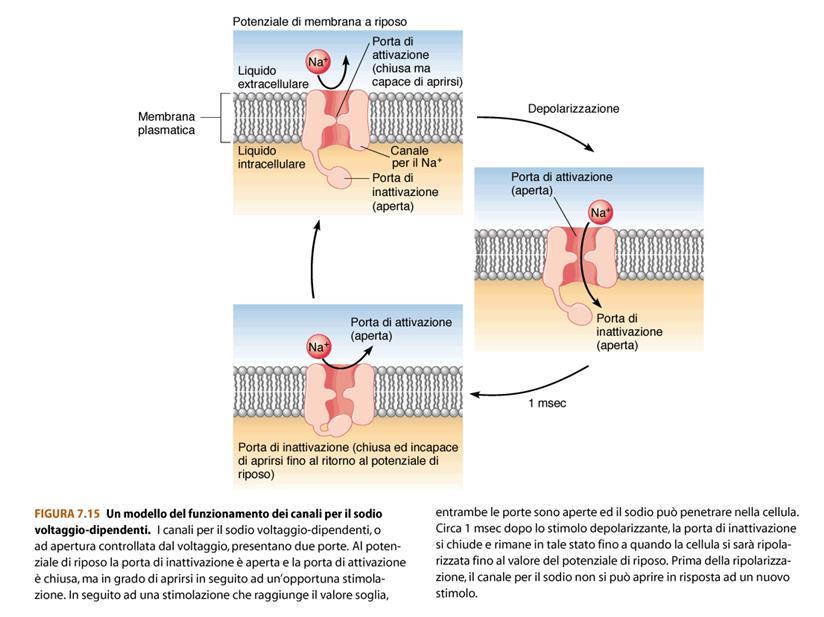

60 CANALI VOLTAGGIO-DIPENDENTI PER IL SODIO
61 In un tempo brevissimo il Na+ porta all interno della cellula molte cariche positive, invertendo in questo modo il potenziale di membrana che passa da -70mV a +30mV K+ Na+ ENTRA +
62 A questo punto: si chiudono i canali voltaggio-dipendenti per il Na+ (il Na+ non entra più, non entrano più cariche positive) e si aprono i canali voltaggio-dipendenti per il K+. Il K+ esce portando fuori cariche positive (il K+ è tutto dentro e l interno è positivo!) Il potenziale di membrana ritorna negativo. + Na+ NON ENTRA PIU K+ ESCE
 e così viene ripristinata la situazione di partenza K+ - Na+ non")
63 Grazie alla pompa sodio-potassio (che utilizza energia) viene portato fuori il Na+ (3 ioni Na+) e dentro il K+ (2 ioni K+) e così viene ripristinata la situazione di partenza K+ - Na+ non passa
64 Pompa Na/K permette di ripristinare le condizioni di partenza. Pompa Na/K utilizza energia. In biologia, spesso, l energia viene utilizzata per ripristinare le condizioni di partenza. Possiamo salire le scale (passare da un punto a minor energia potenziale ad un punto a maggior energia potenziale -per rompere un uovo posso lasciarlo cadere) lentamente o velocemente. Potenza = tempo durante il quale una forza viene applicata (Watt = forza/tempo) minore è il tempo, maggiore è la potenza che devo usare per esercitare la stessa forza (forza = potenza X tempo) Per esercitare una forza sufficiente a sollevare un chilo di peso in 10 msec avrei bisogno di una grande potenza. Per salire le scale in un tempo velocissimo posso sedermi su una catapulta che carico grazie ad una manovella per un quarto d ora (immagazzino energia potenziale) e poi faccio scattare il meccanismo che mi catapulta in cima alle scale. Il neurone deve generare il potenziale d azione in un tempo brevissimo: quindi carica il meccanismo e poi fa scattare il grilletto.
65 THE ACTION POTENTIAL
66 APERTURA CANALI VOLTAGGIO-DIPENDENTI PER IL Na+ CHE ENTRA A VALANGA CHIUSURA CANALI VOLTAGGIO-DIPENDENTI PER IL Na+ APERTURA CANALI K+ CHE ESCE CELLULA POLARIZZATA SEPARAZIONE DI CARICHE Dentro negative/fuori positive RITARDO NELLA CHIUSURA DEI CANALI K+ il K+ esce troppo MODIFICAZIONE DEL POTENZIALE DI MEMBRANA Dovuta ad un cambio di permeabilità

67

68
69 Il POTENZIALE D AZIONE è un fenomeno ATTIVO perché ha bisogno di energia per ripristinare le condizioni di partenza è un fenomeno tutto-o-nulla c è o non c è. L ampiezza del potenziale è determinata dal potenziale di equilibrio per il sodio. Non è un potenziale graduato è autorigenerativo quando in un punto della membrana nasce un pot d az esso si propaga per tutta la membrana eccitando i punti vicini della membrana. Il potenziale d azione si propaga per conduzione elettrotonica ai punti adiacenti della membrana (come il potenziale graduato) DEPOLARIZZANDO SEMPRE I PUNTI ADIACENTI DELLA MEMBRANA FINO AL VALORE SOGLIA, SCATENANDO IN ESSI UN NUOVO POTENZIALE D AZIONE E alla base della possibilità di condurre il pot d az lungo i neuroni (un assone di motoneurone spinale che innerva la mano è lungo 1 metro!)

70 Il potenziale d azione si propaga sia in una direzione che nell altra. La depolarizzazione è bidirezionale e quindi, se non ci fosse un sistema di protezione, lo stesso punto continuerebbe ad essere sottoposto a potenziali d azione che tornano sempre indietro.
71 PERIODO REFRATTARIO ASSOLUTO Durante l inizio della fase di ripolarizzazione molti canali per il Na+ sono aperti e la gran parte delle porte di inattivazione dei canali per il Na+ è in posizione di chiusura. QUINDI: non è possibile modificare la permeabilità della membrana per il Na+. Quindi NON è possibile evocare un nuovo potenziale d azione. PERIODO REFRATTARIO RELATIVO Solo uno stimolo molto intenso riesce a generare un secondo pot d az perché molte porte di inattivazione sono ancora chiuse, inoltre molti canali voltaggiodipendenti per il K+ sono aperti e quindi abbiamo molte cariche positive che escono. Pochi sono i canali Na+ che possono aprirsi.
72
73 I pot d az non possono sommarsi tra loro a causa del periodo di refrattarietà assoluto (impedisce la loro sovrapposizione). La grandezza dei potenziali graduati fornisce l informazione sull intensità dello stimolo. Come fanno i pot d az a informare sull intensità dello stimolo?
74 CODICE DI FREQUENZA Più lo stimolo è intenso, maggiore è il numero di potenziali d azione nell unità di tempo perché: più lo stimolo è intenso, prima è possibile evocare un pot d az durante il periodo refrattario relativo L intensità dello stimolo si ottiene mediante una codifica in frequenza di scarica dei pot d az. Poiché un potenziale graduato dura di più di un pot d az, esso può generare una scarica di pot d az. A seconda dell ampiezza di un potenziale graduato, i pot d az possono essere più o meno distanziati nel tempo. Stimolo soglia che dura più del periodo di refr ass: nuovo pot d az Stimolo soprasoglia: può generare un secondo pot d az anche durante il periodo di refr rel

75 Una volta che il pot d az è generato in un assone, esso si propaga senza decremento lungo l assone partendo dal monticolo assonale (o cono d emergenza) e giungendo alle terminazioni dell assone. Il pot d az viaggia generando gradienti elettrochimici nei liquidi intra ed extra cellulari. Tali liquidi permettono alle cariche positive (+) di muoversi dall area della membrana depolarizzata a quella adiacente (-), che viene a sua volta depolarizzata. LA CORRENTE CHE DIFFONDE NELLE ADIACENZE E SEMPRE DI SUFFICIENTE INTENSITA DA DEPOLARIZZARE LA MEMBRANA FINO AL VALORE SOGLIA, GENERANDO UN ALTRO POT D AZ Il primo potenziale d azione prodotto nel monticolo assonale genera una corrente che causa l insorgenza di un secondo potenziale d az nelle zone adiacenti della membrana. A sua volta questo produce un flusso di corrente che causa l insorgenza di un terzo pot d az, e così via fino a giungere al terminale assonico.
76 Legge di Ohm: I = E/R e propagazione del potenziale graduato I = quantità di cariche elettriche che passa (corrente) E = differenza di potenziale R = resistenza La quantità di cariche elettriche che passa aumenta all aumentare della differenza di potenziale e diminuisce all aumentare della resistenza La RESISTENZA dipende: dal DIAMETRO dell assone. Più è piccolo, più fa fatica a passare. Dalla RESISTENZA DI MEMBRANA: la membrana non è completamente isolata (ci sono dei buchi) e quindi un po di ioni escono e si disperdono.

77 Le cellule di Schwann creano una barriera che chiude i buchi attraverso i quali si verifica la dispersione degli ioni (aumenta la resistenza di membrana). È necessario che questa barriera venga interrotta ogni tanto per permettere il verificarsi di un nuovo pot d az. Infatti, il potenziale lungo l assone ricoperto di mielina viene condotto passivamente e tale conduzione risente della resistenza dell assone (resistenza intracellulare). Questo fa capire perché all interno delle famiglie ricoperte da mielina, la velocità di conduzione dipende dal diametro.
78 A livello delle regioni nodali, la membrana presenta un elevata concentrazione di canali voltaggio-dipendenti per il NA+ e il K+. La corrente depolarizzante che nasce all interno dell assone nella regione di un nodo di Ranvier, è sufficiente a depolarizzare la membrana a livello del nodo di Ranvier adiacente fino al valore di soglia, generando così un potenziale d azione. I potenziali d azione nascono in successione ad ogni nodo di Ranvier, propagandosi fino a giungere al terminale dell assone. Il salto del pot d az da un nodo all altro è il motivo per cui la conduzione negli assoni mielinici viene chiamata conduzione saltatoria.

79

80
81
82 CAPITOLO 8 Trasmissione sinaptica ed integrazione neuronale
83 I neuroni comunicano tra di loro attraverso strutture specializzate: le sinapsi. Sinapsi elettriche: non utilizzano neurotrasmettitori. Il segnale elettrico si propaga direttamente da una cellula all altra. Permettono una rapida comunicazione tra neuroni adiacenti sincronizzandone l attività elettrica. Sinapsi chimiche: un neurone secerne un neurotrasmettitore nello spazio extracellulare in risposta ad un potenziale d azione che arriva alla sua terminazione sinaptica il neurotrasmettitore si lega ad un recettore presente nella membrana cellulare di una seconda cellula tale legame favorisce o inibisce l insorgenza di un potenziale d azione nella seconda cellula

84 Nel SNC le sinapsi avvengono tra due neuroni. Non necessariamente la depolarizzazione indotta dal neurotrasmettitore raggiunge la soglia per la generazione del pot d az.

85 SINAPSI Il Ca++ determina la fusione delle vescicole sinaptiche con la superficie interna della membrana presinaptica, determinando l esocitosi e cioè il rilascio del neurotrasmettitore nella fessura sinaptica
86 LA QUANTITA DI NEUROTRASMETTITORE RILASCIATO DIPENDE DALLA CONCENTRAZIONE DI CALCIO NEL CITOPLASMA CHE, A SUA VOLTA, DIPENDE DALLA FREQUENZA DEI POTENZIALI D AZIONE NEL NEURONE PRESINAPTICO In mancanza dell arrivo di un ulteriore pot d az, il rilascio del neurotrasmettitore si arresta in pochi millisecondi, poiché i canali voltaggiodipendenti per il Ca++ restano aperti pochissimo tempo e il Ca++ viene subito attivamente pompato all esterno Se arriva un secondo potenziale PRIMA DELL ALLONTANAMENTO DEL NEUROTRASMETTITORE DALLA FESSURA SINAPTICA, questo determinerà un ulteriore rilascio di neurotrasmettitore (ntr) e, di conseguenza, un aumento di ntr nello spazio sinaptico. La concentrazione di neurotrasmettitore nella fessura sinaptica aumenta proporzionalmente alla frequenza dei pot d az. (SOMMAZIONE TEMPORALE)


87 Sinapsi eccitatoria Sinapsi inibitoria Il neurotrasmettitore interagisce con specifici recettori di membrana, generando una risposta nel neurone postsinaptico. Una volta liberato il ntr può agire solo per pochi millisecondi perché il suo legame con il recettore dura poco e una serie di processi lo allontana rapidamente dallo spazio sinaptico

88 Potenziale postsinaptico (PPS): variazione del potenziale di membrana della cellula postsinaptica indotta dall aumento di permeabilità ad uno o più ioni in conseguenza al legame tra neurotrasmettitore e recettore. Il PPS si forma rapidamente ed altrettanto rapidamente scompare (pochi millisecondi) poiché il canale-recettore si chiude appena il neurotrasmettitore si stacca da esso. PPS eccitatorio (PPSE): il potenziale di membrana raggiunge livelli più vicini al valore soglia (la cellula postsinaptica viene depolarizzata). I PPSE sono potenziali graduati la cui ampiezza depolarizzante cresce in funzione del numero di molecole di neurotrasmettitore che si legano al recettore. Il recettore-canale che si apre permette al Na+ a al K+ di passare. Il Na+ tende ad entrare MOLTO, il K+ ad uscire POCO. Quindi, il flusso verso l interno di Na+ prevale su quello verso l esterno di K+. La depolarizzazione è di ampiezza maggiore nel punto di origine e decresce con la distanza.
 o per il cloro (Cl- che entra dentro")
89 PPS inibitorio (PPSI): il potenziale di membrana raggiunge livelli più lontani dal valore soglia (la cellula postsinaptica viene iperpolarizzata). Il legame del neurotrasmettitore con il recettore apre un canale per il potassio, che fuoriesce dalla cellula iperpolarizzandola (rendendo l interno più negativo) o per il cloro (Cl- che entra dentro la cellula). Diminuisce la probabilità che si generi un pot d az I PPSI sono potenziali graduati la cui ampiezza cresce in funzione del numero di molecole di neurotrasmettitore che si legano al recettore. L iperpolarizzazione è di ampiezza maggiore nel punto di origine e decresce con la distanza.
90 A livello del monticolo assonale viene effettuata la sommazione algebrica dei PPSE e dei PPSI: integrazione neuronale. Quando la sommazione dei PPSE prevale sui PPSI ed è in grado di portare il potenziale di membrana a livello del monticolo assonale al valore soglia, nasce un pot d az.
91 SOMMAZIONE TEMPORALE Si può verificare in quanto i PPS durano di più dei pot d az Un secondo pot d az determina liberazione di neurotrasmettitore quando le molecole di ntr liberate dal primo pot d az sono ancora presenti nella fessura sinaptica: si aprono più canali. Se il secondo pot d az arriva dopo un po di tempo, il ntr liberato dal primo pot d az è già stato rimosso e quindi i loro effetti sui canali non si possono sommare. SOMMAZIONE SPAZIALE Si verifica in quanto i PPS, originati in differenti sinapsi, diffondono verso il monticolo assonale sommandosi tra loro. Se i neuroni A e B sono attivati nello stesso momento, i risultanti PPSE si sommano producendo una depolarizzazione sufficiente ad innescare un pot d az Se i neuroni A e B generano un pot d az in tempi diversi, ciascuno di essi induce un PPSE, ma nessun PPSE è sufficientemente ampio da generare un pot d az nella cellula postsinaptica.

92 In realtà i dendriti ed il corpo cellulare di un neurone postsinaptico possono ricevere input da centinaia o centinaia di migliaia di differenti neuroni: CONVERGENZA Alcune sinapsi sono eccitatorie, altre inibitorie e quindi il numero di possibili combinazioni di input sinaptici è astronomico.
93 SYNAPTIC POTENTIALS AND CELLULAR INTEGRATION
94 Sinapsi asso-assoniche: il ntr rilasciato dal neurone presinaptico si lega al recettore del neurone postsinaptico inducendo una modificazione del quantitativo di Ca++ che entra nel terminale assonico in risposta ad un pot d az. Questo provoca una modificazione della quantità di ntr che viene liberato: facilitazione presinaptica: il neurone postsinaptico libera più ntr inibizione presinaptica: il neurone postsinaptico libera meno ntr
95 Nel sistema nervoso somatico il ntr è l Ach. Nel sistema nervoso centrale i ntr vengono classificati in eccitatori ed inibitori e sono prevalentemente costituiti da aminoacidi: ntr eccitatori aspartato glutammato ntr inibitori glicina GABA acido gamma-aminobutirrico Il GABA è un ntr inibitorio, pur avendo caratteristiche chimiche simili al glutammato da cui deriva. Questo dipende dal fatto che questi due ntr si legano a recettori completamente diversi tra loro. L azione di qualsiasi ntr dipende sempre dal tipo di recettore con il quale interagisce e non dalla sua struttura chimica.
96 Benzodiazepine: (sonniferi) hanno dei recettori specifici. L interazione delle BDZ con i propri siti di legame facilita l interazione del GABA con il proprio recettore determinando un aumento della FREQUENZA di apertura del canale allo ione Cl- con aumento della permeabilità della membrana per questo ione. Questo determina inibizione neuronale per iperpolarizzazione con Azione ansiolitica o ipnoinducente Azione miorilassante Azione anticonvulsivante e sedativa. Barbiturici: I barbiturici aumentano le capacità del GABA di inibire la neurotrasmissione (meccanismo condiviso con le BZP) attraverso il prolungamento del tempo di apertura del canale cloro nel complesso recettore-gaba con effetto iperpolarizzante che causa l'inibizione. A differenza delle benzodiazepine i barbiturici possono aprire il canale cloro anche in assenza di GABA; questo avviene a concentrazioni più alte di quelle che occorrono per potenziare l'azione del GABA. Sono estremamente pericolosi in quanto bloccano l eccitabilità della membrana: si può morire.
97 ANATOMY REVIEW: CHEMICAL SYNAPSE AND ELECTRICAL SYNAPSE
98 CAPITOLO 11 Da pag. 314 Il sistema nervoso somatico
99 CAPITOLO 12 Fisiologia del muscolo
100 I MUSCOLI Con poche eccezioni i muscoli scheletrici sono collegati ad almeno due ossa. E sono ancorati alle ossa per mezzo dei tendini, corde di tessuto connettivale elastico che trasmettono la forza dal muscolo all osso. Il muscolo è composto da molti fascicoli: Ciascun fascicolo è costituito da cellule muscolari, tessuto connettivo, vasi sanguigni e nervi. Cellule muscolari (fibre muscolari): hanno molti nuclei perché derivano embriologicamente dalla fusione di diverse cellule che si trovano subito sotto la membrana citoplasmatica della fibra muscolare (sarcolemma). Il citoplasma contiene mitocondri (dove viene prodotta l'energia necessaria allo svolgimento delle funzioni vitali della cellula) e centinaia di elementi a forma di bacchetta, le miofibrille. Ogni miofibrilla è costituita da un fascio di filamenti sovrapposti spessi e sottili, formati rispettivamente dalle proteine miosina e actina. Ciascuna miofibrilla è avvolta da una rete membranosa, il reticolo sarcoplasmatico, che ha il ruolo di magazzino del Ca++. Il reticolo sarcoplasmatico è strettamente connesso ai tubuli trasversali (tubuli a T) che sono delle introflessioni del sarcolemma all interno del citoplasma.

101

102 SISTEMA NERVOSO SOMATICO un singolo motoneurone collega il sistema nervoso centrale al muscolo scheletrico FIGURA FIGURA UNITA MOTORIA L unità motoria è costituita da un motoneurone e da tutte le cellule muscolari da esso innervate. Un singolo motoneurone innerva molte fibre muscolari. Una singola fibra muscolare è innervata da un singolo motoneurone. Quando un motoneurone è attivato, induce la contrazione di tutte le fibre muscolari che innerva.
103 Ciascuna diramazione dell assone di un motoneurone forma sinapsi con una fibra muscolare scheletrica a livello di una regione altamente specializzata della membrana (chiamata PLACCA MOTRICE: numerose invaginazioni che presentano un grande numero di recettori per il neurotrasmettitore), formando la GIUNZIONE NEUROMUSCOLARE (= sinapsi tra motoneurone e muscolo) I terminali assonici del motoneurone (BOTTONI SINAPTICI) immagazzinano e rilasciano ACETILCOLINA (Ach), che è l unico neurotrasmettitore periferico del sistema nervoso somatico.

104 Il meccanismo della trasmissione neuromuscolare è uguale a quello che si verifica nelle sinapsi eccitatorie tra cellule nervose. Quando un motoneurone è attivato dalla convergenza su di esso di molti segnali eccitatori, si genera un potenziale d azione che si propaga fino a tutti i bottoni sinaptici che formano le giunzioni neuromuscolari con le fibre dell unità motoria.
105 Quando il pot d az arriva al bottone presinaptico apertura canali Ca++ voltaggio-dipendenti presenti nel bottone sinaptico Il Ca++ entra L aumento di concentrazione intracellulare di Ca++ permette l esocitosi (le membrana delle vescicole si fonde con la superficie interna della membrana cellulare) delle vescicole di Ach (il magnesio impedisce la fusione: poco magnesio=crampi) L acetilcolina diffonde nella fessura sinaptica ed interagisce con i recettori-canale colinergici nicotinici della placca motrice, determinandone l apertura. Questo permette al Na+ di entrare nella fibra muscolare producendo una depolarizzazione che prende il nome di POTENZIALE DI PLACCA, simile al PPSE, ma il potenziale di placca (a differenza del PPSE che è un potenziale graduato depolarizzante) è sempre di ampiezza sufficiente a depolarizzare la membrana della fibra muscolare fino al valore soglia, generando sempre, quindi, nella fibra muscolare, un potenziale d azione Il potenziale d azione generato nella fibra muscolare determinerà la contrazione della stessa fibra
106 L INNERVAZIONE DEI MUSCOLI SCHELETRICI DA PARTE DEI MOTONEURONI E SOLO ECCITATORIA (e provoca la contrazione). QUINDI IL RILASCIAMENTO DEL MUSCOLO SI VERIFICA SOLO QUANDO CESSA LA SCARICA DEI MOTONEURONI E CESSANO TUTTE LE CONSEGUENZE DETERMINATE DALL ARRIVO DEL POTENZIALE D AZIONE (aumento della concentrazione di Ca++ nel bottone sinaptico e presenza di acetilcolina nella fessura sinaptica) è necessario rimuovere l Ach: essa viene rimossa lentamente per diffusione ma acetilcolinesterasi: enzima presente sulla membrana del muscolo che permette l esterificazione dell Ach, dividendola in acido acetico (diffusione) e colina (aminoacido) che è molto preziosa e quindi viene recuperata dal bottone presinaptico per andare a formare nuova Ach Bottone presinaptico: se molte vescicole si fondono con la membrana del neurone queste aggiungono continuamente nuovi pezzetti alla membrana. Meccanismo di endocitosi che recupera la vescicola vuota. Questa verrà poi riempita nuovamente di Ach.

107 L assone è rivestito da una guaina mielinica che viene persa quando il motoneurone si avvicina al muscolo. Il motoneurone si sfiocca, separandosi in diverse terminazioni, ognuna delle quali arriva in prossimità del muscolo. Bottone presinaptico: presenti molte vescicole che contengono acetilcolina. Membrana postsinaptica: presenta numerose invaginazioni che hanno lo scopo di aumentare la superficie sinaptica davanti al bottone sinaptico. Fessura sinaptica
108 L Ach liberata dal bottone presinaptico interagisce con i recettori-canale nicotinici della placca neuromuscolare, determinandone l apertura. Questo permette al sodio di entrare nella cellula muscolare producendo una depolarizzazione che prende il nome di potenziale di placca. Tale potenziale è sempre di ampiezza sufficientemente grande da depolarizzare la membrana muscolare fino al valore soglia. Si genera così un pot d az che attiva la contrazione muscolare. Sicurezza della trasmissione sinaptica tra nervo e muscolo: i motoneuroni sono l ultima cellula nervosa prima che il segnale si trasformi in movimento. Potenzialmente il movimento può essere pericoloso e quindi l informazione deve essere precisa. Il SN deve assumere che ogni volta che il motoneurone si depolarizza, questo si traduce in movimento. Ecco perché il potenziale di placca è sufficientemente ampio da determinare sempre un pot d az.
109 THE NEUROMUSCULAR JUNCTION
110 La liberazione di Ach si ha anche spontaneamente (non solo in conseguenza all arrivo del pot d az) quando la vescicola casualmente va a sbattere contro la membrana e si verifica esocitosi. POTENZIALI DI PLACCA IN MINIATURA: modificazioni del pot di membrana delle cellule muscolari non sufficienti a determinare un pot d az. (dovuti ad una fusione casuale di una o più vescicole di acetilcolina con la membrana del motoneurone) Queste modificazioni sono sempre multipli di 0,4 mv. Da tali dati appare evidente che una vescicola sinaptica coinvolta nella trasmissione sinaptica neuromuscolare deve contenere un quantitativo di Ach ( quanto ) in grado di produrre una depolarizzazione di 0,4 mv in una cellula muscolare scheletrica. L arrivo di un pot d az nel terminale sinaptico a determina la liberazione di circa quanti di Ach.
111 I motoneuroni agiscono sul muscolo esclusivamente con sinapsi eccitatorie. Il rilasciamento del muscolo si verifica quando cessa la scarica dei motoneuroni. Alterata trasmissione neuromuscolare: non si ha contrazione muscolare patologia: miastenia gravis malattia autoimmune: il sistema immunitario produce anticorpi contro i recettori nicotinici dell acetilcolina nelle giunzioni neuromuscolari, determinando la distruzione dei recettori dell Ach e, di conseguenza, l incapacità delle cellule muscolari di contrarsi. Veleni: veleno del ragno vedova nera: contiene latrossina: induce rilascio di Ach con conseguenti spasmi muscolari e rigidità, che possono causare paralisi dei muscoli respiratori e morte. Veleno del serpente a sonagli: contiene crotossina: induce il blocco del rilascio di Ach, determinando una paralisi flaccida dei muscoli scheletrici. Curaro: veniva usato dagli indiani del Sud America per rendere velenose le frecce. Contiene tubocurarina che si lega ai recettori nicotinici, impedendo così il legame dell Ach.

112 SARCOMERO: modulo che si ripete più volte dentro cui sono comprese le miofibrille Sono ancorati i filamenti sottili Sono uniti tra loro i filamenti spessi 1 filamento spesso è circondato da 6 filamenti sottili BANDA A: filamenti spessi sovrapposti a quelli sottili ZONA H: solo filamenti spessi BANDA I: solo filamenti sottili

113 Filamento sottile: E costituito da molecole di actina concatenati a formare un polimero filamentoso costituito da due catene di actina avvolte ad elica. Ogni molecola possiede un sito per legare la miosina. Proteine regolatrici: tropomiosina molecola filamentosa che si colloca sopra numerose molecole di actina in modo da bloccare i siti di legame con la miosina. Troponina complesso di tre proteine una si attacca al filamento di actina una si fissa alla tropomiosina una contiene un sito che si può legare in maniera reversibile agli ioni calcio

114 Filamento spesso: E costituito da centinaia di molecole di miosina, accoppiate a due a due e avvolte l una all altra, che terminano con una estroflessione (ponti trasversali). Le molecole di miosina si fissano le une alle altre in corrispondenza della parte terminale della coda, facendo in modo che le due teste si dispongano con direzione opposta rispetto al centro. Linea M Zona nuda: porzione centrale senza ponti trasversali. Ogni testa possiede due siti critici: sito di fissazione all actina si lega alle molecole di actina sito ATPasico ha attività enzimatica. Catalizza l idrolisi dell ATP ad ADP e fosforo Titina: è una proteina elastica organizzata in filamenti che si estendono per tutta la lunghezza di un filamento spesso e che mantiene i filamenti spessi nella corretta posizione rispetto a quelli sottili. Se una forza esterna viene esercitata sul muscolo, i filamenti di titina si oppongono allo stiramento come una molla.
 e la zona H si accorciano: i filamenti sottili scivolano lungo i filamenti spessi, muovendosi verso l interno della zona H e riducendone l ampiezza.")
115 La banda A non si accorcia: i filamenti spessi non cambiano lunghezza. Le bande I (filamenti sottili) e la zona H si accorciano: i filamenti sottili scivolano lungo i filamenti spessi, muovendosi verso l interno della zona H e riducendone l ampiezza. Il muscolo si contrae perché i filamenti spessi e sottili delle miofibrille scorrono l uno sull altro: meccanismo dello scorrimento dei filamenti.
116 ad ogni pot d az del motoneurone segue sempre un pot d az nella cellula muscolare il pot d az si propaga per tutto il sarcolemma arrivando ai tubuli a T questo provoca il rilascio di calcio dal reticolo sarcoplasmatico il Ca++ dà il via al ciclo dei ponti trasversali contrazione del muscolo
 il Ca++ dal citoplasma al suo interno (Rs=magazzino per il Ca++) Sulla membrana del RS ci sono: canali voltaggio-dipendenti per il Ca++ che si aprono quando nel tubulo a T arriva un")
117 Quando la cellula muscolare è a riposo, la concentrazione di Ca++ nel citoplasma è molto bassa perché sulla membrana del RS vi sono pompe che trasportano attivamente (contro il gradiente di concentrazione) il Ca++ dal citoplasma al suo interno (Rs=magazzino per il Ca++) Sulla membrana del RS ci sono: canali voltaggio-dipendenti per il Ca++ che si aprono quando nel tubulo a T arriva un pot d az proteine («recettori per la rianodina») che legano funzionalmente i tubuli a T e il RS, permettendo l uscita di Ca++ all arrivo del pot d az nel tubulo a T canali per il Ca++ che si aprono in seguito al legame con altri ioni Ca++: liberazione a valanga di Ca++.


118 Quando la concentrazione di Ca++ è sufficiente, il Ca++ si lega ad una delle tre proteine che formano la troponina, la quale cambia forma. Il cambiamento di forma della troponina induce la tropomiosina a spostarsi, permettendo l esposizione dei siti di fissazione per la miosina presenti sulle molecole di actina. La miosina può fissarsi all actina
119 MECCANISMI MOLECOLARI DELLA CONTRAZIONE MUSCOLARE Ciclo dei ponti trasversali Oscillazione avanti-indietro dei ponti trasversali e ciclica fissazione-distacco dei ponti trasversali ai filamenti sottili: il ponte trasversale spinge i filamenti sottili verso il centro del sarcomero. 1. Quando l ATP si fissa alla miosina, ne cambia la conformazione della testa, provocandone il distacco dall actina ADP + fosforo 2. ADP e fosforo rimangono legati al sito ATPasico 5. Actina e miosina sono strettamente legate ed incapaci di staccarsi se non viene fornita energia 4. La testa della miosina gira verso il centro del sarcomero, tirando il filamento sottile, 3. La testa della miosina si lega ad una molecola di actina
120 THE SLIDING FILAMENT THEORY
121 Un ponte trasversale genera forza solo quando è attivo (colpo di forza). Una cellula muscolare genera forza continuamente durante la contrazione poiché molti ponti trasversali sono attivi contemporaneamente anche se non simultaneamente: alcuni sono all inizio del ciclo, altri a metà, altri alla fine (es. movimenti delle gambe mentre si cammina). Siccome i ponti trasversali ai capi terminali dei filamenti spessi sono orientati in direzione opposta, il colpo di forza attira verso il centro i filamenti sottili da entrambi i lati della banda A, provocando l accorciamento del sarcomero. Alla fine della contrazione i filamenti sottili ritornano nella loro posizione originaria in maniera passiva. Ogni testa di miosina può completare 5 cicli in 1 secondo. In ogni filamento spesso possono avvenire migliaia di colpi di forza in 1 secondo poiché vi sono centinaia di teste.
122 quando il pot d az avvia la liberazione di Ca++ dal RS, questo rilascio non continua all infinito perché appena la concentrazione di Ca++ supera un certo limite il Ca++ si fissa a recettori (con bassissima affinità per il Ca++ e che quindi non vengono attivati finché non vi è tantissimo Ca++) che chiudono i canali voltaggio-dipendenti per il Ca++. Il Ca++ non esce più e viene portato dentro dalla pompa attiva il legame tra calcio e troponina è reversibile e la diminuzione di concentrazione di Ca++ porta il Ca++ a dissociarsi dalla troponina la troponina e la tropomiosina ritornano in posizione di riposo si riduce il numero di siti esposti sull actina si riduce il numero di ponti trasversali attivi quando la concentrazione di Ca++ ritorna ai valori di riposo tutti i ponti trasversali sono inattivi il muscolo è rilassato

123 Accoppiamento eccitazione-contrazione quando una cellula muscolare riceve uno stimolo da un motoneurone, la cellula si depolarizza generando un potenziale d azione che a sua volta scatena la contrazione
124 I muscoli devono essere in grado di rispondere immediatamente e a lungo al segnale nervoso: ATP deve essere disponibile in pochissimo tempo (la fosforilazione a livello di substrato e la fosforilazione ossidativa, normali processi che nelle cellule muscolari e non muscolari forniscono energia, richiedono un tempo più lungo) creatinfosfato = riserva di fosfato. E presente in quantità sufficiente a far fronte ad un fabbisogno quattro o cinque volte superiore alla norma. La reazione del creatinfosfato con l ADP è catalizzata dall enzima creatinchinasi ed è reversibile: Creatinfosfato + ADP (creatinchinasi) creatina + ATP Cellula muscolare a riposo: reazione in equilibrio Cellula muscolare attiva: ATP cala, ADP aumenta, la reazione va verso destra (legge dell azione di massa). Quindi l ADP viene trasformato in ATP che viene utilizzato nel ciclo dei ponti trasversali consumando creatinfosfato. Cellula muscolare si rilascia: aumento di ATP perché non viene consumato, diminuzione di ADP, la reazione va verso sinistra, le scorte di creatinfosfato si ripristinano. La scorta di creatinfosfato è limitata, quindi questa reazione può produrre ATP solo per un tempo breve ma sufficiente affinché si attivino le altre reazioni metaboliche che forniscono ATP.
125 Il metabolismo della cellula muscolare cambia in relazione all intensità dell esercizio: Esercizio costante ma intensità moderata: l ATP è fornito dalla fosforilazione ossidativa che utilizza come carburante per i primi secondi il glucosio fornito dalle scorte di glicogeno, poi nuovo glucosio e acidi grassi forniti dal circolo sanguigno e dopo circa trenta minuti quasi esclusivamente gli acidi grassi. Esercizio intenso: l ATP è fornito principalmente dalla glicolisi anaerobica, in conseguenza alla quale si forma acido lattico, che si accumula nel tessuto muscolare e provoca la sensazione di bruciore e dolore che si avverte nei muscoli dopo un esercizio intenso.
126 Cosa rende un muscolo più forte di un altro? Com è possibile che vengano usati gli stessi gruppi muscolari, come ad esempio quelli del braccio e della mano, per sollevare una seggiola (molta forza) oppure un fermaglio per la carta (poca forza)? per lanciare una palla (contrazione breve ma intensissima) oppure trasportare una valigia (contrazione prolungata)? Per capirlo è necessario partire dalla considerazione che quando una cellula muscolare si contrae in risposta ad un singolo potenziale d azione il risultato è sempre lo stesso: in una frazione di secondo la forza aumenta al massimo e poi rapidamente scende a zero = SCOSSA SINGOLA Se la fibra muscolare viene stimolata di nuovo dà esattamente la stessa risposta
127 La scossa singola isolata si può vedere solo in condizioni artificiali create in laboratorio: Muscolo isolato ancorato ad un apparato che consente di misurare la forza contrattile, stimolato elettricamente. UNITA MOTORIA: motoneurone e tutte le fibre muscolari che innerva. Quando il motoneurone «spara» tutte le fibre muscolari ad esso connesse si contraggono. SCOSSA SINGOLA: risposta meccanica di una sola fibra muscolare, di un unità motoria di un intero muscolo, ad un singolo pot d az
128 periodo latente: ritardo dovuto agli eventi che caratterizzano l accoppiamento eccitazionecontrazione. Tempo tra la comparsa del pot d az nella cellula muscolare e l inizio della contrazione. fase di contrazione: può durare dai 10 ai 100 msec o più a seconda dei muscoli e cessa al raggiungimento del picco massimo di tensione. E caratterizzata dall aumento di concentrazione di calcio nel citosol perché viene liberato dal RS. fase di rilasciamento: tra il picco della forza e la fine della contrazione. Caratterizzata dalla diminuzione della concentrazione di calcio in quanto esso viene ripreso dal RS. Il numero di ponti trasversali attivi va diminuendo. La stimolazione ripetitiva di un muscolo provoca diverse scosse in fila, ognuna delle quali ha la stessa forma e grandezza.
129 Tensione: È sinonimo di forza Viene comunemente espressa in unità di massa, per esempio in grammi; La tensione muscolare di un grammo è equivalente alla forza generata dal muscolo per sostenere un peso di 1 grammo sospeso ad un filo
130 La stimolazione ripetitiva di un muscolo provoca diverse scosse in fila, ognuna delle quali ha la stessa forma e grandezza. E un evento tutto-o-nulla in quanto un potenziale d azione libererà sempre la stessa quantità di calcio dal reticolo sarcoplasmatico, che provocherà sempre lo stesso aumento di concentrazione, che attiverà lo stesso numero di ponti trasversali, producendo sempre la stessa forza.
131 le scosse singole variano molto da un muscolo all altro Alcune fibre muscolari sono più «forti»: diametro maggiore = maggiore forza Le fibre muscolari differiscono nella velocità con cui raggiungono il picco di forza: Fibre rapide Fibre lente
132 Quella che abbiamo visto finora viene detta «Contrazione isometrica» Contrazione isometrica e isotonica: Differiscono per la possibilità o meno che ha il muscolo di potersi accorciare durante la contrazione (il sarcomero si accorcia SEMPRE!). ISOTONICO = uguale tensione (isos=uguale; tonos=tensione) ISOMETRICO= uguale misura (isos=uguale; metrum=misura)

133

134 CONTRAZIONE ISOMETRICA: si genera una tensione senza accorciamento del muscolo perché il carico è superiore alla forza generata dal muscolo. CONTRAZIONE ISOTONICA: si genera una tensione (Tensione = forza espressa in grammi. Tensione di 1 g = forza generata dal peso di 1 g appeso ad un filo) almeno pari alle forze che vi si oppongono (carico: ad esempio il peso di una valigia). Il muscolo si accorcia. La forza aumenta ma il muscolo non si accorcia e quindi la lunghezza rimane uguale: CONTRAZIONE ISOMETRICA Il muscolo si accorcia e il carico si muove. La tensione rimane uguale: CONTRAZIONE ISOTONICA
135 Al contrario della contrazione isometrica, la contrazione isotonica non è un evento tutto-o nulla, in quanto l intensità e la forma della contrazione isotonica dipendono dal carico su cui agisce il muscolo. Anche il periodo tra lo stimolo e l inizio del plateau varia, poiché il muscolo richiede più tempo per sviluppare una forza maggiore (perché è necessario reclutare un numero maggiore di ponti trasversali e quindi bisogna liberare un numero maggiore di siti di legame). Se il carico è superiore alla quantità di forza che il muscolo può sviluppare, la contrazione è isometrica.
136 Quando il muscolo si contrae in modo isometrico, i suoi sarcomeri si accorciano, anche se il muscolo nel suo complesso non si accorcia. I sarcomeri (detti componente contrattile, CC) si collegano alla componente elastica (CE). Quando un muscolo si contrae isometricamente, la CC si accorcia e la CE si stira della medesima lunghezza. In questo modo l effetto complessivo sulla lunghezza è nullo.
137 La forza generata da un muscolo dipende da: 1. La forza sviluppata dalle singole fibre muscolari - Dipende dal numero di ponti trasversali che si legano all actina. Più ponti trasversali si legano maggiore è la forza (come al tiro alla fune!). I fattori che influenzano il numero di ponti trasversali attivi sono: - frequenza di stimolazione - diametro della fibra - variazioni della lunghezza della fibra 2. Il numero di fibre muscolari che si contraggono

138 Frequenza di stimolazione Le contrazioni muscolari isometriche sono di fatto eventi riproducibili, tutto o nulla, solo se il muscolo viene stimolato ad una frequenza abbastanza bassa da assicurare che le scosse singole siano ben separate nel tempo. A frequenza più elevate, il tasso con cui il calcio viene liberato dal RS supera quello con cui il calcio viene attivamente recuperato dal RS aumentandone la concentrazione nel citosol. A concentrazioni maggiori di calcio, un numero maggiore di legami calcio-troponina è presente, di conseguenza aumenta anche il numero di siti leganti la miosina disponibili, determinando un numero maggiore di legami actina-miosina Fenomeno della scala: Scosse singole ravvicinate nel tempo ma indipendenti tra loro. Si ritiene che sia dovuta ad un aumento del calcio citosolico tra una contrazione e l altra.
139 I fenomeni della sommazione e del tetano, che avvengono a frequenze di stimolazione ancora maggiori, sono dovuti alla sovrapposizione delle scosse singole. A paragone con un potenziale d azione una scossa singola è un fenomeno più lento. Per questo in una fibra muscolare si possono sviluppare numerosi potenziali d azione nel tempo impiegato per dare origine ad una contrazione completa. Quando un muscolo viene stimolato ripetitivamente, in modo che il pot d az arrivi prima che la scossa precedente sia giunta a completamento, le scosse si sovrappongono, sviluppando una forza maggiore. Questo fenomeno è chiamato SOMMAZIONE. Quando le scosse singole sono così frequenti che la rimozione del calcio dal citosol non può avvenire in modo altrettanto rapido di quanto esso venga liberato. Questo impedisce il rilasciamento (che si ha SOLO quando il calcio viene rimosso)
140 A frequenze di stimolazione superiori, la sommazione raggiunge un valore massimo chiamato tetano (è lo stesso nome dato ad una malattia in cui nel corso di un infezione batterica la tossina prodotta induce i motoneuroni a stimolare il muscolo in modo improprio). Tetano non fuso o incompleto: Piccole oscillazioni della forza: i picchi si hanno quando la quantità di calcio è così alta che la troponina risulta saturata e quindi tutti i siti di legame sono esposti. Tetano completo: La concentrazione di calcio è così alta che la troponina è saturata in modo permanente. Aumentando ancora di più la frequenza di stimolazione la forza non aumenta in quanto non ci sono altri siti di legame da liberare.

 dal numero di sarcomeri (e quindi di filamenti spessi e sottili) più sarcomeri in parallelo: più forza Siccome il numero di filamenti spessi e sottili per unità di sezione trasversale non varia")
141 Diametro delle fibre: forza contrattile: la capacità di generare forza da parte di un muscolo. Viene valutata misurando la forza tetanica massimale o il picco di tensione durante una contrazione isometrica. La capacità di produrre forza da parte di una fibra dipende dal numero di ponti trasversali in ciascun sarcomero più ponti trasversali sono presenti, più possono essere attivi: più forza (tiro alla fune) dal numero di sarcomeri (e quindi di filamenti spessi e sottili) più sarcomeri in parallelo: più forza Siccome il numero di filamenti spessi e sottili per unità di sezione trasversale non varia significativamente da un muscolo ad un altro, il diametro della fibra è una variabile fondamentale nel determinare la forza contrattile. maggiore è il DIAMETRO maggiore è la forza generata (muscoli ipertrofici sollevatore di pesi)
142 Variazioni della lunghezza della fibra: A seconda della lunghezza a riposo del muscolo, questo può generare maggiore o minore forza perché alle variazioni di lunghezza del muscolo corrispondono variazioni di lunghezza dei singoli sarcomeri I ponti trasversali sono tutti attivi, ma i filamenti sottili iniziano ad accavallarsi gli uni con gli altri e questo interferisce con lo sviluppo della forza. Le linee Z toccano i filamenti spessi e la maggior parte della forza generata dai ponti trasversali viene esercitata sul sarcomero stesso invece di essere trasmessa alle estremità della fibra muscolare Normale intervallo di lunghezza quando i muscoli si trovano nel corpo: il movimento è limitato dalle ossa cui sono attaccati. Generalmente operano nell intervallo di lunghezze entro le quali sono in grado di generare la massima forza. La tensione generata diminuisce linearmente all aumentare della sua lunghezza. Il grado di sovrapposizione dei filamenti spessi e sottili diminuisce all allungarsi dei sarcomeri. I ponti trasversali di miosina che non hanno un corrispettivo sito di attacco non possono fissarsi all actina (NON SONO ATTIVI) e quindi non possono generare forza.
143 La forza generata da un muscolo dipende da: 1. La forza sviluppata dalle singole fibre muscolari - Dipende dal numero di ponti trasversali che si legano all actina. Più ponti trasversali si legano maggiore è la forza (come al tiro alla fune!). I fattori che influenzano il numero di ponti trasversali attivi sono: - frequenza di stimolazione - diametro della fibra - variazioni della lunghezza della fibra 2. Il numero di fibre muscolari che si contraggono La massima forza che una fibra muscolare può sviluppare corrisponde alla massima tensione tetanica (massima frequenza di stimolazione capace di aumentare la tensione) che è circa 5 volte superiore alla forza sviluppata da una scossa singola. Dato che la forza deve variare moltissimo per utilizzare gli stessi muscoli per compiti diversi (sollevare una graffetta oppure una valigia) è chiaro che la variazione di frequenza dei pot d az può coprire solo una piccola parte dell ampio ambito di variazioni di forza.

144 Modulazione della forza generata dal muscolo in toto: Quando un muscolo si contrae solo di rado tutte le sue fibre generano forza (=ricevono potenziali d azione). Alcune unità motorie sono attive, altre si accorciano passivamente. Quando sono richieste forze maggiori, il sistema nervoso aumenta il numero di fibre attive. Reclutamento: aumento del numero di unità motorie attive. Le unità motorie differiscono per il numero di fibre muscolari che le compongono.
145 Principio della dimensione: se il muscolo deve generare forze piccole vengono reclutate unità motorie con meno fibre muscolari (più piccole) se il muscolo deve generare forze maggiori vengono reclutate fibre muscolari più grandi. Il controllo fine è più facile quando la forza richiesta è piccola: è possibile avere piccole variazioni di forza reclutando piccole unità motorie addizionali (aumenta di poco il numero totale di fibre attive). Non si può avere controllo fine quando sono richiesti grandi sforzi perché è possibile reclutare solo unità motorie grandi, che determinano ampi incrementi di forza. Unità motorie grandi sono comandate da motoneuroni grandi che raggiungono più difficilmente la soglia per il pot d az Unità motorie piccole sono comandate da motoneuroni piccoli che raggiungono più facilmente la soglia per il pot d az Quando un input sinaptico aumenta gradualmente i motoneuroni piccoli scaricano prima e quelli grandi dopo.
146 La forza generata dal muscolo in toto dipende da una combinazione di fattori che agiscono sia sulle singole fibre che dal numero di fibre attive. Nel determinare il movimento muscolare, però intervengono anche altri fattori quali la velocità di contrazione. Per determinare la velocità di accorciamento di un muscolo, occorre stimolarlo a contrarsi in modo isotonico ed esprimere in un grafico la variazione di lunghezza in funzione del tempo. Muscolo stimolato con scossa semplice in maniera isotonica. All aumentare del carico: aumenta il periodo di latenza diminuisce la durata dell accorciamento diminuisce la velocità dell accorciamento All aumentare del carico la velocità diminuisce progressivamente fino ad annullarsi quando il carico è uguale alla tensione massima. La velocità massima si ha quando nessun carico è applicato.
.")

147 Alcuni tipi di fibre muscolari si possono accorciare più rapidamente di altri (impiegano meno tempo di altre a raggiungere il picco di tensione). Fibre a contrazione rapida: si contraggono rapidamente composti da miosina rapida: idrolizza ATP velocemente e quindi può completare in 1 sec un numero maggiore di cicli dei ponti trasversali Fibre a contrazione lenta: si contraggono lentamente composti da miosina lenta: idrolizza ATP più lentamente. Numero minore di cicli di ponti trasversali.
148 Tutte le fibre muscolari possono produrre ATP mediante fosforilazione ossidativa (mitocondri) e mediante fosforilazione a livello del substrato (via glicolitica), ma alcune preferiscono un tipo e altre l altro: fibre ossidative ricche di mitocondri (sede della fosforilazione ossidativa), bassa concentrazione di enzimi glicolitici produzione di ATP mediante fosforilazione ossidativa Non producono acido lattico se vengono rifornite adeguatamente di ossigeno Sono resistenti alla fatica piccolo calibro ben vascolarizzate hanno la possibilità di utilizzare l ossigeno per produrre energia e quindi devono poter accedere facilmente all ossigeno ematico (molti capillari). Il piccolo diametro permette di ridurre la distanza che deve percorrere l ossigeno per raggiungere i mitocondri. Contengono mioglobina: proteina di colore rossastro che fissa reversibilmente l ossigeno. Funziona da riserva intracellulare di ossigeno. Lo libera quando la concentrazione intracellulare di ossigeno diminuisce. E una scorta limitata che può essere utilizzata solo per breve tempo. Il colore rosso della mioglobina rende rosse queste fibre che vengono dette fibre rosse. fibre glicolitiche elevata concentrazione di enzimi glicolitici, pochi mitocondri rapida produzione di ATP attraverso la fosforilazione a livello del substrato (via glicolitica) ma meno efficace (ne producono meno in proporzione) Non necessitano di ossigeno Viene generato acido lattico (responsabile della fatica muscolare) Si affaticano più rapidamente grande calibro circondate da pochi capillari. Non contengono mioglobina: fibre bianche
149 Le fibre muscolari scheletriche possono essere classificate come: fibre rapide e fibre lente sulla base della loro velocità di contrazione fibre glicolitiche e fibre ossidative sulla base delle loro capacità metaboliche Possiamo trovare delle combinazioni di queste caratteristiche: fibre ossidative lente, fibre ossidative rapide, fibre glicolitiche rapide Generalmente i muscoli contengono tutti e tre i tipi di fibre, ma in diverse proporzioni. Le diverse unità motorie, però, contengono solo fibre dello stesso tipo.
150 Fibre ossidative lente: contengono miosina lenta alta capacità ossidativa diametro piccolo: generano piccole forze Sono le prime ad essere reclutate Fibre ossidative rapide: contengono miosina con attività ATPasica intermedia tra quella lenta e quella veloce alta capacità ossidativa diametro intermedio: forze intermedie Sono le seconde ad essere reclutate Fibre glicolitiche rapide: contengono miosina rapida alta capacità glicolitica diametro grande: forze grandi Sono le ultime ad essere reclutate. Vengono reclutate solo nel caso di necessità di forze molto intense come il sollevamento pesi o lo sprint finale in una gara di mezzofondo o di maratona.

151 Fatica: progressiva diminuzione della capacità di conservare una forza di contrazione costante nel tempo. Compare più rapidamente quando un muscolo viene stimolato ad alta frequenza e la forza che viene generata è maggiore. Le cause precise della fatica non sono note ma a seconda del tipo di esercizio si sviluppa maggiore o minore fatica: esercizio intenso reclutate le fibre glicolitiche produzione di acido lattico La fatica insorge prima esercizio poco intenso reclutate fibre ossidative no produzione di acido lattico La fatica insorge più tardi (diminuzione riserve energetiche?) Fatica neuromuscolare: la generazione di forze molto intense è determinata da una stimolazione da parte dei motoneuroni ad alta frequenza: si abbassa il livello di Ach nelle terminazioni sinaptiche e quindi vi è minor efficacia della trasmissione neuromuscolare.
152 A seconda dello sport che un atleta vuole praticare è necessario utilizzare tipi diversi di esercizio. Esercizi diversi permettono di adattare l organismo ad esigenze diverse. Grazie all esercizio regolare protratto per un tempo prolungato è possibile modificare l architettura cellulare del muscolo. Maratoneta, grande resistenza: esercizio aerobico (es. corsa regolare a ritmo lento) di lunga durata e bassa intensità aumenta la capacità ossidativa delle fibre muscolari: alcune fibre glicolitiche vengono convertite in fibre ossidative (le fibre lente rimangono lente e quelle rapide rimangono rapide, perché non viene modificato il tipo di miosina presente): aumento forma e dimensione dei mitocondri e aumento del numero di capillari che circondano le fibre. Il diametro delle fibre diminuisce (l ossigeno raggiunge più rapidamente i mitocondri) e quindi diminuisce la capacità della cellula di generare forza. Pugile, muscoli grossi e forti: esercizio intenso aumenta la capacità glicolitica delle fibre muscolari: alcune fibre ossidative vengono convertite in fibre glicolitiche: diminuzione delle dimensioni e del numero di mitocondri, aumento concentrazione enzimi glicolitici, aumento del diametro medio della fibra determinato dalla sintesi di nuove miofibrille che permettono al muscolo di generare più forza. Il muscolo risulta più voluminoso e compatto.

153
154 RESPIRAZIONE

155
156 CAPITOLO 16 L apparato respiratorio: la ventilazione polmonare
 Emissione di aria dagli alveoli")
157 Respirazione esterna: scambio dell ossigeno e dell anidride carbonica tra l atmosfera e i tessuti del corpo La funzione della respirazione esterna è di - Portare l ossigeno ai tessuti - Rimuovere l anidride carbonica Può essere divisa in quattro eventi principali VENTILAZIONE POLMONARE Immissione di aria dall atmosfera negli alveoli polmonari (inspirazione) Emissione di aria dagli alveoli polmonari all atmosfera (espirazione) DIFFUSIONE DELL O 2 E DELLA CO 2 FRA ALVEOLI E SANGUE TRASPORTO DELL O 2 E DELLA CO 2 DA E VERSO LE CELLULE grazie al sangue DIFFUSIONE DELL O 2 E DELLA CO 2 FRA SANGUE E TESSUTI Respirazione interna (respirazione cellulare): utilizzo dell ossigeno all interno dei mitocondri per generare ATP grazie alla fosforilazione ossidativa e produzione di anidride carbonica come prodotto di scarto.

158 Gli organi principali del sistema respiratorio sono i polmoni, che si trovano nella cavità toracica. Ciascun polmone è diviso in lobi (destro, tre lobi; sinistro, due lobi). L aria entra ed esce dai polmoni attraverso le vie respiratorie superiori e attraverso una rete di condotti chiamata tratto respiratorio. vie respiratorie superiori: passaggi per l aria che si trovano nella testa e nel collo. L aria entra nella cavità nasale e/o nella cavità orale che conducono alla faringe, e da essa l aria entra nella prima struttura del tratto respiratorio, la laringe. tratto respiratorio: tutte le vie di passaggio dalla laringe ai polmoni (incluse quelle presenti all interno degli stessi polmoni)

159 Zona di conduzione trachea, un condotto di circa 2,5 cm di diametro e 10 cm di lunghezza che decorre parallelamente e anteriormente all esofago. la trachea si divide nei bronchi di destra e di sinistra che portano l aria in ciascun polmone. i bronchi si dividono in piccoli condotti chiamati bronchi secondari Ciascun bronco secondario si divide in bronchi terziari più piccoli che, a loro volta, si ramificano in bronchi ancora più piccoli. L intera ramificazione risulta essere formata da circa 8 milioni di tubuli, i più piccoli dei quali hanno un diametro inferiore a 0,5 mm. Quando i tubuli presentano un diametro inferiore al millimetro, vengono chiamati bronchioli ( piccoli bronchi ). I bronchioli si ramificano ulteriormente a formare i bronchioli terminali Zona respiratoria bronchioli respiratori, terminano nei dotti alveolari, che portano agli alveoli, la sede nella quale si verifica lo scambio dei gas. Molti alveoli sono raggruppati in strutture chiamate sacchi alveolari, che assomigliano a grappoli di uva; alcuni alveoli, invece, si aprono sui bronchioli respiratori.


.")
160 Oltre alla sottigliezza della membrana respiratoria, anche l abbondanza del numero degli alveoli e dei capillari facilita la diffusione dei gas. I 300 milioni di alveoli nei polmoni formano una superficie totale di circa 100 metri quadri (circa un campo da tennis). Gli alveoli sono ricoperti da una fitta rete di capillari (detta «lenzuolo di sangue»).
, lo sterno, le vertebre toraciche, i muscoli e tessuti connettivi")
161 I polmoni si trovano nella cavità toracica. La parete toracica è composta da strutture che proteggono i polmoni: la gabbia toracica (formata da 12 paia di coste o costole), lo sterno, le vertebre toraciche, i muscoli e tessuti connettivi associati. I muscoli della parete toracica responsabili della respirazione sono i muscoli intercostali interni ed esterni che si inseriscono sulle costole, e il diaframma a forma di cupola che delimita inferiormente la parete toracica e separa le cavità toracica e addominale. La superficie interna della parete toracica e la superficie esterna dei polmoni sono coperte da una membrana chiamata pleura; ciascun polmone è inoltre ricoperto da un proprio distinto sacco pleurico. La porzione di sacco pleurico attaccato al tessuto polmonare viene chiamato pleura viscerale; la porzione attaccata alla parete toracica viene chiamata pleura parietale. Tra le due pleure si trova un compartimento molto sottile chiamato spazio intrapleurico che è riempito da una piccola quantità (circa 15 millilitri) di liquido intrapleurico.
162 Zona di conduzione (laringe, trachea, bronchi e bronchioli) permette il passaggio dell aria dalla faringe ai polmoni. Non si verificano scambi di gas. Contiene approssimativamente 150 ml di aria. Viene detta «spazio morto anatomico». L aria viene umidificata e la temperatura si avvicina a quella corporea.. La zona di conduzione è ricoperta da un epitelio che contiene le cellule a calice e le cellule ciliate. Zona respiratoria (bronchioli respiratori, dotti alveolari, alveoli e sacchi alveolari) è la sede degli scambi di gas nei polmoni; gli alveoli costituiscono la sede principale di scambio. Esaltazione della superficie e spessore minimo, così da facilitare la diffusione dei gas. La differenza anatomica principale tra la zona di conduzione e quella respiratoria consiste nello spessore delle pareti che costituiscono gli spazi aerei: solo gli spazi aerei delimitati da pareti sufficientemente sottili possono partecipare agli scambi di gas.
163 La parete di un alveolo contiene cellule di tipo I e di tipo II. Le cellule di tipo I e le cellule endoteliali dei capillari formano la membrana respiratoria attraverso cui ha luogo lo scambio gassoso. Le cellule di tipo II secernono la sostanza tensioattiva polmonare. Negli alveoli si trovano anche i macrofagi alveolari.

164 La parete interna degli alveoli è ricoperta da un sottile strato di acqua. Durante l inspirazione gli alveoli si espandono, di conseguenza anche lo strato di acqua si deve espandere. La tensione superficiale di un liquido è una misura del lavoro necessario ad aumentare di una certa quantità la sua superficie. Maggiore è la tensione superficiale, maggiore è il lavoro necessario per stendere il liquido. La presenza di una sostanza simile a un detergente chiamata sostanza tensioattiva polmonare o fattore surfattante diminuisce la tensione superficiale degli alveoli in quanto interferisce con i legami idrogeno tra le molecole d acqua. La sostanza tensioattiva polmonare: - Diminuisce il lavoro necessario a espandere gli alveoli - Diminuisce la tendenza dell alveolo a collassare a causa della presenza del liquido che ricopre la sua parete interna.
165 ANATOMY REVIEW
166 muscoli inspiratori primari: diaframma e muscoli intercostali esterni muscoli espiratori primari: muscoli intercostali interni e quelli addominali (sebbene l espirazione sia principalmente un processo passivo che non richiede alcuna contrazione muscolare) Respirazione tranquilla: -inspirazione: il diaframma si contrae determinando una trazione verso il basso della superficie inferiore dei polmoni le costole si innalzano -espirazione: il diaframma si rilascia ed i polmoni si retraggono in virtù del RITORNO ELASTICO dei polmoni, delle pareti toraciche e delle strutture addominali Respirazione forzata: -espirazione (le forze elastiche non sono sufficienti): contrazione dei muscoli addominali che preme i visceri addominali verso l alto, contro la superficie inferiore del diaframma

167

168 Legge di Boyle Data una quantità finita di gas (quale l aria) all interno di un contenitore, la pressione è inversamente proporzionale al volume del contenitore. Le molecole di gas sono in uno stato di costante agitazione termica. Esse continuano a sbattere contro le pareti del contenitore e a rimbalzare. Ciascuna collisione esercita una certa forza sulle pareti. La pressione è la forza totale esercitata dal numero totale di collisioni diviso la superficie delle pareti (forza per unità di superficie) Se il volume del contenitore aumenta, la pressione esercitata dal gas (numero di collisioni con le pareti) diminuisce, mentre se il volume diminuisce, la pressione aumenta.
169 Sono quattro le pressioni principali associate alla ventilazione. Polmone a riposo: tra due respiri, alla fine di un espirazione tranquilla la pressione atmosferica (P atm ) pressione dell aria esterna. A livello del mare la pressione atmosferica è normalmente 760 mm Hg. Tutte le altre pressioni polmonari sono espresse relativamente alla pressione atmosferica. la pressione alveolare (P alv ) pressione dell aria all interno degli alveoli. A riposo è pari alla pressione atmosferica, quindi è 0 mm Hg. la pressione intrapleurica (P ip ) pressione all interno del sacco pleurico. A riposo è -4 mm Hg La pressione transpolmonare corrisponde alla differenza tra la pressione alveolare e quella intrapleurica (P alv P ip ) E la forza applicata ai polmoni per espanderli
170 PULMONARY VENTILATION Legge di Boyle
171 La ventilazione (movimento di aria dentro e fuori dai polmoni) ha luogo grazie alla presenza di gradienti di pressione tra gli alveoli e l aria esterna (atmosferica) e l aria si muove a causa del gradiente di pressione dalle zone di alta pressione a quelle di bassa pressione. l inspirazione si verifica quando la pressione atmosferica è maggiore di quella alveolare aria all interno degli alveoli l espirazione si ha quando la pressione negli alveoli è maggiore di quella nell atmosfera fuoriuscita di aria dagli alveoli
172 il flusso d aria dentro e fuori dai polmoni viene definito come flusso di volume, la cui velocità viene determinata da un gradiente di pressione (P atm P alv ) e da una resistenza, in base alla formula: Flusso = (P atm P alv ) / R Quindi, la forza che guida il movimento dell aria dipende dalla differenza tra la pressione atmosferica e quella alveolare. Siccome la pressione atmosferica è costante, sono le modificazioni della pressione alveolare a determinare la direzione del movimento d aria. Capacità funzionale residua (CFR) volume di aria che si trova nei polmoni quando essi sono a riposo e tutti i muscoli respiratori sono rilassati.

173 La pressione alveolare viene determinata da due fattori: la quantità (in termini molecolari) di aria negli alveoli e il volume stesso degli alveoli. A riposo, gli alveoli contengono un volume di aria che, ad una pressione pari a quella atmosferica, è uguale alla capacità funzionale residua. All inizio dell inspirazione, i polmoni si espandono in seguito alla contrazione dei muscoli inspiratori. Tale contrazione determina un aumento del volume degli alveoli e, quindi, una diminuzione della pressione alveolare, in accordo con la legge di Boyle. La riduzione della pressione alveolare determina un gradiente di pressione che attira aria nei polmoni. Durante l espirazione avviene il contrario: il torace e i polmoni tendono a ritornare nella condizione di riposo, facendo diminuire il volume degli alveoli e facendo aumentare la pressione alveolare. Questo determina un gradiente di pressione che fa fuoriuscire l aria dai polmoni. Un espirazione forzata può essere determinata dalla contrazione dei muscoli espiratori in un processo chiamato espirazione attiva. La contrazione dei muscoli espiratori produce una maggiore e più rapida diminuzione di volume della cavità toracica, che determina un maggiore aumento della pressione alveolare e quindi del gradiente di pressione che spinge l aria fuori dagli alveoli.
174 ESPIRAZIONE Il rilassamento dei muscoli inspiratori fa sì che la parete toracica e i polmoni, che sono strutture elastiche, ritornino alla loro posizione di riposo. Il rilassamento dei muscoli inspiratori si verifica nel momento in cui i motoneuroni dei muscoli inspiratori smettono di essere attivi. Mentre la parete toracica e i polmoni ritornano alla posizione iniziale, il volume dei polmoni diminuisce, determinando un valore di pressione alveolare maggiore di quella atmosferica.


175 La pressione intrapleurica varia con le fasi della ventilazione Ricordiamo che lo spazio intrapleurico contiene liquido intrapleurico e non aria. La pressione intrapleurica è sempre inferiore a quella alveolare e risulta essere sempre negativa durante la respirazione normale in quanto forze opposte esercitate dalla parete toracica (che tende ad espandersi come succede ad una molla che è stata compressa), e dai polmoni (che tendono a contrarsi come un palloncino che è stato gonfiato) tendono a separare la pleura parietale da quella viscerale MA la tensione superficiale del liquido intrapleurico impedisce la separazione (es.: vetrini da microscopio bagnati) determinando una pressione intrapleurica negativa (liquido=incomprimibile, inespandibile)
, la pressione intrapleurica non è più negativa in quanto essa si pone in equilibrio con quella atmosferica.")
176 pneumotorace (aria nello spazio intrapleurico) Per poter mantenere una pressione intrapleurica negativa è necessario che il sacco intrapleurico sia a tenuta d aria. Se il sacco pleurico viene rotto (coltellata o colpo d arma da fuoco al torace), la pressione intrapleurica non è più negativa in quanto essa si pone in equilibrio con quella atmosferica. In assenza di pressione intrapleurica negativa, i polmoni collassano e la parete toracica si espande. Fortunatamente ciascun polmone è isolato dall altro nella propria cavità pleurica e quindi se un polmone collassa, l altro può continuare la sua funzione. Il pneumotorace spontaneo viene causato quando uno stato patologico danneggia la parete della pleura adiacente a un bronco o a un alveolo permettendo all aria che si trova nei polmoni di entrare nello spazio intrapleurico. Le patologie che comunemente possono causare un pneumotorace spontaneo sono la polmonite e l enfisema.

177 Pressione transpolmonare corrisponde alla differenza tra la pressione alveolare e quella intrapleurica (P alv P ip ) L espandersi della parete toracica attrae verso l esterno il liquido intrapleurico, causando una diminuzione della pressione intrapleurica. Questa diminuzione della pressione intrapleurica induce un aumento della pressione transpolmonare determinando una maggiore azione distensiva sui polmoni e quindi gli alveoli si espandono assieme alla parete toracica.
178 Flusso d aria = (P atm P alv ) / R Fattori che influenzano l instaurarsi dei gradienti di pressione e la resistenza delle vie respiratorie: Compliance (distensibilità) polmonare Misura della facilità con la quale i polmoni possono essere stirati. Viene definita come il cambiamento di volume polmonare ( V) determinato da una certa modificazione nella pressione transpolmonare (P alv P ip ): Compliance polmonare = V/(P alv P ip ) V = Compliance polmonare x (P alv P ip ) Una maggior compliance polmonare è vantaggiosa in quanto è necessario un minor cambiamento della pressione transpolmonare per spostare un dato volume d aria e, di conseguenza, viene richiesto un minor lavoro o una minore contrazione muscolare. La compliance polmonare dipende dall elasticità dei polmoni I polmoni sono elastici grazie alla presenza di fibre elastiche nel tessuto connettivo: le forze esercitate da queste fibre si oppongono all espansione polmonare in quanto, nel momento in cui il polmone viene stirato, esse tendono a ritornare nella loro posizione iniziale. e dalla tensione superficiale del fluido che ricopre gli alveoli La tensione superficiale di un liquido è una misura del lavoro necessario ad aumentare di una certa quantità la sua superficie. Maggiore è la tensione superficiale, maggiore lavoro è necessario per stendere ulteriormente il liquido. La tensione superficiale nei polmoni è determinata dall interfaccia aria-liquido, formata dal sottile strato di liquido che ricopre la superficie interna degli alveoli. All espandersi del tessuto polmonare, si estende anche lo strato di liquido che riveste gli alveoli. Quindi, nel momento in cui i polmoni si espandono, viene richiesto del lavoro non solo per stirare il tessuto elastico ma anche per aumentare la superficie dello strato liquido che riveste gli alveoli. Di conseguenza, la tensione superficiale è un fattore che si oppone all espansione, diminuendo la compliance polmonare.
179 Aumento della compliance polmonare: sostanza tensioattiva polmonare: sostanza simile ad un detergente che diminuisce la tensione superficiale negli alveoli. Viene secreta dalle cellule alveolari di tipo II poste nelle pareti degli alveoli. Essa interferisce con il legame idrogeno tra le molecole d acqua. Di conseguenza, la sostanza tensioattiva aumenta la compliance polmonare e diminuisce il lavoro respiratorio. Riduzione della compliance polmonare: se il tessuto polmonare si ispessisce, come avviene in presenza di tessuto cicatriziale nella tubercolosi o se la produzione di sostanza tensioattiva diminuisce, come avviene nella sindrome da insufficienza respiratoria del neonato (nella quale i bambini prematuri non producono sufficiente sostanza tensioattiva). Quando diminuisce la compliance polmonare, i muscoli respiratori devono compiere un lavoro maggiore per espandere i polmoni.
 e inversamente proporzionale al raggio (r) P = 2T/r Maggior concentrazione di tensioattivo negli alveoli piccoli =minor tensione superficiale =minor pressione")
180 Il tensioattivo polmonare (surfattante) e la legge di Laplace La pressione d aria (P) necessaria a prevenire il collasso di un alveolo è direttamente proporzionale alla tensione superficiale (T) e inversamente proporzionale al raggio (r) P = 2T/r Maggior concentrazione di tensioattivo negli alveoli piccoli =minor tensione superficiale =minor pressione necessaria
181 PULMONARY VENTILATION Fino a resistenza
182 La velocità con la quale l aria entra ed esce dai polmoni è determinata da: Il gradiente di pressione tra l aria atmosferica e gli alveoli La resistenza delle vie respiratorie
183 Flusso d aria = (P atm P alv ) / R Resistenza delle vie respiratorie: quando la resistenza aumenta è necessario un gradiente di pressione maggiore per determinare un dato flusso d aria. È influenzata in gran parte dalle dimensioni del loro raggio: al diminuire del raggio aumenta la resistenza. Nei polmoni sani la resistenza al flusso è bassa in quanto il raggio dei condotti presenti nella zona di conduzione è relativamente grande. Nonostante il raggio diminuisca procedendo verso gli alveoli, l area della sezione totale dei tubuli più piccoli aumenta perché la ramificazione è maggiore. Data la bassa resistenza, in condizioni di normalità la pressione alveolare non deve essere molto diversa da quella atmosferica per ottenere una normale velocità di flusso d aria < 2 mm Hg Quando la resistenza aumenta è necessario un gradiente di pressione maggiore per determinare un dato flusso d aria
184 attività contrattile dei muscoli lisci: responsabili delle variazioni a lungo termine muscolo liscio che si trova nelle pareti dei bronchioli: Quando si contrae, il raggio dei bronchioli si riduce (questo fenomeno è chiamato broncocostrizione), aumentando la resistenza. La contrazione e il rilassamento del muscolo liscio bronchiolare è soggetto sia ad un controllo estrinseco (segnali neuronali e ormonali) che ad un controllo intrinseco (mediatori chimici locali). influenzato dal sistema nervoso autonomo: la stimolazione simpatica determina il rilassamento del muscolo liscio e aumenta il raggio dei bronchioli (fenomeno chiamato broncodilatazione), mentre la stimolazione parasimpatica determina la contrazione del muscolo liscio e la broncocostrizione. Anche l adrenalina rilasciata dalla ghiandola surrenale durante la stimolazione simpatica determina broncodilatazione. L istamina, una sostanza chimica rilasciata localmente durante le reazioni allergiche, causa la contrazione del muscolo liscio determinando broncocostrizione. L istamina stimola anche la secrezione di muco, che si accumula nelle vie respiratorie e aumenta la resistenza al flusso. alti livelli di anidride carbonica: i bronchioli si dilatano bassi livelli di anidride carbonica:i bronchioli si restringono
185 PULMONARY VENTILATION Dopo resistenza
186 Asma aumento acuto della resistenza delle vie respiratorie determinata dalla contrazione spastica del muscolo liscio che si trova nei bronchioli accoppiata ad un aumento della secrezione di muco e ad un infiammazione delle pareti dei bronchioli. I sintomi comprendono la tosse, la dispnea (respirazione difficoltosa) e la respirazione ansimante. L asma dipende spesso da un ipersensibilità ad alcuni allergeni (sostanze che stimolano le risposte allergiche), quali i funghi, le particelle di polvere, il pelo degli animali, ma può anche essere causata dallo stress, dall attività fisica, da alcuni cibi o dal respirare in condizioni di bassa temperatura ambiente. Siccome la resistenza delle vie respiratorie diviene maggiore, sono necessari gradienti di pressione maggiori per determinare un uguale flusso d aria, aumentando notevolmente il lavoro respiratorio. La terapia dell asma varia a seconda degli individui ma può includere broncodilatatori che inducono un rilassamento dei muscoli lisci delle vie respiratorie e corticosteroidi, che riducono l infiammazione. malattie ostruttive polmonari croniche aumento cronico della resistenza delle vie respiratorie
187 spirometro apparecchio che permette di misurare i volumi di aria inspirata ed espirata è formato da una campana rovesciata e riempita di aria posta in un contenitore pieno di acqua e collegata tramite un tubo flessibile al paziente. Il paziente respira l aria contenuta nella campana. Quando il paziente inspira, il volume d aria nella campana diminuisce e la campana scende più profondamente nell acqua. Quando il paziente espira, il volume d aria nella campana aumenta e la campana sale verso l alto. La campana è connessa ad una penna che si muove verso l alto e verso il basso seguendo il movimento della campana tramite un sistema a puleggia. La penna è posizionata in modo tale da scrivere sulla carta che ricopre un cilindro rotante che si muove ad una velocità nota e costante. I movimenti della penna lasciano un tratto sulla carta che indica il volume di aria che entra ed esce dai polmoni.
188
189 volume corrente (VC) Il volume di aria che entra ed esce dai polmoni durante un singolo atto respiratorio non forzato. A riposo è di 500 ml. volume di riserva inspiratoria (VRI) Il volume di aria che può ancora essere inspirato al termine di una normale inspirazione ed è in media circa 3000 ml. volume di riserva espiratoria (VRE) Il volume di aria che può essere ancora espirato al termine di una normale espirazione ed è in media circa 1000 ml. volume residuo (VR) Il volume di aria che rimane nei polmoni dopo una massima espirazione e corrisponde a circa 1200 ml. Anche in seguito ad una massima espirazione, rimane dell aria nei polmoni e nelle vie respiratorie in quanto l adesione dei foglietti pleurici in seguito all esistenza di una pressione intrapleurica negativa impedisce il completo collasso dei polmoni.
190 capacità polmonare totale (CPT=VC+VRI+VRE+VR) E il massimo volume a cui i polmoni possono essere espansi con un inspirazione forzata massima (circa 5700 ml) capacità inspiratoria (CI=VC+VRE) è il volume massimo di aria che può essere inspirata alla fine di un espirazione tranquilla e corrisponde a circa 3500 ml. capacità vitale (CV=VC+VRI+VRE) è il volume massimo di aria che può essere espirata successivamente ad un inspirazione massimale e corrisponde a circa 4500 ml. capacità funzionale residua (CFR=VRE+VR) è il volume di aria che rimane nei polmoni al termine di un espirazione tranquilla e corrisponde approssimativamente a 2200 ml. La capacità funzionale residua è il volume d aria presente nei polmoni quando la persona si trova tra due atti respiratori e i muscoli respiratori sono rilassati. In queste condizioni la tendenza elastica dei polmoni a ritornare nella posizione di partenza è bilanciata da quella della parete toracica.
191 malattie polmonari ostruttive determinano un aumento nella resistenza delle vie respiratorie il volume residuo spesso aumenta in quanto un aumento nella resistenza non solo rende difficoltosa l inspirazione ma anche l espirazione. I polmoni sono troppo pieni di aria e sia la capacità funzionale residua che la capacità polmonare totale spesso risultano aumentate malattie polmonari restrittive qualcosa interferisce con l espansione dei polmoni un danno strutturale ai polmoni, alla pleura o alla parete toracica che diminuisce la capacità polmonare totale e la capacità vitale. test della capacità vitale forzata (CVF) il paziente esegue una massima inspirazione e quindi espira forzatamente il più rapidamente ed efficacemente possibile. Volume d'aria espirata forzatamente dopo un'inspirazione massimale. Una CVF ridotta è indice di un disordine polmonare restrittivo. test del volume espiratorio forzato (FEV) la quantità di aria espirata durante il primo secondo di un'espirazione forzata dopo un inspirazione massimale. E la misura della percentuale di CVF che può essere espirata in un certo periodo di tempo, di norma un secondo (FEV 1 ). Un FEV 1 normale corrisponde all 80% della CVF, il che significa che individuo normale deve essere in grado di espirare l 80% della capacità vitale forzata in un secondo. Una FEV 1 inferiore a 80% indica un aumento di resistenza, che è una caratteristica delle affezioni ostruttive polmonari.
192 ventilazione al minuto quantità totale di aria che entra ed esce dal sistema respiratorio in un minuto: = volume corrente X frequenza respiratoria (numero di atti respiratori al minuto, 12) = VC x frequenza respiratoria = (500 ml/atto respiratorio) x (12 atti respiratori/minuto) = 6000 ml/minuto Più importante della ventilazione al minuto è però la quantità di aria fresca che raggiunge gli alveoli. Solo una porzione dell aria che viene respirata partecipa realmente allo scambio gassoso in quanto una significativa porzione di aria semplicemente riempie le vie aeree della zona di conduzione. Ad esempio, considerando una normale inspirazione di 500 ml di aria, solamente 350 ml raggiungono in realtà gli alveoli; i restanti 150 ml (30% di un volume corrente pari a 500 ml, in questo esempio) riempiono la trachea, i bronchi e i bronchioli. Il volume combinato di queste vie non deputate allo scambio gassoso viene chiamato spazio morto anatomico.
193 Al termine dell espirazione, la zona di conduzione è piena di aria vecchia (aria composta da una quantità maggiore di anidride carbonica e minore di ossigeno rispetto all aria atmosferica), essendo l aria che occupava gli alveoli durante l inspirazione e dove aveva scambiato con il sangue ossigeno e anidride carbonica. Nella inspirazione successiva, questi 150 ml di aria vecchia proseguono verso gli alveoli assieme a 350 ml di aria fresca (assumendo un volume corrente di 500 ml). Quindi, da un punto di vista funzionale, solo 350 ml di aria fresca entrano negli alveoli ad ogni atto respiratorio. La ventilazione alveolare è una misura del volume di aria fresca che raggiunge gli alveoli ogni minuto. E simile alla ventilazione al minuto ad eccezione del fatto che il volume corrente è corretto rispetto al volume dello spazio morto (SM): Ventilazione alveolare = (VC x frequenza respiratoria) (SM x frequenza respiratoria) = (500 ml/atto respiratorio x 12 atti respiratori/minuto) (150 ml/atto respiratorio x 12 atti respiratori/minuto) = 4200 ml/minuto
194 Nelle situazioni in cui la domanda di ossigeno da parte dei tessuti aumenta, ad esempio durante l attività fisica, la ventilazione alveolare deve aumentare per far fronte a tale domanda. Ciò può essere ottenuto aumentando il volume corrente, oppure aumentando la frequenza respiratoria, oppure aumentando entrambi. La maniera più efficiente consiste nell aumentare il volume corrente piuttosto che nell aumentare la frequenza respiratoria, perché mentre all aumentare del ritmo respiratorio viene sottratto lo spazio morto ad ogni atto respiratorio all aumentare del volume corrente l aumento di volume va tutto ad aggiungersi all aria nuova che raggiunge gli alveoli. Ventilazione alveolare = (VC x frequenza respiratoria) (SM x frequenza respiratoria) Ventilazione alveolare = (500 x 12) (150 x 12)= =4200 Aumento del 60% del volume corrente: Ventilazione alveolare = (800 x 12) (150 x 12)= =7800 Aumento del 60% della frequenza respiratoria: Ventilazione alveolare = (500 x 20) (150 x 20)= =7000
195 CAPITOLO 17 Il sistema respiratorio: Lo scambio dei gas e la regolazione del respiro

196 quoziente respiratorio: rapporto tra la quantità di anidride carbonica prodotta dall organismo e la quantità di ossigeno consumata. A riposo, le cellule consumano 250 ml di ossigeno al minuto e producono 200 ml di anidride carbonica: 200 ml/250 ml=0.8 Il sangue deossigenato ritorna attraverso le vene sistemiche all atrio destro del cuore e da qui entra nel ventricolo destro che lo pompa ai polmoni attraverso le arterie polmonari, trasportando 750 ml di ossigeno e 2700 ml di anidride carbonica ai polmoni. Il sangue ossigenato lascia i polmoni e ritorna all atrio sinistro attraverso le vene polmonari. Entra quindi nel ventricolo sinistro, da cui viene pompato verso le cellule dell organismo attraverso le arterie sistemiche, trasportando nelle vene polmonari approssimativamente 1000 ml di ossigeno al minuto (750 ml/min. già nel sangue deossigenato ml/min. aggiunto dagli alveoli) e 2500 ml di anidride carbonica al minuto (2700 ml/min. presente nel sangue deossigenato 200 ml/min. lasciato negli alveoli). Nei capillari sistemici, 250 ml di ossigeno diffondono ogni minuto dal sangue alle cellule e 200 ml di anidride carbonica diffondono dalle cellule al sangue. Il sangue, ora deossigenato, ritorna all atrio destro ed il ciclo inizia nuovamente.
197 Il movimento di ossigeno e di anidride carbonica tra l aria alveolare e il sangue si ottiene per diffusione e dipende dal gradiente di concentrazione. - L ossigeno si trova ad una concentrazione maggiore negli alveoli e, quindi, diffonde nel sangue - l anidride carbonica si trova ad una maggiore concentrazione nel sangue e, di conseguenza, diffonde negli alveoli. L ossigeno e l anidride carbonica possono attraversare le membrane cellulari per semplice diffusione. La velocità del trasporto nella diffusione semplice è proporzionale al gradiente di concentrazione alla superficie della membrana attraverso cui la sostanza si muove e alla permeabilità della membrana per quella particolare sostanza.

198 La membrana respiratoria è composta da tre strati: le cellule epiteliali di tipo I nella parete alveolare le cellule endoteliali nella parete dei capillari e, tra di loro, le rispettive membrane basilari. La membrana respiratoria fornisce una superficie molto estesa di membrana estremamente sottile che favorisce una grande velocità di diffusione per l ossigeno e l anidride carbonica tra l aria alveolare e il sangue.
199 la pressione di un gas dipende dalla sua temperatura e dal numero di molecole di gas contenute in un dato volume. Un gas (l aria, per esempio) è spesso formato da una miscela composta da più di un tipo di molecole. La pressione totale di tale gas è data dalla somma delle pressioni dei singoli gas che compongono la miscela: P total = P 1 + P 2 + P P n Dove n indica il numero dei gas. pressione parziale: quota di pressione della miscela dovuta alla presenza di quel singolo gas Per esempio, se l elio e l azoto vengono mescolati in uguali proporzioni e la pressione della miscela è 500 mmhg, allora, metà pressione (250 mmhg) viene esercitata dall elio e l altra metà dall azoto. In questo esempio, 500 mmhg è la pressione totale della miscela e 250 mmhg è la pressione parziale di elio o di azoto.
200 GAS EXCHANGE
201 Le molecole di gas possono esistere in forma gassosa oppure trovarsi disciolte nei liquidi (IMPORTANTE! Altrimenti l ossigeno e l anidride carbonica non potrebbero essere trasportati dal sangue...) Anche quando le molecole di gas sono disciolte in un liquido esse esercitano una certa pressione parziale. Quando una miscela di gas viene in contatto con un liquido, le molecole di gas si dissolveranno nel liquido finché il sistema non raggiunge un equilibrio: situazione in cui il numero di molecole di gas che si dissolvono nel liquido equivale a quelle che, nello stesso intervallo di tempo, passano dal liquido allo stato gassoso. Quando il sistema è in equilibrio, le molecole di gas disciolte e quelle allo stato gassoso hanno la stessa pressione parziale. La concentrazione di molecole di gas nel liquido è proporzionale alla pressione parziale del gas e dipende anche dalla solubilità del gas in quel particolare liquido Ad una certa pressione parziale le concentrazioni relative di diversi gas disciolti differiscono da un gas all altro in quanto alcuni gas sono più solubili di altri. In altri termini, alcuni gas si mescolano con il liquido più facilmente di altri l anidride carbonica è circa 30 volte più solubile nel sangue dell ossigeno
202 La relazione tra la concentrazione, la pressione parziale e la solubilità di un gas è descritta dalla legge di Henry: c = kp c: concentrazione molare (moli di gas / litri di liquido) del gas disciolto P: pressione parziale del gas in atmosfere (1 atmosfera = 760 mmhg) k: costante della legge di Henry, che varia al variare del tipo di gas e della temperatura Sommozzatore che risale troppo rapidamente alla superficie. Sott acqua la pressione aumenta di 760 mmhg (un atmosfera), ogni 10 metri di profondità. I gas, specialmente l azoto, si sciolgono più velocemente nel sangue alle alte pressioni presenti nell acqua profonda. Se un sommozzatore risale in superficie troppo velocemente, quando la pressione diminuisce, nel sangue si formano bolle di azoto. Queste bolle di solito si fermano nelle articolazioni e nel sistema nervoso determinando nel primo caso problemi minori quali dolenzia durante la mobilizzazione articolare, nel secondo problemi gravi quali paralisi. Le bolle di gas possono anche occludere i vasi sanguigni provocando una condizione clinica nota come embolia gassosa. Risalendo in superficie poco alla volta i sommozzatori possono proteggersi da tali rischi permettendo all azoto di lasciare lentamente i tessuti in cui è disciolto.

203 Nelle miscele di gas, ciascun gas diffonde in base al suo gradiente di pressione parziale aria atmosferica PO 2 è 160 mmhg PCO 2 è 0.23 mmhg.
204 Nelle miscele di gas, ciascun gas diffonde in base al suo gradiente di pressione parziale aria atmosferica PO 2 è 160 mmhg PCO 2 è 0.23 mmhg. negli alveoli PO 2 è 100 mmhg PCO 2 è 40 mmhg. Scambi di gas Miscela aria vecchia e nuova Il sangue che entra nei capillari polmonari è sangue deossigenato PO 2 di 40 mmhg PCO 2 di 46 mmhg. Come questo sangue passa negli alveoli, l ossigeno e l anidride carbonica diffondono in base ai loro gradienti di pressione parziale: l ossigeno diffonde dagli alveoli al sangue mentre l anidride carbonica diffonde dal sangue agli alveoli In conclusione, la diffusione raggiunge un equilibrio tra l aria alveolare e il sangue capillare tale per cui il sangue che lascia i capillari polmonari e che entra nelle vene polmonari ha una PO 2 di 100 mmhg e una PCO 2 di 40 mmhg (le stesse pressioni parziali che si riscontrano negli alveoli).
205 La diffusione è un processo molto rapido, che si completa nel tempo in cui il sangue ha percorso circa un terzo della lunghezza dei capillari, affinché i gas possano ancora entrare in equilibrio tra il sangue capillare e l aria alveolare anche nel caso in cui il sangue scorra ad una velocità fino a tre volte quella normale, come succede, ad esempio, durante l attività fisica. La velocità di diffusione è rapida grazie alla sottigliezza della membrana respiratoria. Quando la membrana respiratoria risulta ispessita, lo scambio dei gas è impedito. Edema polmonare: l eccesso di fluido che occupa gli spazi interstiziali dei polmoni e gli alveoli aumenta la distanza tra gli alveoli e i capillari, aumentando in questo modo lo spessore della barriera di diffusione tra l aria e il sangue. determinando un sangue arterioso sistemico con una PO 2 più bassa e una PCO 2 più alta del normale. L organismo cercherà di compensare la diminuzione della PO 2 e l aumento di PCO 2 nel sangue arterioso, aumentando la ventilazione.
206 Il sangue ossigenato nei capillari polmonari ritorna all atrio sinistro attraverso le vene polmonari. quindi, fluisce nel ventricolo sinistro che lo pompa ai capillari sistemici dove ha luogo lo scambio tra le cellule del tessuto e il sangue. PO 2 di 100 mmhg PCO 2 di 40 mmhg. Nel fluido interstiziale che circonda i capillari PO 2 è più bassa a causa del consumo di ossigeno da parte delle cellule PCO 2 è più alta a causa della produzione di anidride carbonica da parte di esse l ossigeno si muove dal sangue ai tessuti e l anidride carbonica dai tessuti al sangue. Le pressioni parziali di ossigeno e anidride carbonica nel sangue venoso che lascia i capillari sistemici varia in base all attività metabolica del tessuto e al flusso di sangue al tessuto. Durante un intensa attività fisica, ad esempio, l attività metabolica del muscolo scheletrico è alta rispetto al suo flusso di sangue, e, quindi, nel fluido interstiziale attorno al tessuto muscolare la PO 2 sarà più bassa e la PCO 2 sarà più alta di quella presente in tessuti meno attivi. In presenza di gradienti di pressione più alti tra il sangue dei capillari e il fluido interstiziale, gli scambi di gas avvengono più rapidamente e un maggior numero di molecole di gas vengono scambiate. Tutto il sangue venoso ritorna all atrio destro mescolandosi prima di essere pompato dal ventricolo destro all arteria polmonare (sangue venoso misto). a riposo, PO 2 è 40 mmhg PCO 2 è 46 mmhg. Durante un intensa attività fisica, però, il valore di PO 2 diminuisce e quello di PCO 2 aumenta.
207 ACCOPPIAMENTO VENTILAZIONE-PERFUSIONE ha lo scopo di facilitare un efficiente scambio dei gas mantenendo il flusso d aria alveolare proporzionale al flusso di sangue nei capillari polmonari. EFFETTI DELLA PO2 SULLE ARTERIOLE (alta PO2 = vasodilatazione; bassa PO2 = vasocostrizione) grande flusso di aria agli alveoli alta PO2: arriva molto ossigeno agli alveoli e il sangue è poco per prenderlo tutto le arteriole si dilatano arriva più sangue agli alveoli permette al sangue di raccogliere l abbondante ossigeno nell aria che arriva agli alveoli poco flusso di aria negli alveoli bassa PO2: arriva poco ossigeno agli alveoli e il sangue ne prende in proporzione di più che nella norma le arteriole si costringono arriva meno sangue agli alveoli questo permette di deviare il sangue verso gli alveoli dove arriva aria molto ricca di ossigeno e la quantità di sangue è proporzionalmente adeguata a raccogliere l ossigeno che arriva. EFFETTI DELLA PCO2 SUI BRONCHIOLI (alta PCO2 = dilataz. bronch.; bassa PCO2 = costriz. bronch.) grande flusso di aria agli alveoli la PCO2 diminuisce perché in proporzione ne viene liberata poca dal sangue dei capillari alveolari i bronchioli si costringono: passa meno aria la PCO2 ritorna ai livelli normali poco flusso d aria agli alveoli la PCO2 aumenta perché in proporzione ne viene liberata molta dal sangue dei capillari alveolari i bronchioli si dilatano: passa più aria la dilatazione dei bronchioli permette di portar via tutta la CO2 liberata dal sangue alveolare
208 Se un piccolo tumore o altro riduce il flusso di aria agli alveoli: più bassa PO2 nell aria alveolare perché arriva poca aria e il sangue porta via molto ossigeno più alta PCO2 nell aria alveolare perché la poca aria che arriva non riesce a portare via tutta la CO2 liberata dal sangue quindi vasocostrizione: il sangue viene conservato per altri siti più ricchi di ossigeno e meno ossigeno viene portato via dal sangue i bronchioli si dilatano così il flusso di aria riesce a portar via tutta la CO2 liberata dal sangue risultato la PO2 si alza perché poco sangue porta via ossigeno (vasocostrizione) e perché in proporzione arriva più aria (dilatazione bronchioli) e quindi c è più ossigeno disponibile allo scambio la PCO2 si abbassa perché il gran flusso di aria (dilatazione bronchioli) riesce a portare via molta CO2 e perché meno sangue (vasocostrizione) libera CO2.
209 La PO 2 e la PCO 2 alveolare sono normalmente determinate da tre fattori: (1) la PO 2 e la PCO 2 dell aria inspirata non cambiano in modo apprezzabile ed è possibile considerarle costanti (2) la ventilazione alveolare al minuto (il volume di aria fresca che raggiunge gli alveoli ciascun minuto) (3) la velocità con la quale i tessuti respiratori consumano ossigeno e producono anidride carbonica. il fattore cruciale è la ventilazione alveolare relativa alla velocità di consumo di ossigeno e di produzione di anidride carbonica. ventilazione alveolare (o il flusso d aria) costante ma il consumo di ossigeno aumenta la PO 2 dell aria alveolare diminuisce. consumo di ossigeno costante ma la ventilazione alveolare diminuisce la PO 2 dell aria alveolare diminuisce se entrambi, il consumo di ossigeno e la ventilazione polmonare, aumentano in modo proporzionale la PO 2 alveolare non cambia.
210 Quando la ventilazione alveolare aumenta relativamente al consumo di ossigeno e alla produzione di anidride carbonica (ossia, quando la ventilazione alveolare supera le richieste dei tessuti), la PO 2 alveolare aumenta e la PCO 2 diminuisce. Quando la ventilazione alveolare diminuisce relativamente al consumo di ossigeno e alla produzione di anidride carbonica (ossia, quando la ventilazione alveolare non risponde alle richieste dei tessuti), la PO 2 alveolare diminuisce e la PCO 2 aumenta. Iperpnea: se il consumo di ossigeno e la produzione di anidride carbonica aumentano, allora anche la ventilazione polmonare aumenta. La PO 2 e la PCO 2 arteriose non si modificano. Ipoventilazione: la ventilazione polmonare non è sufficiente a rispondere alle richieste dei tessuti. PCO 2 arteriosa aumenta oltre il valore normale di 40 mmhg PO 2 arteriosa diminuisce sotto il valore normale di 100 mmhg Iperventilazione: la ventilazione alveolare supera le richieste dei tessuti PCO 2 arteriosa diminuisce sotto il valore di 40 mmhg PO 2 arteriosa aumenta oltre il valore di 100 mmhg
211 IL TRASPORTO DELL OSSIGENO NEL SANGUE Ciascun litro di sangue arterioso contiene circa 200 ml di ossigeno. Circa 3 ml di questo ossigeno (1.5%) è disciolto nel plasma o nel cytosol degli eritrociti, e solo questo ossigeno disciolto contribuisce alla Pressione parziale di ossigeno PO 2 nel sangue. I rimanenti 197 ml di ossigeno (98.5%) è trasportato legato alla emoglobina, una proteina che si trova negli eritrociti. Emoglobina consiste di quattro subunità, ciascuna delle quali contiene una globina (catena polipeptidica globulare) ed un gruppo eme, che contiene ferro. Ciascun gruppo eme è capace di legarsi ad una molecola di ossigeno, quindi, ciascuna molecola di emoglobina può trasportare quattro molecole di ossigeno. La quantità di ossigeno trasportato dall emoglobina è funzione della PO2, quindi dell ossigeno disciolto nel plasma. Emoglobina satura: quando tutti i siti di legame per l ossigeno sono occupati Ossiemoglobina: complesso costituito da emoglobina e dall ossigeno ad essa legato Desossiemoglobina: molecola di emoglobina priva di ossigeno.


212

213 Nei polmoni, le molecole di ossigeno che si spostano dall aria alveolare al sangue capillare, si legano all emoglobina quando il sangue raggiunge i tessuti respiratori, le molecole di ossigeno si dissociano dall emoglobina e diffondono alle cellule Il legame o il rilascio di ossigeno dipende dalla PO 2 del fluido che circonda l emoglobina: Alta PO 2 facilita il legame dell ossigeno con l emoglobina Bassa PO 2 facilita il rilascio dell ossigeno da parte dell emoglobina
214 L anemia è una diminuzione della capacità di trasportare ossigeno del sangue. Vi sono molte cause di anemia, inclusa una insufficienza o una mancanza di emoglobina. Con una minore capacità funzionale dell emoglobina nel sangue, la capacità di trasportare ossigeno diminuisce, e i tessuti non possono essere riforniti dell ossigeno che necessitano, anche quando la PO 2 del sangue è normale. Quindi, le persone che soffrono di anemia si stancano più facilmente.
, in quanto la capacità dell emoglobina di legarsi all ossigeno dipende da quante molecole di ossigeno sono già legate ad essa.")
215 curva di dissociazione emoglobina-ossigeno: grafico della percentuale di saturazione di emoglobina in funzione della PO 2 il legame tra ossigeno ed emoglobina non è rappresentato da una funzione lineare ma da una funzione a forma di S (sigmoide), in quanto la capacità dell emoglobina di legarsi all ossigeno dipende da quante molecole di ossigeno sono già legate ad essa. In particolare, il legame di una molecola di ossigeno all emoglobina aumenta l affinità della molecola di emoglobina per l ossigeno e quindi aumenta la probabilità che un altro ossigeno si leghi all emoglobina. A pressioni parziali molto basse (inferiori ai 15 mmhg, che di norma non si riscontrano nel sangue) molte molecole di emoglobina non hanno alcun ossigeno legato ad esse. In queste condizioni, l affinità dell emoglobina per l ossigeno è relativamente bassa, e un dato aumento nella PO 2 porta ad un piccolo aumento nella percentuale di saturazione. All aumentare della PO 2, (tra 15 mmhg e 60 mmhg) più molecole di emoglobina hanno legata almeno una molecola di ossigeno, determinando un aumento nell affinità dell emoglobina per altre molecole di ossigeno. Questa relazione è illustrata nella porzione ripida della curva di dissociazione emoglobinaossigeno A valori più alti di 60 mmhg, la pendenza della curva diminuisce in quanto, all aumentare della saturazione, è disponibile un minor numero di siti di legame. Ad un valore di PO 2 superiore agli 80 mmhg, la curva diventa pressoché piatta.
216 I cambiamenti nell affinità dell emoglobina per l ossigeno si riflettono in uno spostamento verso destra o verso sinistra della curva di dissociazione emoglobina-ossigeno Diminuzioni di affinità spostano la curva verso destra indicando che è necessaria una PO 2 più alta per ottenere qualsiasi livello di saturazione. L ossigeno viene scaricato più facilmente dall emoglobina, rendendolo più facilmente disponibile ai tessuti. Aumenti di affinità spostano la curva verso sinistra indicando che è sufficiente una PO 2 più bassa per ottenere qualsiasi livello di saturazione. L ossigeno viene caricato più facilmente dall emoglobina. Fattori che influenzano l affinità dell emoglobina per l ossigeno
217 1. Temperatura. All aumentare del metabolismo dei tessuti, la temperatura aumenta, diminuendo, quindi, l affinità dell emoglobina per l ossigeno. Più ossigeno viene scaricato nei tessuti molto attivi. La diminuzione della temperatura del sangue durante il passaggio nei polmoni, aumenta l affinità dell emoglobina per l ossigeno, promovendone il legame.
218 2. ph. (effetto Bohr) Gli ioni idrogeno si legano all emoglobina, e l ossigeno viene scaricato. Un aumento nella concentrazione di ioni idrogeno (una diminuzione di ph) diminuisce l affinità. La concentrazione di ioni idrogeno tende ad aumentare nel tessuto attivo, facilitando la liberazione di ossigeno.
219 3. PCO 2. Quando l anidride carbonica si lega all emoglobina, la conformazione dell emoglobina si modifica, diminuendo la sua affinità per l ossigeno Un aumento della PCO 2 nel sangue, come avviene quando l attività metabolica aumenta, sposta la reazione verso destra
220 4. 2,3-DPG. (2,3-difosfoglicerato), è un composto chimico prodotto negli eritrociti come composto intermedio della glicolisi, la via anaerobica grazie alla quale gli eritrociti ottengono l energia, la cui presenza diminuisce l affinità dell emoglobina per l ossigeno. monossido di carbonio: è tossico in quanto si lega più velocemente dell ossigeno all emoglobina, impedendo quindi il legame dell ossigeno e diminuendo, di conseguenza, la capacità di trasportare ossigeno del sangue.
221 IL TRASPORTO DELL ANIDRIDE CARBONICA NEL SANGUE Dell anidride carbonica presente nel sangue il 5-6% è disciolto (PCO2) il 5-8% è legata all emoglobina come carbaminoemoglobina l 86-90% è disciolta nel sangue sotto forma di ioni bicarbonato (HCO 3- ) (magazzino di CO2). Il bicarbonato viene formato a partire dall anidride carbonica negli eritrociti dei capillari sistemici.

222 Le cellule respiratorie producono anidride carbonica ad una velocità di circa 200 ml/min a riposo che deve essere rimossa dai sistemi circolatorio e respiratorio essa diffonde, in base al suo gradiente di pressione parziale, prima nel fluido interstiziale e poi nel plasma la PCO 2 aumenta, determinando un gradiente di pressione tra il plasma e gli eritrociti tale per cui la pressione nel plasma è maggiore l anidride carbonica diffonde dal plasma agli eritrociti quando il livello di ioni bicarbonato negli eritrociti aumenta, essi vengono trasportati nel plasma e scambiati con degli ioni cloro grazie ad una proteina da trasporto che si trova nella membrana degli eritrociti del plasma (scambio dei cloruri)

223 Nei polmoni, l anidride carbonica diffonde dal sangue che scorre nei capillari polmonari agli alveoli, per essere espirata, diminuendo, quindi, i livelli di anidride carbonica nel sangue La perdita di anidride carbonica fa in modo che gli ioni bicarbonato e idrogeno negli eritrociti si combinino e formino l acido carbonico, che viene convertito in anidride carbonica e acqua. L anidride carbonica, quindi, diffonde negli alveoli per essere espirata.
224 GAS TRANSPORT


225 Respirazione tranquilla: ciclo respiratorio consiste nella contrazione dei muscoli respiratori durante l inspirazione, seguita dal rilassamento degli stessi muscoli durante l espirazione. Respirazione più intensa: l espirazione diventa un processo attivo che richiede la contrazione dei muscoli espiratori. Quindi, durante la respirazione attiva: inspirazione: contrazione dei muscoli inspiratori e rilassamento dei muscoli espiratori espirazione: rilassamento dei muscoli inspiratori e contrazione dei muscoli espiratori esterno interno I muscoli respiratori sono muscoli scheletrici che vengono stimolati alla contrazione dall input nervoso proveniente dai neuroni motori somatici. Il nervo frenico innerva il diaframma I nervi intercostali interni ed esterni innervano i muscoli intercostali.
226 La respirazione è sia sotto controllo volontario che involontario. Quando parliamo, cantiamo, tratteniamo il respiro o sospiriamo per lo sgomento o l esasperazione, alteriamo il respiro in modo volontario. Per la maggior parte del tempo, però, respiriamo in modo automatico senza alcuno sforzo consapevole. Il controllo centrale della respirazione non è stato ancora compreso anche se sembra che i centri respiratori si trovino nel ponte e nel bulbo del tronco dell encefalo.

227 Nel ponte e nel bulbo del tronco dell encefalo si trovano due classi generali di neuroni, i neuroni inspiratori e i neuroni espiratori che generano potenziali d azione durante l inspirazione e l espirazione, rispettivamente. gruppo respiratorio ventrale (nel bulbo): contiene due regioni di neuroni espiratori primari ed una regione di neuroni inspiratori primari gruppo respiratorio dorsale (nel bulbo): contiene principalmente neuroni inspiratori, anche se sono presenti alcuni neuroni espiratori gruppo respiratorio pontino (nel ponte): contiene neuroni inspiratori, espiratori e neuroni misti, che hanno un attività legata sia all inspirazione che all espirazione
228 generatore di pattern centrale (sede e meccanismo d azione sconosciuti!): rete di neuroni che genera un pattern ripetuto e regolare di attività nervosa chiamato ritmo respiratorio. La respirazione dipende anche dall attività di altre regioni cerebrali, inclusa la corteccia cerebrale, il cervelletto, il sistema limbico, l ipotalamo e le aree di regolazione cardiovascolare del bulbo. Questo spiega, per esempio, perché i pattern respiratori cambiano quando una persona è in preda alla rabbia o alla paura.

229
230 Molti tipi di input sensoriale possono alterare la respirazione, probabilmente in seguito ad una comunicazione indiretta con il generatore di pattern centrale: chemocettori centrali e periferici (cellule recettoriali sensibili agli stimoli chimici) localizzati nel cervello e nelle arterie sistemiche. recettori da stiramento polmonari che si trovano nel muscolo liscio delle vie respiratorie polmonari i recettori nel rivestimento del tratto respiratorio: sono stimolati dalle particelle che vengono inalate quali il fumo o la polvere e da alcune sostanze chimiche. La stimolazione di questi recettori determina la broncocostrizione, l iperpnea, la tosse e lo starnuto, i quali, in certe condizioni, aiutano a liberare i polmoni da queste sostanze. La tosse ha inizio dai recettori nella trachea, mentre lo starnuto da quelli che si trovano nel naso e nella faringe. i propiocettori nei muscoli e nelle articolazioni (che dell organismo): riconoscono i movimenti promuovono l aumento in ventilazione che interviene durante l attività fisica. i barocettori arteriosi (che riconoscono i cambiamenti nella pressione del sangue) i nocicettori e i termocettori che si trovano in tutto il corpo.

231 I chemocettori controllano le pressioni parziali di ossigeno e di anidride carbonica nel sangue arterioso ed inviano tale informazione ai centri respiratori in modo tale che essi possano aggiustare la ventilazione in risposta ai cambiamenti di queste variabili. chemocettori periferici si trovano nei glomi carotidei vicino al seno carotideo cellule sensoriali specializzate, in contatto diretto con il sangue arterioso e comunicano (grazie alla secrezione di un messaggero chimico) con i neuroni afferenti che proiettano ai centri respiratori bulbari. Rispondono alle modificazioni della Po 2 arteriosa o del ph (che cambia al modificarsi della PCO 2 ). chemocettori centrali si trovano nel midollo allungato neuroni bulbari che rispondono direttamente ai cambiamenti della concentrazione degli ioni idrogeno nel fluido cerebrospinale che circonda questa struttura. l attivazione dei chemocettori determina un incremento di ventilazione una diminuzione nell attivazione dei chemocettori determina una diminuzione di ventilazione.

232 DRG = Dorsal Respiratory Group VRG = Ventral Respiratory Group
233 IL SISTEMA CARDIOVASCOLARE
234 CAPITOLO 13 Il sistema cardiovascolare: funzione cardiaca
235 Il sistema cardiovascolare è costituito da tre elementi: 1. il sangue - un fluido che porta sostanze alle cellule e ne allontana altre plasma: parte liquida del sangue (acqua, proteine, elettroliti e altro) e cellule eritrociti (globuli rossi): contengono emoglobina, proteina che trasporta O e CO2 leucociti (globuli bianchi): difendono il corpo dai microrganismi piastrine: frammenti di cellule che intervengono nella coagulazione 2. i vasi sanguigni - condotti attraverso cui il sangue circola arterie: dal cuore ai tessuti vene: dai tessuti al cuore differiscono per il diametro: arterie, arteriole, capillari, venule, vene e per lo spessore e la composizione della parete (arterie: pareti spesse ed elastiche (per contrastare la pressione del sangue); arteriole: anelli di muscolatura liscia (per regolare il flusso ematico); capillari: parete molto sottile (per permettere lo scambio dei gas) 3. il cuore - pompa muscolare che distribuisce il sangue nei vasi la sua principale funzione è fornire adeguate quantità di sangue a tutti gli organi e tessuti del corpo
. Ventricolo destro: sangue ai polmoni. Ventricolo sinistro: sangue a tutti gli organi del corpo.")
 Valvole semilunari separano i ventricoli dalle arterie: polmonare: tra il ventricolo dx e il tronco")
236 Gli atri ricevono il sangue dai vasi venosi e lo trasferiscono ai ventricoli che lo spingono verso i vasi arteriosi (aorta, tronco polmonare). Ventricolo destro: sangue ai polmoni. Ventricolo sinistro: sangue a tutti gli organi del corpo. Valvole atrioventricolari (valvole AV) separano gli atri dai ventricoli (sangue dall atrio al ventricolo): destra: tricuspide sinistra: bicuspide (valvola mitralica) Valvole semilunari separano i ventricoli dalle arterie: polmonare: tra il ventricolo dx e il tronco polmonare aortica: tra il ventricolo sx e l aorta.
237


238
239
240 Pressione dell atrio maggiore di quella del ventricolo: valvole aperte Pressione del ventricolo maggiore di quella dell atrio: valvole chiuse
241 Pressione del ventricolo maggiore di quella arteriosa: valvole aperte Pressione del ventricolo minore di quella arteriosa: valvole chiuse

242 Sistema circolatorio: circolo polmonare: cuore destro-polmone circolo sistemico: cuore sinistro-corpo ventricolo sx-sangue ossigenato valvola aortica aorta tessuti-sangue deossigenato vene cave atrio destro valvola tricuspide ventricolo destro valvola polmonare arterie polmonari polmoni-sangue ossigenato vene polmonari atrio sinistro valvola mitralica ventricolo sinistro

243 ATTIVITA ELETTRICA DEL CUORE: il muscolo cardiaco deve contrarsi in maniera sincrona: prima entrambi gli atri e poi entrambi i ventricoli. Il muscolo cardiaco ha una struttura a sarcomeri come il muscolo scheletrico ed è dotato di giunzioni comunicanti: regioni in cui due cellule adiacenti sono connesse da proteine di membrana (connessoni) che formano dei canali che permettono il passaggio di ioni e di piccole molecole tra le cellule. Questo permette l accoppiamento elettrico di cellule adiacenti: quando le proprietà elettriche di una cellula cambiano, il flusso ionico attraverso le giunzioni comunicanti induce una modificazione dello stato elettrico nelle cellule adiacenti. Grazie a questo meccanismo le cellule del cuore possono funzionare in maniera coordinata contraendosi all unisono in quanto il pot d az una volta generato si propaga a tutta la rete cellulare.
244 ATTIVITA MIOGENA DEL CUORE: Le contrazioni del muscolo cardiaco sono indotte da segnali che originano all interno del muscolo stesso e non richiedono stimoli da parte del S.N.C. (come succede per il muscolo scheletrico, attività neurogena=il segnale iniziale origina nel neurone). CELLULE AUTORITMICHE: Piccola percentuale di cellule muscolari modificate che generano una forza contrattile piccola o nulla MA CHE SONO SPECIALIZZATE A INIZIARE E/O CONDURRE I POTENZIALI D AZIONE che provocano le contrazioni del muscolo cardiaco: 1. Cellule pacemaker (segnaritmo) che avviano i potenziali d azione 2. Fibrocellule del sistema di conduzione che consentono ai potenziali d azione di propagarsi lungo il cuore in maniera estremamente coordinata. I potenziali d azione viaggiano ad una velocità superiore ai 4 m/sec (diametro maggiore) CELLULE CONTRATTILI: Cellule che generano la forza contrattile. I potenziali viaggiano ad una velocità di 0,3-0,5 m/sec
.")
245 Cellule cardiache con attività pacemaker (generano depolarizzazioni spontanee): ATTIVITA MIOGENICA nodo senoatriale nodo atrioventricolare Le contrazioni del muscolo cardiaco sono, in condizioni normali, attivate solo dalle cellule del nodo SA ( il nodo AV dà inizio alla contrazione solo nel caso in cui il SA non funzioni bene). I potenziali d azione generati dal nodo SA arrivano all AV determinando il periodo refrattario assoluto. La frequenza del SA è maggiore di quella dell AV (lo «batte» sempre!). Le cellule pacemaker, sono associate ad altre fibre muscolari, le fibre di conduzione, specializzate nel condurre velocemente i pot d az lungo il miocardio: sono di diametro maggiore e quindi conducono i pot d az più rapidamente. SISTEMA DI CONDUZIONE DEL CUORE: cellule pacemaker + fibre di conduzione esso provoca un onda di eccitazione, che si muove prima attraverso gli atri (depolarizzazione e contrazione) e poi attraverso i ventricoli (depolarizzazione e contrazione).

246 1. Inizia il pot d az nel nodo SA e attraverso la via internodale (fibre di conduzione) si diffonde agli atri. 2. Pot d az raggiunge il nodo AV che rallenta la propagazione del segnale (è fondamentale per consentire all onda di eccitazione di diffondere completamente attraverso gli atri prima di raggiungere i ventricoli) 3. Prosegue lungo il fascio atrio-ventricolare (fascio di His) (sola connessione elettrica tra atri e ventricoli) 4. Il fascio si divide in branca destra, che porta l impulso al ventricolo dx, e branca sinistra, che porta l impulso al ventricolo sx 5. Le branche si ramificano in fibre del Purkinje che permettono la diffusione del segnale nei ventricoli

247 Quindi la contrazione ventricolare inizia all apice e diffonde verso l alto, dove il sangue lascia i ventricoli Il rallentamento è essenziale in quanto consente all onda di eccitazione di diffondere completamente attraverso gli atri prima di raggiungere i ventricoli


248 ATTIVITA ELETTRICA DELLE CELLULE PACEMAKER Perché una cellula pacemaker genera spontaneamente pot d az? Perché non riesce a mantenere ad un livello stabile il proprio pot di membrana. Dopo un pot d az la cellula inizia subito una lenta depolarizzazione che raggiunge la soglia e innesca un altro pot d az quindi il pot di membrana ritorna a -60/-70mV e ricomincia a depolarizzarsi la lenta depolarizzazione e il pot d az sono generati dalle variazioni di permeabilità al K+ (dentro), al Na+ (fuori) e al Ca++ (fuori).
249
250 ATTIVITA ELETTRICA DELLE CELLULE PACEMAKER Il sistema nervoso autonomo può modificare la forza e il ritmo delle contrazioni del cuore in quanto agisce direttamente sulla permeabilità agli ioni delle cellule pacemaker: neuroni simpatici: aumento P al Na+ e al Ca++ neuroni parasimpatici: aumento P al K+ e diminuzione P al Na+ e al Ca++
251 I pot d az cardiaci delle cellule contrattili sono diversi da quelli a punta dei muscoli scheletrici e dei neuroni (durata 1-2 msec) perché sono ampi durano centinaia di msec: ossia quasi tutto il tempo in cui le cellule cardiache si contraggono e si rilasciano Questo impedisce che nel muscolo cardiaco si sommino le contrazioni, anche quando la frequenza dei potenziali è elevata e il cuore batte rapidamente. Se questo succedesse, il cuore perderebbe la capacità di rilasciarsi completamente tra una contrazione e l altra per potersi riempire di sangue.
252 ATTIVITA ELETTRICA DELLE CELLULE CARDIACHE CONTRATTILI A seconda della sede della cellula contrattile i pot d az possono essere diversi nella forma e nella velocità di propagazione in quanto le cellule differiscono per dimensioni e tipo e numero di canali ionici. 0. Depolarizzazione: apertura canali Na+ voltaggio-dipendenti (Na+ entra): depolarizzazione 1. Inattivazione canali Na+. La fase di ripolarizzazione dura di più perché chiusura canali K+ (K+ non esce) e apertura canali Ca++ (Ca++ entra: oltre a contribuire al platau, contribuisce alla liberazione di Ca++ da parte del RS- liberazione di Ca calcio-indotta) 2. Plateau 3. Pk aumenta: il flusso di K+ in uscita porta il pot di membrana verso valori più negativi. Diminuzione di PCa (il Ca++ non entra più) 4. Potenziale di riposo
253 Accoppiamento eccitazione-contrazione E praticamente uguale a quello del muscolo scheletrico: pot d az si propaga lungo i tubuli a T apertura canali Ca++ voltaggio-dipendenti sul RS Ca++ si fissa alla troponina la tropomiosina si sposta dai siti di fissazione per la miosina ciclo dei ponti trasversali il Ca++ che entra nella cellula durante la fase di plateau si fissa alla troponina agisce sui canali voltaggio-dipendenti sul RS determinando una maggiore liberazione di Ca++ (liberazione del Ca++ calcio-indotta) perché il muscolo cardiaco si rilasci è necessaria la rimozione del Ca++ che avviene tramite diversi meccanismi che attivamente riportano il Ca++ dentro il RS

254
255 L elettrocardiogramma registra il flusso di corrente elettrica che attraversa il cuore durante un ciclo cardiaco, per mezzo di elettrodi collocati sulla cute. Triangolo di Einthoven: 3 elettrodi che vengono collegati 2 a 2. In ogni coppia, un elettrodo viene designato come positivo e uno come negativo. Le coppie sono riferite come derivazioni (I, II, III) e ciascuna misura la differenza di potenziale elettrico di superficie tra l elettrodo positivo e quello negativo.
256


257 Eccitamento cardiaco correlato all elettrocardiogramma Il nodo SA genera un impulso: incomincia l eccitamento atriale Impulso ritardato al nodo AV onda P finisce L impulso passa all apice del cuore: incomincia l eccitamento ventricolare L eccitamento ventricolare è completo complesso QRS Nodo SA Nodo AV Rami del fascio di His Fibre del Purkinje R P L onda T, che fa seguito all onda S, corrisponde alla ripolarizzazione ventricolare Q Q S

258
259 IL CICLO CARDIACO (cuore sinistro) viene diviso in sistole: contrazione ventricolare (fasi 2 e 3) diastole: rilasciamento ventricolare (fasi 4 e 1) 1. Sia gli atri che i ventricoli sono rilasciati. Valvole AV aperte, il sangue passa ai ventricoli. Valvole semilunari chiuse: la pressione del ventricolo è più bassa di quella delle arterie. Gli atri si contraggono spremendo il sangue residuo nei ventricoli. Gli atri si rilasciano. Aperte 2. Sistole ventricolare: la pressione dei ventricoli supera quella atriale, le valvole AV si chiudono. Le valvole semilunari sono chiuse perché la pressione ventricolare è ancora più bassa di quella arteriosa. Il sangue non può andare da nessuna parte: contrazione isovolumetrica 3. P ventricolare > P arteriosa: valvole semilunari si aprono. Volume ventricolare diminuisce perché il sangue passa nelle arterie: eiezione ventricolare. La pressione ventricolare diminuisce. 4. Chiusura valvole semilunari. Rilassamento ventricoli: rilasciamento isovolumetrico perché un po di sangue è presente nei ventricoli. 1. Quando la pressione ventricolare diventa più bassa di quella atriale le valvole AV si aprono e il ciclo si ripete: riempimento ventricolare
260 Aumento di pressione nei ventricoli perché la sistole atriale spreme il sangue residuo nei ventricoli e questo determina un aumento di pressione Pressione vicina allo zero perché il ventricolo è rilasciato Lento innalzarsi della pressione dovuto al riempimento passivo
261 Valvola aortica chiusa. Il sangue fluisce continuamente verso i vasi arteriosi più a valle. Lenta diminuzione di volume, lenta diminuzione di pressione La valvola aortica si apre, aumenta il flusso, aumenta la pressione Minimo di pressione aortica Diminuisce il flusso di sangue, diminuisce la pressione Chiusura della valvola aortica per il crollo della pressione ventricolare (diastole). La pressione aortica rimane più elevata di quella ventricolare perché le sue pareti elastiche immagazzinano la pressione durante la sistole. Durante la diastole il flusso sanguigno nei vasi a valle è assicurato grazie a questa pressione immagazzinata dall aorta. Massimo di pressione aortica

262 Contrazione atriale Volume costante perché tutte le valvole sono chiuse Rapida diminuzione del volume in seguito all apertura della valvola semilunare Volume aumenta perché le valvole AV sono aperte Massimo volume ventricolare raggiunto durante il ciclo cardiaco EDV - ESV: volume di sangue pompato dal cuore ad ogni battito cardiaco Minimo volume ventricolare raggiunto durante il ciclo cardiaco Alla fine della sistole ventricolare, nel ventricolo rimane un po di sangue (circa la metà, 65 ml). La quantità di sangue che rimane nel ventricolo dipende dalla forza di contrazione del cuore (modulata dal sistema nervoso autonomo e dagli ormoni). > forza = > volume di eiezione ventricolare perché diminuisce il volume telesistolico < forza = < volume di eiezione ventricolare. Rimane più sangue nel ventricolo. Frazione di eiezione: la porzione di volume telediastolico che viene espulsa dal ventricolo.

263 Ad ogni battito cardiaco si possono sentire due toni, uno delicato e di bassa frequenza, detto primo tono cardiaco ed uno più rumoroso ed acuto, detto secondo tono cardiaco. Questi toni coincidono con l inizio della sistole, quando le valvole AV si chiudono e con l inizio della diastole, quando le valvole semilunari si chiudono e sono provocati dal sangue che si precipita turbolento tra i loro lembi che si avvicinano.
264 il sangue - è composto da due costituenti plasma: parte liquida del sangue (acqua, proteine, elettroliti e altro) e parte corpuscolata eritrociti (globuli rossi): contengono emoglobina, proteina che trasporta O e CO2 leucociti (globuli bianchi): difendono il corpo dai microrganismi piastrine: frammenti di cellule che intervengono nella coagulazione PLASMA Piccoli soluti: sono simili a quelli del liquido interstiziale perché le pareti dei capillari sono altamente permeabili ad essi. Proteine: molto più concentrate nel liquido interstiziale perché non passano. Albumine: sintetizzate nel fegato. Sono le più concentrate e quindi contribuiscono molto alla pressione osmotica del plasma. Globuline: di vario tipo. Trasportano varie sostanze. Intervengono nella formazione dei coaguli.ruolo nella difesa da sostanze estranee e microrganismi. Fibrinogeno: sintetizzato nel fegato. Coagulazione del sangue.
265 PARTE CORPUSCOLATA le cellule del sangue vengono prodotte dal midollo osseo. Eritrociti: i globuli rossi sono le cellule più abbondanti. Trasportano ossigeno e anidride carbonica grazie alla presenza dell emoglobina che contiene ferro. La produzione di eritrociti (eritropoiesi) è regolata dall ormone eritropoietina, secreto dalle cellule renali e liberato in circolo, che raggiunge il midollo osseo. Gli eritrociti vivono 120 giorni nel sangue e quindi vengono rimossi e distrutti ad opera del fegato e della milza e il prodotto della loro degradazione è la bilirubina (colore giallo delle urine). Per la produzione degli eritrociti è importante il ferro (emoglobina) e l acido folico e la vitamina B12 (necessarie per la sintesi del DNA e la divisione cellulare). Qualsiasi carenza può portare ad anemia (ridotta capacità di trasportare ossigeno del sangue). Anemia da carenza di ferro: poco ferro. Anemia perniciosa: poca vitamina B12 o poco acido folico. L anemia può anche essere causata da emorragia o da eccessiva velocità di degradazione degli eritrociti. Occupano circa il 45% del volume totale di sangue: ematocrito (contributo percentuale). Leucociti: i globuli bianchi sono circa 1000 volte meno numerosi di quelli rossi. Contribuiscono a difendere l organismo dai microrganismi e da altri agenti estranei. Si trovano anche in altri tessuti perché possono passare attraverso i pori dei capillari e raggiungere le zone infette. Piastrine: frammenti di cellule che originano quando porzioni di grosse cellule del midollo osseo (megacariociti) vengono degradate. Sono importanti nell attivare la sequenza degli eventi che porta alla coagulazione del sangue.
 e dal tipo di flusso (turbolento o laminare). Raggio: al diminuire del raggio aumenta la resistenza.")
266 La resistenza in un tubo è una misura del grado con cui il tubo ritarda o resiste al flusso del liquido che lo percorre e dipende dalle dimensioni fisiche del tubo (raggio), dalle proprietà del fluido (viscosità) e dal tipo di flusso (turbolento o laminare). Raggio: al diminuire del raggio aumenta la resistenza. Viene controllato dalla muscolatura liscia delle piccole arterie e arteriole (vasodilatazione e vasocostrizione). Viscosità ematica: il principale determinante è l ematocrito. Se aumenta (per aumento numero eritrociti (policitemia) o diminuzione volume plasmatico (disidratazione)), aumenta la viscosità. Tipo di flusso: il sangue si muove principalmente con un flusso di tipo laminare. Scorre tranquillo lungo i vasi e quello che scorre al centro scorre più velocemente perché le pareti esercitano una resistenza su quello che scorre vicino ad esse. Se viene spinto a muoversi velocemente, il flusso diviene turbolento e il sangue scorre con direzioni e velocità diverse nei diversi punti del vaso. Queste variazioni aumentano la resistenza e creano vibrazioni che possono essere avvertite sotto forma di rumore.
267 Frequenza cardiaca: n /min numero di contrazioni ventricolari al minuto Volume di eiezione ventricolare: l volume di sangue pompato da ciascun ventricolo ad ogni battito. Gittata cardiaca: l/min velocità con la quale un ventricolo pompa sangue = frequenza cardiaca x volume di eiezione ventricolare Tutto ciò che influenza la frequenza cardiaca o il volume di eiezione ventricolare, influenza la gittata cardiaca. In un soggetto adulto a riposo: 72 battiti/min x 70 ml (0,07 l) = 5 l/min Il volume totale di sangue nel corpo è circa 5 litri: in un solo minuto il ventricolo è in grado di pompare l intero volume di sangue.
268 Influenze sulla frequenza cardiaca: SN simpatico afferenze al nodo SA: aumento stimolazione simpatica = aumento frequenza pot d az = aumento frequenza cardiaca aumento gittata cardiaca al nodo AV e ad altre porzioni del sistema di conduzione aumento della velocità di conduzione dei pot d az: il ventricolo si contrae prima e più rapidamente (=sistole più breve). SN parasimpatico afferenze al nodo SA: aumento stimolazione parasimpatica = diminuzione frequenza pot d az = diminuzione frequenza cardiaca diminuzione gittata cardiaca al nodo AV e ad altre porzioni del sistema di conduzione diminuzione della velocità di conduzione dei pot d az: il ventricolo si contrae dopo e più lentamente (=sistole più lunga). Il sistema simpatico e quello parasimpatico sono sempre attivi simultaneamente ma se l attività simpatica aumenta, diminuisce quella parasimpatica e viceversa. Questo tipo di regolazione permette che le modificazioni di attività di ciascun sistema si rinforzino a vicenda, visto che hanno effetti opposti (> simpatico e < parasimpatico = aumento frequenza cardiaca). Adrenalina viene secreta dalla midollare del surrene. aumento frequenza pot d az = aumento frequenza cardiaca aumento gittata cardiaca aumento della velocità di conduzione dei pot d az attraverso le fibre muscolari cardiache.
269 Influenze sul volume di eiezione ventricolare: Il volume di eiezione ventricolare è influenzato da: a) la forza di contrazione ventricolare 1. La contrattilità ventricolare (misura della capacità del ventricolo di generare pressione) SN simpatico: innerva tutto il miocardio. Aumentando la quantità di calcio che entra nella cellula ad ogni pot d az, aumenta la forza di contrazione degli atri (passa più sangue nei ventricoli) e dei ventricoli (aumenta la gittata cardiaca). Adrenalina: aumento della forza di contrazione e quindi del volume di eiezione ventricolare e quindi della gittata cardiaca. 2. la forza di contrazione ventricolare dipende anche da quanto il miocardio ventricolare viene stirato quando il ventricolo si riempie di sangue: controllo intrinseco effetto Starling: se aumenta il volume telediastolico (volume di sangue contenuto nel ventricolo alla fine della diastole), la forza di contrazione ventricolare aumenta, aumenta il volume di eiezione ventricolare e quindi la gittata cardiaca legge del cuore di Starling: quando il ritorno venoso (volume di sangue che entra nel cuore proveniente dai vasi venosi) varia, il cuore regola automaticamente il flusso in uscita per adattarlo a quello in entrata. L aumento del volume telediastolico provoca l allungamento delle fibre muscolari del miocardio ventricolare e, al contrario di quello che succede nel muscolo scheletrico, questo determina un aumento della forza di contrazione perché a. la lunghezza ottimale del sarcomero per il muscolo cardiaco è superiore alla lunghezza di riposo b. lo stiramento delle fibre muscolari aumenta l affinità della troponina per il calcio e quindi aumenta il numero di ponti trasversali attivi ad ogni contrazione.
270 3. Volume telediastolico (volume di sangue contenuto nel ventricolo alla fine della diastole) in accordo con la legge di Starling il volume di eiezione ventricolare aumenta o diminuisce all aumentare o diminuire del volume telediastolico. Il volume telediastolico è determinato dalla precarico: pressione del sangue sospinto all interno del ventricolo alla fine della diastole (aria insufflata in un palloncino determina la pressione all interno del palloncino) Influenze sul volume di eiezione ventricolare: Il volume di eiezione ventricolare è influenzato da: b) la forza che si oppone al passaggio del sangue nei tronchi arteriosi 1. L aumento della pressione arteriosa (postcarico) tende a far diminuire il volume di eiezione ventricolare.
271
272 Vene stesso diametro delle arterie, ma la loro parete ha uno spessore che è circa la metà. Hanno elevata complianza: pareti sottili e facilmente estensibili: magazzino di volume. Possono perdere un sostanziale volume di sangue prima che la diminuzione di pressione abbia una conseguenza sulla gittata cardiaca.

273 MISURAZIONE DELLA PRESSIONE SANGUIGNA si basa sul fenomeno della turbolenza e del rumore che la accompagna. Sfigmomanometro: bracciale gonfiabile e misuratore di pressione all interno del bracciale + stetoscopio Il bracciale viene gonfiato finché la pressione al suo interno non supera sicuramente la pressione arteriosa sistolica e quindi l arteria brachiale viene schiacciata e il flusso sanguigno al suo interno viene interrotto. Si riduce lentamente la pressione all interno del bracciale. Quando si sente un rumore, significa che la pressione nel bracciale è leggermente inferiore alla pressione arteriosa sistolica e quindi il sangue riesce a passare con flusso turbolento. Mano a mano che la pressione nel bracciale scende, il flusso si fa sempre meno turbolento (raggio >) finché i suoni scompaiono: pressione diastolica.
274 STRUTTURA E FUNZIONE DEI VASI SANGUIGNI Arterie allontanano il sangue dal cuore e lo conducono ai tessuti. Diametro e spessore pareti grandi. Pareti hanno grosse quantità di tessuto fibroso ed elastico. Agiscono come serbatoio di pressione perché hanno poca complianza: non si espandono molto, aumenta la pressione perché non aumenta il volume. Vene stesso diametro delle arterie, ma la loro parete ha uno spessore che è circa la metà. La pressione è significativamente più bassa che nelle arterie. Possiedono valvole unidirezionali che consentono il fluire di sangue verso il cuore e non il reflusso verso gli organi e tessuti. Hanno elevata complianza: pareti sottili e facilmente estensibili: magazzino di volume. Possono perdere un sostanziale volume di sangue prima che la diminuzione di pressione abbia una conseguenza sulla gittata cardiaca. Arteriole portano il sangue dalle arterie ai capillari. La loro funzione è di regolare il flusso attraverso il letto capillare posto a valle grazie alla contrazione o il rilasciamento della muscolatura liscia delle pareti che vengono regolati dal sistema nervoso autonomo, da agenti chimici e da ormoni. Capillari sono i vasi sanguigni più piccoli e più numerosi. La sottile parete dei capillari facilita la loro funzione principale: scambio di sostanze tra sangue e cellule tissutali che nella maggior parte dei tessuti avviene mediante diffusione semplice. Venule sono leggermente più piccole delle arteriole. Contengono poca o nessuna muscolatura liscia. Partecipano allo scambio di sostanze e portano sangue dai capillari alle vene.

275 Il circolo polmonare e quello sistemico sono posti in serie, ossia il sangue deve passare in sequenza attraverso i due circuiti per ritornare al punto di partenza. Ma se analizziamo il flusso ematico in ciascuno dei due circoli, il flusso in essi è in parallelo: il sangue non scorre in sequenza da un organo all altro, ma scorre attraverso l aorta nelle arterie che da essa si diramano ognuna verso un organo e lo stesso all interno di ciascun organo. Vantaggi: ogni organo riceve sangue ricco di ossigeno e non depauperato da un altro organo. È possibile regolare il flusso di sangue ai diversi organi, indipendentemente uno dall altro (grazie alle arteriole e alla muscolatura liscia delle loro pareti).
276 CAPITOLO 14 Il sistema cardiovascolare: Vasi sanguigni, flusso e pressione del sangue
 è di circa 2-8 mmhg.")
277 All inizio del circolo sistemico la pressione arteriosa media (MAP la pressione media che vige nell aorta durante tutto il ciclo cardiaco) è di circa 85 mmhg. Alla fine del circolo, nelle grosse vene della cavità toracica che riportano il sangue al cuore, la pressione, definita pressione venosa centrale (CVP) è di circa 2-8 mmhg. Il gradiente pressorio che consente al sangue di fluire nel circolo sistemico è dato dalla differenza tra la pressione arteriosa media e la pressione venosa centrale. Poiché la pressione venosa centrale è molto bassa, possiamo assumere che Il gradiente di pressione ( P) che permette il flusso del sangue attraverso il circolo sistemico sia virtualmente identico alla pressione arteriosa media.
278 LEGGI FISICHE CHE REGOLANO IL FLUSSO E LA PRESSIONE DEL SANGUE flusso di sangue = gradiente di pressione / resistenza Grazie alla sua azione di pompa il cuore innalza la pressione arteriosa media, creando una differenza di pressione tra le arterie e le vene che consente al cuore di scorrere. La pressione diminuisce gradualmente mano a mano che il sangue scorre dalle arterie alle vene (caduta di pressione). CIRCOLO SISTEMICO Inizio: pressione arteriosa media (pressione media nell aorta durante il ciclo cardiaco) = 90 mmhg Fine: pressione venosa centrale (nelle vene toraciche che vanno al cuore) = 0 mmhg Quindi, il gradiente di pressione che permette il flusso di sangue attraverso il circolo sistemico è virtualmente identico alla pressione arteriosa media. CIRCOLO POLMONARE Inizio: pressione media nell arteria polmonare = 15 mmhg Fine: pressione venosa polmonare = 0 mmhg. Il gradiente di pressione è inferiore a quello del circolo sistemico. Siccome il flusso ematico attraverso il circolo sistemico è uguale a quello attraverso il circolo polmonare (5 litri/min) e il gradiente di pressione è maggiore nel circolo sistemico, ciò significa che il circolo polmonare offre minor resistenza!
279 La resistenza del sistema vascolare dipende dalla resistenza di tutti i singoli vasi che lo compongono. Nel circolo sistemico, le resistenze combinate di tutti gli organi e dei vasi vengono dette resistenza periferica totale (TPR) ma il contributo principale viene dato dalle arteriole e dalle piccole arterie. Di norma resta costante perché la vasocostrizione di un distretto annulla la vasodilatazione di un altro. Nel circolo sistemico possiamo sostituire alla formula: flusso = gradiente di pressione / resistenza flusso: volume di sangue al minuto = gittata cardiaca gradiente di pressione: pressione arteriosa media - pressione venosa centrale = 90 mmhg - 0 mmhg = pressione arteriosa media resistenza: resistenza periferica totale gittata cardiaca = pressione arteriosa media / resistenza periferica totale CO = MAP / TPR
280 In assenza di variazioni della resistenza dei singoli organi, ogni variazione della pressione arteriosa media influenza allo stesso modo il flusso ematico di ciascun organo, facendo sì che la quota percentuale della gittata cardiaca diretta a ciascun organo non cambi. pressione arteriosa media (MAP) Pressione venosa centrale (CVP) Il flusso resta uguale Si modificano Il flusso diminuisce diminuisce
 / resistenza periferica totale (TPR) CO = MAP / TPR MAP = CO x TPR CO = HR (frequenza cardiaca) x SV (volume di eiezione ventricolare) MAP = HR x SV x TPR Quindi la pressione arteriosa media")
281 Dovuto ad aumento HR o SV Abbiamo visto che la relazione flusso = gradiente di pressione / resistenza nel circolo sistemico può essere indicata con gittata cardiaca (CO) = pressione arteriosa media (MAP) / resistenza periferica totale (TPR) CO = MAP / TPR MAP = CO x TPR CO = HR (frequenza cardiaca) x SV (volume di eiezione ventricolare) MAP = HR x SV x TPR Quindi la pressione arteriosa media dipende da: frequenza cardiaca HR volume di eiezione ventricolare SV resistenza periferica totale TPR che vengono regolati mediante meccanismi riflessi che coinvolgono il sistema nervoso autonomo.
POTENZIALE DI MEMBRANA A RIPOSO. Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI
 POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
Laila Craighero. Sezione di Fisiologia umana Università di Ferrara
 Laila Craighero Sezione di Fisiologia umana Università di Ferrara crh@unife.it Oppure scritto Se volete consultare un testo semplice: Fisiologia Germann Stanfield Edizione: III / 2009 Pubblicato da Edises
Laila Craighero Sezione di Fisiologia umana Università di Ferrara crh@unife.it Oppure scritto Se volete consultare un testo semplice: Fisiologia Germann Stanfield Edizione: III / 2009 Pubblicato da Edises
Il neurone. I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi.
 attraverso le sinapsi.") Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
MUSCOLO. Muscolo striato scheletrico. FGE aa
 MUSCOLO Muscolo striato scheletrico FGE aa.2015-16 Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli
MUSCOLO Muscolo striato scheletrico FGE aa.2015-16 Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli
Principali differenze tra sinapsi neuromuscolare e sinapsi centrali
 Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Durata P. A. [ms] propriocettive muscolari A(β) Tatto, cinestesia,
![Durata P. A. [ms] propriocettive muscolari A(β) Tatto, cinestesia,](/thumbs/63/48783499.jpg "Durata P. A. [ms] propriocettive muscolari A(β) Tatto, cinestesia,") TIPI DI FIBRE NERVOSE NEI MAMMIFERI Durata P. A.: durata del potenziale d azione; periodo R. A.: periodo refrattario assoluto; nrd: nervi delle radici dorsali; ns: nervi simpatici Fibra Diametro Velocità
TIPI DI FIBRE NERVOSE NEI MAMMIFERI Durata P. A.: durata del potenziale d azione; periodo R. A.: periodo refrattario assoluto; nrd: nervi delle radici dorsali; ns: nervi simpatici Fibra Diametro Velocità
giunzioni comunicanti connessone cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali precoci veloci bidirezionali
 La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA
 LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA Lezione_6 1 STUTTURA MOTONEURONE FIBRA MUSCOLARE GIUNZIONE NMUSCOLARE Lezione_6 2 Sintesi e degradazione di acetilcolina ACh è rilasciata da
LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA Lezione_6 1 STUTTURA MOTONEURONE FIBRA MUSCOLARE GIUNZIONE NMUSCOLARE Lezione_6 2 Sintesi e degradazione di acetilcolina ACh è rilasciata da
La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi elettro-chimici.
 LA TRASMISSIONE SINAPTICA Il neurone : unità elementare responsabile dei messaggi nervosi La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi
LA TRASMISSIONE SINAPTICA Il neurone : unità elementare responsabile dei messaggi nervosi La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi
Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata.
 Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata. Costante e stabile Negativa all interno Peculiare di ogni tipo di cellula Il Potenziale di
Potenziale di riposo: Una ddp a cavallo della membrana plasmatica a riposo o quando la cellula non è stimolata. Costante e stabile Negativa all interno Peculiare di ogni tipo di cellula Il Potenziale di
Differenze tra sinapsi neuromuscolare e sinapsi centrali
 Sinapsi centrali ed Integrazione sinaptica Differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore solo Ach. Le sinapsi centrali utilizzano
Sinapsi centrali ed Integrazione sinaptica Differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore solo Ach. Le sinapsi centrali utilizzano
Comunicazione tra cellule
 Comunicazione tra cellule I segnali-messaggi possono essere: elettrici o chimici Il messaggio deve avere le seguenti caratteristiche: 1. Contenere informazione 2. Essere indirizzato 3. Essere trasmesso
Comunicazione tra cellule I segnali-messaggi possono essere: elettrici o chimici Il messaggio deve avere le seguenti caratteristiche: 1. Contenere informazione 2. Essere indirizzato 3. Essere trasmesso
La trasmissione dell Impulso Nervoso
 La trasmissione dell Impulso Nervoso Il Neurone Neuroni: cellule specializzate per la trasmissione degli impulsi. Dendriti prolungamenti riccamente ramificati e un lungo Assone fibroso. La parte terminale
La trasmissione dell Impulso Nervoso Il Neurone Neuroni: cellule specializzate per la trasmissione degli impulsi. Dendriti prolungamenti riccamente ramificati e un lungo Assone fibroso. La parte terminale
TESSUTO E SISTEMA MUSCOLARE
 TESSUTO E SISTEMA MUSCOLARE Panoramica del tessuto muscolare Ci sono tre tipi di tessuto muscolare Scheletrico (striato) : è attaccato alle ossa e muove parti dello scheletro; movimenti volontari e alcuni
TESSUTO E SISTEMA MUSCOLARE Panoramica del tessuto muscolare Ci sono tre tipi di tessuto muscolare Scheletrico (striato) : è attaccato alle ossa e muove parti dello scheletro; movimenti volontari e alcuni
La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico.
 La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico. N1 N2 Punto di discontinuità Elettriche Chimiche: Eccitatorie Inibitorie La sinapsi I neuroni comunicano
La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico. N1 N2 Punto di discontinuità Elettriche Chimiche: Eccitatorie Inibitorie La sinapsi I neuroni comunicano
Muscolo, un sistema a cilindri
 Muscolo, un sistema a cilindri Motilità La motilità è una caratteristica fondamentale della materia vivente Dagli organismi unicellulari a quelli più evoluti, la motilità si manifesta nelle forme più svariate
Muscolo, un sistema a cilindri Motilità La motilità è una caratteristica fondamentale della materia vivente Dagli organismi unicellulari a quelli più evoluti, la motilità si manifesta nelle forme più svariate
Muscolo Scheletrico. Anatomia funzionale, eccitazione, contrazione. FGE aa
 Muscolo Scheletrico Anatomia funzionale, eccitazione, contrazione FGE aa.2016-17 Obiettivi Anatomia funzionale del muscolo (cenni) Struttura della fibra muscolare; l unità funzionale del muscolo striato-scheletrico:
Muscolo Scheletrico Anatomia funzionale, eccitazione, contrazione FGE aa.2016-17 Obiettivi Anatomia funzionale del muscolo (cenni) Struttura della fibra muscolare; l unità funzionale del muscolo striato-scheletrico:
Muscolo scheletrico e cardiaco
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo scheletrico e cardiaco Nicotina: agonista dei recettori colinergici nicotinici (recettori ionotropi) presenti sulla
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo scheletrico e cardiaco Nicotina: agonista dei recettori colinergici nicotinici (recettori ionotropi) presenti sulla
David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis. Biologia.blu. C - Il corpo umano
 David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis Biologia.blu C - Il corpo umano 1 Il sistema nervoso 2 Il sistema nervoso (SN) Il SN coordina le attività delle diverse
David Sadava, H. Craig Heller, Gordon H. Orians, William K. Purves, David M. Hillis Biologia.blu C - Il corpo umano 1 Il sistema nervoso 2 Il sistema nervoso (SN) Il SN coordina le attività delle diverse
La pompa sodio-potassio
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
I sistemi muscolare e scheletrico
 I sistemi muscolare e scheletrico Il sistema muscolare Esistono tre tipi di muscoli con caratteristiche strutturali e funzionali diverse: o il muscolo striato o scheletrico; o il muscolo liscio; o il muscolo
I sistemi muscolare e scheletrico Il sistema muscolare Esistono tre tipi di muscoli con caratteristiche strutturali e funzionali diverse: o il muscolo striato o scheletrico; o il muscolo liscio; o il muscolo
L apparato muscolare: tessuto muscolare scheletrico. Cattedra di Fisiologia Umana
 L apparato muscolare: tessuto muscolare scheletrico Cattedra di Fisiologia Umana 1. Organizzazione dei muscoli scheletrici Un muscolo scheletrico è composto da fasci di fibre muscolari racchiusi dall epimisio.
L apparato muscolare: tessuto muscolare scheletrico Cattedra di Fisiologia Umana 1. Organizzazione dei muscoli scheletrici Un muscolo scheletrico è composto da fasci di fibre muscolari racchiusi dall epimisio.
Fisiologia cellulare e Laboratorio di Colture cellulari
 Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
SISTEMA NERVOSO SISTEMA NERVOSO CENTRALE (SNC) SISTEMA NERVOSO PERIFERICO (SNP) ENCEFALO: CERVELLO CERVELLETTO MIDOLLO ALLUNGATO (TRONCO ENCEFALICO)
 SISTEMA NERVOSO PERIFERICO (SNP) ENCEFALO: CERVELLO CERVELLETTO MIDOLLO ALLUNGATO (TRONCO ENCEFALICO)") IL SISTEMA NERVOSO SISTEMA NERVOSO SISTEMA NERVOSO CENTRALE (SNC) SISTEMA NERVOSO PERIFERICO (SNP) ENCEFALO: CERVELLO CERVELLETTO MIDOLLO ALLUNGATO (TRONCO ENCEFALICO) MIDOLLO SPINALE VOLONTARIO: NERVI
IL SISTEMA NERVOSO SISTEMA NERVOSO SISTEMA NERVOSO CENTRALE (SNC) SISTEMA NERVOSO PERIFERICO (SNP) ENCEFALO: CERVELLO CERVELLETTO MIDOLLO ALLUNGATO (TRONCO ENCEFALICO) MIDOLLO SPINALE VOLONTARIO: NERVI
Il potenziale di membrana a riposo
 Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA)
") Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Ioni fosfato e proteine (anioni) sono prevalenti nella cellula
 sono prevalenti nella cellula") Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
E presente nella parete di organi interni cavi (arterie, vene, canale digerente, vescica).
.") Il muscolo liscio E presente nella parete di organi interni cavi (arterie, vene, canale digerente, vescica). E formato da cellule piccole (lunghezza 20-600 µm, diametro 2-5 µm), con singolo nucleo, che
Il muscolo liscio E presente nella parete di organi interni cavi (arterie, vene, canale digerente, vescica). E formato da cellule piccole (lunghezza 20-600 µm, diametro 2-5 µm), con singolo nucleo, che
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA
 FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
Corso di Laurea Magistrale in Medicina e Chirurgia. Biofisica e Fisiologia I. Muscolo liscio
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
La funzione fondamentale del tessuto muscolare è la contrazione, durante la quale il muscolo sviluppa forza e si accorcia spostando un carico e
 Il Muscolo La funzione fondamentale del tessuto muscolare è la contrazione, durante la quale il muscolo sviluppa forza e si accorcia spostando un carico e producendo lavoro meccanico. I muscoli sono responsabili
Il Muscolo La funzione fondamentale del tessuto muscolare è la contrazione, durante la quale il muscolo sviluppa forza e si accorcia spostando un carico e producendo lavoro meccanico. I muscoli sono responsabili
SISTEMA NERVOSO! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani
 ! SISTEMA NERVOSO! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani Il Sistema Nervoso coordina le attività della vita di relazione e svolge le seguenti funzioni:! riceve stimoli ed elabora
! SISTEMA NERVOSO! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani Il Sistema Nervoso coordina le attività della vita di relazione e svolge le seguenti funzioni:! riceve stimoli ed elabora
Laura Condorelli 2014 Pagina 1. Figura 1 controllo del sistema nervoso autonomo sui muscoli lisci
 SISTEMA MUSCOLARE Esistono 2 tipi differenti di muscoli: - Muscoli lisci - Muscoli striati I muscoli lisci sono quelli viscerali, cioè quelli attaccati agli organi interni (come l apparato digerente e
SISTEMA MUSCOLARE Esistono 2 tipi differenti di muscoli: - Muscoli lisci - Muscoli striati I muscoli lisci sono quelli viscerali, cioè quelli attaccati agli organi interni (come l apparato digerente e
Le sinapsi elettriche
 Caratteristica peculiare delle cellule nervose è quella di condurre e comunicare informazioni. Le zone specializzate a livello delle quali le cellule entrano in comunicazione sono definite sinapsi La trasmissione
Caratteristica peculiare delle cellule nervose è quella di condurre e comunicare informazioni. Le zone specializzate a livello delle quali le cellule entrano in comunicazione sono definite sinapsi La trasmissione
TARGETS DEI FARMACI ATTUALMENTE IN USO
 TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
Anatomia microscopica e funzionale del muscolo scheletrico. La fibra del muscolo scheletrico E l unità istologica elementare dei muscoli.
 Domenica 11 Novembre 2012 CORSO DI FORMAZIONE TECNICO EDUCATORE REGIONALE LA MACCHINA DELL UOMO: CENNI DI FISIOLOGIA, ANATOMIA DELL APPARATO LOCOMOTORE LA CONTRAZIONE MUSCOLARE Relatore: Dott.Michele Bisogni
Domenica 11 Novembre 2012 CORSO DI FORMAZIONE TECNICO EDUCATORE REGIONALE LA MACCHINA DELL UOMO: CENNI DI FISIOLOGIA, ANATOMIA DELL APPARATO LOCOMOTORE LA CONTRAZIONE MUSCOLARE Relatore: Dott.Michele Bisogni
sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio in senso lato sinapsi neurone effettori elettriche chimiche
 sinapsi 1 sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio neurone in senso lato effettori cellule muscolari, gh. esocrine, gh. endocrine sinapsi elettriche chimiche 2 sinapsi
sinapsi 1 sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio neurone in senso lato effettori cellule muscolari, gh. esocrine, gh. endocrine sinapsi elettriche chimiche 2 sinapsi
Tessuto Nervoso. Parte b
 Tessuto Nervoso Parte b Sinapsi Siti dove gli impulsi nervosi passano da una cellula presinaptica (neurone) ad una postsinaptica (neurone, cellula muscolare o ghiandolare). Sinapsi Elettrica Solo retina
Tessuto Nervoso Parte b Sinapsi Siti dove gli impulsi nervosi passano da una cellula presinaptica (neurone) ad una postsinaptica (neurone, cellula muscolare o ghiandolare). Sinapsi Elettrica Solo retina
La conduzione dell impulso nervoso
 La conduzione dell impulso nervoso La recezione sensoriale Soglia di percezione degli stimoli sensoriali Vista: percezione della luce di una candela a 48 km di distanza, in una notte serena e limpida Udito:
La conduzione dell impulso nervoso La recezione sensoriale Soglia di percezione degli stimoli sensoriali Vista: percezione della luce di una candela a 48 km di distanza, in una notte serena e limpida Udito:
Il sistema nervoso (SN)
") Il sistema nervoso (SN) Il SN coordina le attività delle diverse parti del corpo e permette di entrare in relazione con l ambiente esterno, la gestione di tutte le informazioni è centralizzata nell encefalo.
Il sistema nervoso (SN) Il SN coordina le attività delle diverse parti del corpo e permette di entrare in relazione con l ambiente esterno, la gestione di tutte le informazioni è centralizzata nell encefalo.
IL TESSUTO MUSCOLARE MUSCOLO LISCIO, STRIATO E CARDIACO
 IL TESSUTO MUSCOLARE MUSCOLO LISCIO, STRIATO E CARDIACO Il muscolo striato presenta cellule polinucleate (con nuclei periferici) con caratteristiche bande chiare e scure MUSCOLO VOLONTARIO, MUSCOLO SCHELETRICO
IL TESSUTO MUSCOLARE MUSCOLO LISCIO, STRIATO E CARDIACO Il muscolo striato presenta cellule polinucleate (con nuclei periferici) con caratteristiche bande chiare e scure MUSCOLO VOLONTARIO, MUSCOLO SCHELETRICO
muscolo scheletrico i tessuti muscolari sono costituiti da cellule eccitabili
 muscolo scheletrico muscolo scheletrico (striato volontario, attivato dai motoneruroni α) muscolo cardiaco (striato involontario) muscolo liscio (involontario) i tessuti muscolari sono costituiti da cellule
muscolo scheletrico muscolo scheletrico (striato volontario, attivato dai motoneruroni α) muscolo cardiaco (striato involontario) muscolo liscio (involontario) i tessuti muscolari sono costituiti da cellule
Aspetti meccanici della contrazione muscolare
 Aspetti meccanici della contrazione muscolare ARGOMENTI Innervazione del muscolo scheletrico Placca neuro muscolare Unità neuro motorie Tipi di contrazione Scosse semplici Relazioni tensione-lunghezza
Aspetti meccanici della contrazione muscolare ARGOMENTI Innervazione del muscolo scheletrico Placca neuro muscolare Unità neuro motorie Tipi di contrazione Scosse semplici Relazioni tensione-lunghezza
Il sistema nervoso autonomo: simpatico e parasimpatico
 Il sistema nervoso autonomo: simpatico e parasimpatico Proprietà dell omeostasi: 1- mantenimento della costanza dell ambiente interno 2- aumento o diminuzione del controllo tonico 3- controllo antagonista
Il sistema nervoso autonomo: simpatico e parasimpatico Proprietà dell omeostasi: 1- mantenimento della costanza dell ambiente interno 2- aumento o diminuzione del controllo tonico 3- controllo antagonista
Lezioni di Fisiologia
 Corso di Laurea Infermieristica N AA 2011/2012 Lezioni di Fisiologia SISTEMA NERVOSO E ORGANI DI SENSO Prof. A. Pulcini Dipartimento di Scienze Chirurgiche Policlinico Umberto I Sapienza Roma Il Sistema
Corso di Laurea Infermieristica N AA 2011/2012 Lezioni di Fisiologia SISTEMA NERVOSO E ORGANI DI SENSO Prof. A. Pulcini Dipartimento di Scienze Chirurgiche Policlinico Umberto I Sapienza Roma Il Sistema
SINAPSI. avviene senza l intervento. di mediatori chimici.
 SINAPSI Specializzazioni strutturali tipiche che consentono la trasmissione unidirezionale di impulsi elettrici da un neurone ad altri neuroni od a cellule effettrici periferiche. Sinapsi elettriche :
SINAPSI Specializzazioni strutturali tipiche che consentono la trasmissione unidirezionale di impulsi elettrici da un neurone ad altri neuroni od a cellule effettrici periferiche. Sinapsi elettriche :
Funzione Insieme al tessuto osseo è responsabile della locomozione e del movimento delle varie parti del corpo. Origine Deriva dal mesoderma
 TESSUTO MUSCOLARE www.slidetube.it Funzione Insieme al tessuto osseo è responsabile della locomozione e del movimento delle varie parti del corpo. Origine Deriva dal mesoderma Caratteristiche E costituito
TESSUTO MUSCOLARE www.slidetube.it Funzione Insieme al tessuto osseo è responsabile della locomozione e del movimento delle varie parti del corpo. Origine Deriva dal mesoderma Caratteristiche E costituito
Elettricità cellulare
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Elettricità cellulare 30/3/2006 Sistemi biologici Essenzialmente costituiti da acqua (solvente) e da differenti soluti (molti dei
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Elettricità cellulare 30/3/2006 Sistemi biologici Essenzialmente costituiti da acqua (solvente) e da differenti soluti (molti dei
PROF. ALESSANDRO MALFATTI. Corso di Fisiologia e Benessere degli animali in produzione. STRUTTURA e FUNZIONE
 STRUTTURA e FUNZIONE Ogni neurone presenta 4 zone: DENDRITI: ricezione dell'impulso proveniente da altri neuroni SOMA (PIRENOFORO): integrazione delle numerose afferenze, spesso contrastanti sintesi del
STRUTTURA e FUNZIONE Ogni neurone presenta 4 zone: DENDRITI: ricezione dell'impulso proveniente da altri neuroni SOMA (PIRENOFORO): integrazione delle numerose afferenze, spesso contrastanti sintesi del
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.
 Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
Accoppiamento eccitazione-contrazione
 Accoppiamento eccitazione-contrazione Potenziale d'azione motoneurone Potenziale d'azione muscolo Contrazione muscolo Ricordate cosa c è alla giunzione muscolare? Anni 30 40: Heilbrun ipotizza un ruolo
Accoppiamento eccitazione-contrazione Potenziale d'azione motoneurone Potenziale d'azione muscolo Contrazione muscolo Ricordate cosa c è alla giunzione muscolare? Anni 30 40: Heilbrun ipotizza un ruolo
Stimolazione artificiale del cuore
 Stimolazione artificiale del cuore Il ciclo del cuore: Impulso elettrico di stimolazione da parte di un gruppo di fibre nervose il nodo sinoatriale, Stimolatori artificiali Pacemaker impiantati chirurgicamente
Stimolazione artificiale del cuore Il ciclo del cuore: Impulso elettrico di stimolazione da parte di un gruppo di fibre nervose il nodo sinoatriale, Stimolatori artificiali Pacemaker impiantati chirurgicamente
Classificazione del tessuto muscolare. 1. Posizione rispetto ad altre strutture corporee 2. Composizione istologica 3. Modalità di controllo
 Classificazione del tessuto muscolare 1. Posizione rispetto ad altre strutture corporee 2. Composizione istologica 3. Modalità di controllo controllo anatomia istologia Volontario Involontario scheletrico
Classificazione del tessuto muscolare 1. Posizione rispetto ad altre strutture corporee 2. Composizione istologica 3. Modalità di controllo controllo anatomia istologia Volontario Involontario scheletrico
POTENZIALE DI MEMBRANA
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Scaricato da 1
 Muscolo cardiaco scheletrico liscio Eterogeneità: muscolo liscio vascolare gastrointestinale urinario respiratorio riproduttivo oculare Anatomia: le fibre contrattili del muscolo liscio sono disposte in
Muscolo cardiaco scheletrico liscio Eterogeneità: muscolo liscio vascolare gastrointestinale urinario respiratorio riproduttivo oculare Anatomia: le fibre contrattili del muscolo liscio sono disposte in
I TRE TIPI DI CELLULE MUSCOLARI MICROSCOPIO OTTICO: SEZIONE LONGITUDINALE DI FIBRE MUSCOLARI STRIATE
 I TRE TIPI DI CELLULE MUSCOLARI MICROSCOPIO OTTICO: SEZIONE LONGITUDINALE DI FIBRE MUSCOLARI STRIATE 1 2 3 SEZIONE TRASVERSALE DI UN MUSCOLO SCHELETRICO: IN NERO LE FIBRE MUSCOLARI, I SETTI CONNETTIVALI
I TRE TIPI DI CELLULE MUSCOLARI MICROSCOPIO OTTICO: SEZIONE LONGITUDINALE DI FIBRE MUSCOLARI STRIATE 1 2 3 SEZIONE TRASVERSALE DI UN MUSCOLO SCHELETRICO: IN NERO LE FIBRE MUSCOLARI, I SETTI CONNETTIVALI
Il sistema muscolare
 Il sistema muscolare Tipi di muscoli Scheletrici (volontari e striati) Cardiaco (involontario ma striato) Lisci (involontari) I principali muscoli scheletrici Le proprietà del muscolo Le proprietà specifiche
Il sistema muscolare Tipi di muscoli Scheletrici (volontari e striati) Cardiaco (involontario ma striato) Lisci (involontari) I principali muscoli scheletrici Le proprietà del muscolo Le proprietà specifiche
Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA
 Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA Fisiologia cellulare: Membrana cellulare. Trasporto di ioni e molecole attraverso la membrana cellulare. Canali ionici, pompe e proteine vettrici.
Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA Fisiologia cellulare: Membrana cellulare. Trasporto di ioni e molecole attraverso la membrana cellulare. Canali ionici, pompe e proteine vettrici.
Il Tessuto nervoso. E costituito da cellule (chiamate. neuroni) specializzate nella. generazione e nella conduzione di. particolari segnali.
 specializzate nella. generazione e nella conduzione di. particolari segnali.") Il Tessuto nervoso E costituito da cellule (chiamate neuroni) specializzate nella generazione e nella conduzione di particolari segnali. Il Tessuto nervoso Funzione - integrazione tra i diversi organi
Il Tessuto nervoso E costituito da cellule (chiamate neuroni) specializzate nella generazione e nella conduzione di particolari segnali. Il Tessuto nervoso Funzione - integrazione tra i diversi organi
V = 130 mv R = Ω = = A I = V R. Trascuriamo il canale del Cl - Voltaggio totale V. Resistenza totale R
 LA MEMBRANA I canali passivi trans-membranici selettivi per ciascuna specie di ione possono essere schematizzati come una combinazione di resistenze e batterie Trascuriamo il canale del Cl - V = 130 mv
LA MEMBRANA I canali passivi trans-membranici selettivi per ciascuna specie di ione possono essere schematizzati come una combinazione di resistenze e batterie Trascuriamo il canale del Cl - V = 130 mv
Fisiologia. ISTRUTTORI ARTI MARZIALI Roma, 8 Maggio 2016
 ISTRUTTORI ARTI MARZIALI Roma, 8 Maggio 2016 Fisiologia Ivan Cirami Istituto di Medicina e Scienza dello Sport C.O.N.I. - Roma Resp. Regionale Pallavolo Csain Lazio ASD Fisio&Sport - Decimo Roma Pallavolo
ISTRUTTORI ARTI MARZIALI Roma, 8 Maggio 2016 Fisiologia Ivan Cirami Istituto di Medicina e Scienza dello Sport C.O.N.I. - Roma Resp. Regionale Pallavolo Csain Lazio ASD Fisio&Sport - Decimo Roma Pallavolo
3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione
 3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Genesi e mantenimento
3. Fisiologia Cellulare Potenziale transmembranario, potenziale d azione Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Genesi e mantenimento
Testi consigliati. Fondamenti di Anatomia e Fisiologia di F. H. Martini Casa editrice: EdiSES. Modalità di valutazione dell'apprendimento
 Anno Accademico 2011-2012 Infermieristica: Modulo di Fisiologia Umana (2 CFU) Fisioterapia: Modulo di Fisiologia (3 CFU) Testi consigliati Fondamenti di Anatomia e Fisiologia di F. H. Martini Casa editrice:
Anno Accademico 2011-2012 Infermieristica: Modulo di Fisiologia Umana (2 CFU) Fisioterapia: Modulo di Fisiologia (3 CFU) Testi consigliati Fondamenti di Anatomia e Fisiologia di F. H. Martini Casa editrice:
le sinapsi Prof. Davide Cervia - Fisiologia Fisiologia della cellula: trasmissione sinaptica
 le sinapsi 1 2 sinapsi elettriche 3 presenti nel tessuto muscolare liscio e cardiaco, ed in alcuni tessuti neuroendocrini trovate anche nel SNC di invertebrati e di mammiferi tra neuroni ma anche tra cellule
le sinapsi 1 2 sinapsi elettriche 3 presenti nel tessuto muscolare liscio e cardiaco, ed in alcuni tessuti neuroendocrini trovate anche nel SNC di invertebrati e di mammiferi tra neuroni ma anche tra cellule
Tessuto muscolare. La possibilità di compiere movimenti e di mantenere la stazione eretta è affidata alla capacità contrattile della cellula muscolare
 Tessuto muscolare La possibilità di compiere movimenti e di mantenere la stazione eretta è affidata alla capacità contrattile della cellula muscolare Le proprietà fondamentali delle cellule muscolari sono:
Tessuto muscolare La possibilità di compiere movimenti e di mantenere la stazione eretta è affidata alla capacità contrattile della cellula muscolare Le proprietà fondamentali delle cellule muscolari sono:
otenziali graduati (elettrotonici) potenziale d azione
 potenziale d azione") I segnali elettrici che si generano nei neuroni: potenziali graduati (elettrotonici) potenziale d azione sono modificazioni del potenziale di riposo determinate dall apertura o chiusura di canali ionici.
I segnali elettrici che si generano nei neuroni: potenziali graduati (elettrotonici) potenziale d azione sono modificazioni del potenziale di riposo determinate dall apertura o chiusura di canali ionici.
Meccanismo d azione dei NT nelle sinapsi dirette. 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie
 Meccanismo d azione dei NT nelle sinapsi dirette 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie 1) La giunzione neuromuscolare La prima sinapsi ad essere studiata nel dettaglio
Meccanismo d azione dei NT nelle sinapsi dirette 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie 1) La giunzione neuromuscolare La prima sinapsi ad essere studiata nel dettaglio
Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A
 Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A. 2015-2016 Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente Riflessi nervosi: si attuano
Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A. 2015-2016 Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente Riflessi nervosi: si attuano
Proprietà di permeabilità della membrana
 Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
SISTEMA MUSCOLARE! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani
 ! SISTEMA MUSCOLARE! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani ! SISTEMA MUSCOLARE! Nell uomo il movimento si realizza grazie a:! Lo scheletro o apparato scheletrico! Il Sistema muscolare!
! SISTEMA MUSCOLARE! Prof.ssa A. Biolchini Prof.ssa S. Di Giulio Prof. M. Montani ! SISTEMA MUSCOLARE! Nell uomo il movimento si realizza grazie a:! Lo scheletro o apparato scheletrico! Il Sistema muscolare!
Il potenziale di membrana di una cellula che non conduce impulsi si definisce potenziale di membrana a riposo ( PRM ).
.") POTENZIALI DI MEMBRANA Tutte le cellule viventi possiedono e mantengono una concentrazione di ioni diversa nell interno della cellula e nei liquidi extracellulari. - I liquidi extracellulari hanno un elevata
POTENZIALI DI MEMBRANA Tutte le cellule viventi possiedono e mantengono una concentrazione di ioni diversa nell interno della cellula e nei liquidi extracellulari. - I liquidi extracellulari hanno un elevata
I riflessi motorio maniera stereotipata indipendentemente
 I riflessi Automatismo motorio, mediato dal SNC in risposta a un certo stimolo che si ripete in maniera stereotipata ogni volta che viene evocato e si attua indipendentemente dalla coscienza del soggetto.
I riflessi Automatismo motorio, mediato dal SNC in risposta a un certo stimolo che si ripete in maniera stereotipata ogni volta che viene evocato e si attua indipendentemente dalla coscienza del soggetto.
Anatomia e Fisiologia della giunzione neuromuscolare
 Anatomia e Fisiologia della giunzione neuromuscolare La giunzione neuromuscolare La placca neuromuscolare è il contatto tra il terminale della fibra nervosa e la fibra muscolare. Normalmente per ogni fibra
Anatomia e Fisiologia della giunzione neuromuscolare La giunzione neuromuscolare La placca neuromuscolare è il contatto tra il terminale della fibra nervosa e la fibra muscolare. Normalmente per ogni fibra
Cellule nervose e comportamento. Caratteristica importante dell organizzazione cerebrale=
 Cellule nervose e comportamento Caratteristica importante dell organizzazione cerebrale= cellule con proprietà sostanzialmente simili possono svolgere funzioni assai diverse a seconda del tipo di connessioni
Cellule nervose e comportamento Caratteristica importante dell organizzazione cerebrale= cellule con proprietà sostanzialmente simili possono svolgere funzioni assai diverse a seconda del tipo di connessioni
Che ruolo ha la membrana plasmatica nei potenziali di diffusione?
 Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
Le membrane cellulari
 Le membrane cellulari La struttura delle membrane biologiche Le membrane cellulari sono formate da un doppio strato fosfolipidico dove si trovano varie proteine, che possono essere: periferiche: si trovano
Le membrane cellulari La struttura delle membrane biologiche Le membrane cellulari sono formate da un doppio strato fosfolipidico dove si trovano varie proteine, che possono essere: periferiche: si trovano
Permeabilità Di Membrana Cellulare
 Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Contrazione muscolare
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Contrazione muscolare I muscoli hanno due funzioni: generare movimento generare forza I muscoli scheletrici generano anche calore
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Contrazione muscolare I muscoli hanno due funzioni: generare movimento generare forza I muscoli scheletrici generano anche calore
Differenze tra SINAPSI CENTRALI E GIUNZIONI NEUROMUSCOLARI
 Differenze tra SINAPSI CENTRALI E GIUNZIONI NEUROMUSCOLARI 1. Ogni fibrocellula muscolare riceve segnali da un singolo neurone (motoneurone) 2. I potenziali postsinaptici (potenziali di placca) sono sempre
Differenze tra SINAPSI CENTRALI E GIUNZIONI NEUROMUSCOLARI 1. Ogni fibrocellula muscolare riceve segnali da un singolo neurone (motoneurone) 2. I potenziali postsinaptici (potenziali di placca) sono sempre
Il Sistema Nervoso coordina le attività della vita di relazione e svolge le
 Il Sistema Nervoso coordina le attività della vita di relazione e svolge le seguenti funzioni: riceve stimoli ed elabora risposte; memorizza informazioni; elabora ragionamenti. L unità fondamentale del
Il Sistema Nervoso coordina le attività della vita di relazione e svolge le seguenti funzioni: riceve stimoli ed elabora risposte; memorizza informazioni; elabora ragionamenti. L unità fondamentale del
APPUNTI DI FISIOLOGIA DEL SISTEMA NERVOSO. Modello di NEURONE. Definizione di NEURONE: Unità strutturale e funzionale del Sistema nervoso.
 APPUNTI DI FISIOLOGIA DEL SISTEMA NERVOSO Modello di NEURONE Definizione di NEURONE: Unità strutturale e funzionale del Sistema nervoso. Stato elettrochimico a riposo L'analisi chimica evidenzia una distribuzione
APPUNTI DI FISIOLOGIA DEL SISTEMA NERVOSO Modello di NEURONE Definizione di NEURONE: Unità strutturale e funzionale del Sistema nervoso. Stato elettrochimico a riposo L'analisi chimica evidenzia una distribuzione
Citoscheletro, matrice extracellulare e giunzioni cellulari
 Citoscheletro, matrice extracellulare e giunzioni cellulari Le distrofie muscolari Malattie come la distrofia muscolare di Duchenne determinano la mancanza di proteine (come la distrofina) coinvolte nei
Citoscheletro, matrice extracellulare e giunzioni cellulari Le distrofie muscolari Malattie come la distrofia muscolare di Duchenne determinano la mancanza di proteine (come la distrofina) coinvolte nei
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE
 COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
IL SISTEMA NERVOSO. Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO. Sensoriale (dall'esterno)
 Trasmissivo (Cablaggio) SISTEMA NERVOSO. Sensoriale (dall'esterno)") IL SISTEMA NERVOSO Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO Periferico Afferente (dalla periferia verso l'unità centrale) Sensoriale (dall'esterno) Autonomo
IL SISTEMA NERVOSO Centrale (Unità di elaborazione e controllo) Trasmissivo (Cablaggio) SISTEMA NERVOSO Periferico Afferente (dalla periferia verso l'unità centrale) Sensoriale (dall'esterno) Autonomo
COMUNICAZIONE INTERCELLULARE NEL SISTEMA NERVOSO
 COMUNICAZIONE INTERCELLULARE NEL SISTEMA NERVOSO Si verifica attraverso sinapsi, zone di contatto fra un neurone e la cellula bersaglio (un neurone o una cellula non neuronale) Le sinapsi, in base al tipo
COMUNICAZIONE INTERCELLULARE NEL SISTEMA NERVOSO Si verifica attraverso sinapsi, zone di contatto fra un neurone e la cellula bersaglio (un neurone o una cellula non neuronale) Le sinapsi, in base al tipo
5. Trasporto di membrana
 5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
Vena cava superiore) Arteria polmonare valvole semilunari Arterie polmonari sinistre. Arterie polmonari destre) Vena cava inferiore) Ventricolo destro
 Arteria polmonare valvole semilunari Arterie polmonari sinistre. Arterie polmonari destre) Vena cava inferiore) Ventricolo destro") Fisiologia cardiaca Arterie polmonari destre) Vena cava superiore) Arteria polmonare valvole semilunari Arterie polmonari sinistre Atrio destro Vene polmonari sinistre Valvola bicuspide (mitrale) Valvola
Fisiologia cardiaca Arterie polmonari destre) Vena cava superiore) Arteria polmonare valvole semilunari Arterie polmonari sinistre Atrio destro Vene polmonari sinistre Valvola bicuspide (mitrale) Valvola
Formazione di una fibra muscolare scheletrica
 Formazione di una fibra muscolare scheletrica I mioblasti derivano da cellule del midollo osseo. Queste in determinate condizioni di necessità (stress chimico, traumatico etc.) vengono mobilizzate dalla
Formazione di una fibra muscolare scheletrica I mioblasti derivano da cellule del midollo osseo. Queste in determinate condizioni di necessità (stress chimico, traumatico etc.) vengono mobilizzate dalla
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Il funzionamento della cellula Le cellule sono veri e propri esseri viventi: sono capaci di alimentarsi, di crescere, di produrre nuove sostanze, di riprodursi, di espellere rifiuti
METABOLISMO CELLULARE Il funzionamento della cellula Le cellule sono veri e propri esseri viventi: sono capaci di alimentarsi, di crescere, di produrre nuove sostanze, di riprodursi, di espellere rifiuti
Tutte le cellule sono capaci di generare potenziali elettrici a riposo a cavallo delle membrane cellulari e sono in grado di cambiare la loro
 Tutte le cellule sono capaci di generare potenziali elettrici a riposo a cavallo delle membrane cellulari e sono in grado di cambiare la loro permeabilità ai vari ioni. Ci sono però alcune cellule, dette
Tutte le cellule sono capaci di generare potenziali elettrici a riposo a cavallo delle membrane cellulari e sono in grado di cambiare la loro permeabilità ai vari ioni. Ci sono però alcune cellule, dette
TESSUTO MUSCOLARE. MUSCOLO LISCIO TONACA MUSCOLARE dei visceri e dei vasi (controllato dal SNA, indipendente dalla volontà)
") 1 TESSUTO MUSCOLARE È dotato di contrattilità La contrattilità è la capacità di accorciamento La contrattilità si origina dallo scorrimento di proteine con consumo di ATP MUSCOLO STRIATO SCHELETRICO (controllato
1 TESSUTO MUSCOLARE È dotato di contrattilità La contrattilità è la capacità di accorciamento La contrattilità si origina dallo scorrimento di proteine con consumo di ATP MUSCOLO STRIATO SCHELETRICO (controllato
Il trasferimento di segnali tra cellule eccitabili avviene a livello delle sinapsi. La trasmissione sinaptica può essere:
 La trasmissione sinaptica Il trasferimento di segnali tra cellule eccitabili avviene a livello delle sinapsi La trasmissione sinaptica può essere: Elettrica Chimica Sinapsi elettriche Continuità citoplasmatica
La trasmissione sinaptica Il trasferimento di segnali tra cellule eccitabili avviene a livello delle sinapsi La trasmissione sinaptica può essere: Elettrica Chimica Sinapsi elettriche Continuità citoplasmatica
Contrazione muscolare
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Contrazione muscolare I muscoli hanno due funzioni: generare movimento generare forza I muscoli scheletrici generano anche calore
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Contrazione muscolare I muscoli hanno due funzioni: generare movimento generare forza I muscoli scheletrici generano anche calore
La Forza muscolare. University of Rome "Tor Vergata" - Faculty of Medicine and Surgery - School of Sport and Exercise Science
 La Forza muscolare University of Rome "Tor Vergata" - Faculty of Medicine and Surgery - School of Sport and Exercise Science Qualità fisiche di base e derivate Forza max Forza reattiva Forza dinamica massima
La Forza muscolare University of Rome "Tor Vergata" - Faculty of Medicine and Surgery - School of Sport and Exercise Science Qualità fisiche di base e derivate Forza max Forza reattiva Forza dinamica massima
Funzione dei neuroni l impulso nervoso
 Funzione dei neuroni l impulso nervoso L IMPULSO NERVOSO è un segnale elettrico La superficie interna della membrana plasmatica del neurone è leggermente più negativa rispetto a quella esterna per una
Funzione dei neuroni l impulso nervoso L IMPULSO NERVOSO è un segnale elettrico La superficie interna della membrana plasmatica del neurone è leggermente più negativa rispetto a quella esterna per una
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni
 5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
LE SINAPSI. Assone. PdA e trasporto del neurotrasmettitore. Sinapsi assodendritica Recettore
 LE SINAPSI Al fine di rendere più efficiente la comunicazione chimica di tipo paracrina l'evoluzione ha "prodotto" le sinapsi, costituite da due microaree di membrana, appartenenti rispettivamente a due
LE SINAPSI Al fine di rendere più efficiente la comunicazione chimica di tipo paracrina l'evoluzione ha "prodotto" le sinapsi, costituite da due microaree di membrana, appartenenti rispettivamente a due
POTENZIALE DI MEMBRANA
 POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
Cambiamenti durante l esercizio: a mantenere l equilibrio delle funzioni del corpo: OMEOSTASI
 Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della frequenza respiratoria a Aumento della frequenza cardiaca Aumento
Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della frequenza respiratoria a Aumento della frequenza cardiaca Aumento
SISTEMA NEUROENDOCRINO
 Mantenimento dell omeostasi Nervoso: risposta rapida +! Endocrino: risposta lenta SISTEMA NEUROENDOCRINO Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato
Mantenimento dell omeostasi Nervoso: risposta rapida +! Endocrino: risposta lenta SISTEMA NEUROENDOCRINO Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato