La struttura a doppio strato lipidico delle membrane biologiche condiziona fortemente il movimento di gas, ioni e molecole attraverso le membrane.
|
|
|
- Marisa Rota
- 7 anni fa
- Visualizzazioni
Transcript

1 1

2 La struttura a doppio strato lipidico delle membrane biologiche condiziona fortemente il movimento di gas, ioni e molecole attraverso le membrane. L interno idrofobico del doppio strato lipidico rende le membrane impermeabili alla maggior parte degli ioni e delle molecole cariche. In generale, mentre i gas come O2, CO2, NO e le molecole lipofiliche permeano facilmente attraverso le membrane, gli ioni e le molecole idrosolubili permeano con molta difficoltà o non permeano affatto attraverso la fase lipidica della membrana. Tuttavia, i movimenti di sostanze cariche sono necessari per il corretto funzionamento cellulare e pertanto tutte le cellule hanno sviluppato e perfezionato adeguati sistemi molecolari per facilitarne il movimento. Anche grosse proteine e particelle possono permeare la membrana cellulare attraverso specifici sistemi di trasporto. 2
3 I sistemi di trasporto attraverso la membrana possono essere schematizzati come segue: È possibile differenziarli in base al fatto che avvengano secondo gradiente di concentrazione chimica o elettrochimica di una sostanza in cui non vi è dispendio di energia metabolica della cellula, ed in questo caso si parla di trasporto passivo, che viceversa avvengano contro gradiente elettro-chimico e di conseguenza necessitino di energia metabolica, in questo caso si parla di trasporto attivo. Nell ambito del trasporto passivo, ovvero secondo gradiente di concentrazione si possono a loro volta distinguere processi di trasporto che avvengono per diffusione semplice, cioè senza l ausilio di alcuna proteina trasportatrice, da processi di diffusione facilitata, ancora una volta secondo gradiente elettrochimico, che però richiedono l intervento di vettore, ovvero una specifica proteina di trasporto o di un canale. Nel trasporto attivo che come detto avviene contro gradiente di concentrazione, si possono distinguere 2 modalità: nella prima, denominata trasporto attivo primario, la proteina di membrana utilizza direttamente l energia metabolica fornita dall idrolisi del ATP per creare ad esempio gradienti ionici di Na e K tra l ambiente intra ed extracellulare. Nel secondo caso, ovvero nel trasporto attivo secondario, il trasporto di una sostanza (ione o molecola) contro il proprio gradiente di concentrazione è accoppiato al movimento generalmente di uno ione secondo il proprio gradiente elettrochimico, dipendente a sua volta dall attività di un trasporto attivo primario. Al trasporto attivo appartengono i fenomeni di endo ed esocitosi (internalizzazione o espulsione di proteine ed altre macromolecole o fluidi mediante invaginazione o estroflessione della membrana plasmatica) nonché i processi di transcitosi (attraversamento di un epitelio). 3

4 Tutte le molecole o particelle in soluzione possiedono una energia termica: esse si muovono casualmente, urtano tra loro e cambiano continuamente direzione. Se in un determinato volume le molecole sono più concentrate queste diffonderanno verso zone a più bassa concentrazione fino a che non si distribuiranno uniformemente in ogni parte dello spazio disponibile. Questo processo è noto con il termine di diffusione. La diffusione è un processo passivo estremamente lento a livello macroscopico (tissutale) mentre è rapido a livello microscopico (cellulare). 4

5 La diffusione ha alcune proprietà: È un processo passivo che non richiede fonti di energia. Usa solo l energia termica (cinetica) del movimento molecolare. 1. le molecole diffondono da una zona a maggiore concentrazione verso zone a più bassa concentrazione. Più grande è la differenza di concentrazione e maggiore è la velocità delle molecole che diffondono per unità di tempo e per area di membrana. 2. la diffusione delle molecole procede fino a quando la concentrazione è uguale in entrambi i lati della membrana. Si raggiunge quindi un equilibrio dinamico e il movimento delle molecole continua senza spostamenti netti. 3. La diffusione è rapida a breve distanza e molto lenta su scala macroscopica. 4. la diffusione è direttamente proporzionale alla temperatura. Ad alte temperature il movimento molecolare è più rapido e la diffusione più veloce. 5. la diffusione è inversamente proporzionale alla dimensione molecolare. 6. La diffusione può verificarsi in un sistema aperto o attraverso una barriera che separa due sistemi. La diffusione attraverso le membrane è un po più complicata rispetto alla diffusione in un sistema aperto. Chiaramente solo le sostanze che possono attraversare il centro lipidido di una membrana possono muoversi per diffusione. 5

6 6

7 7

8 8
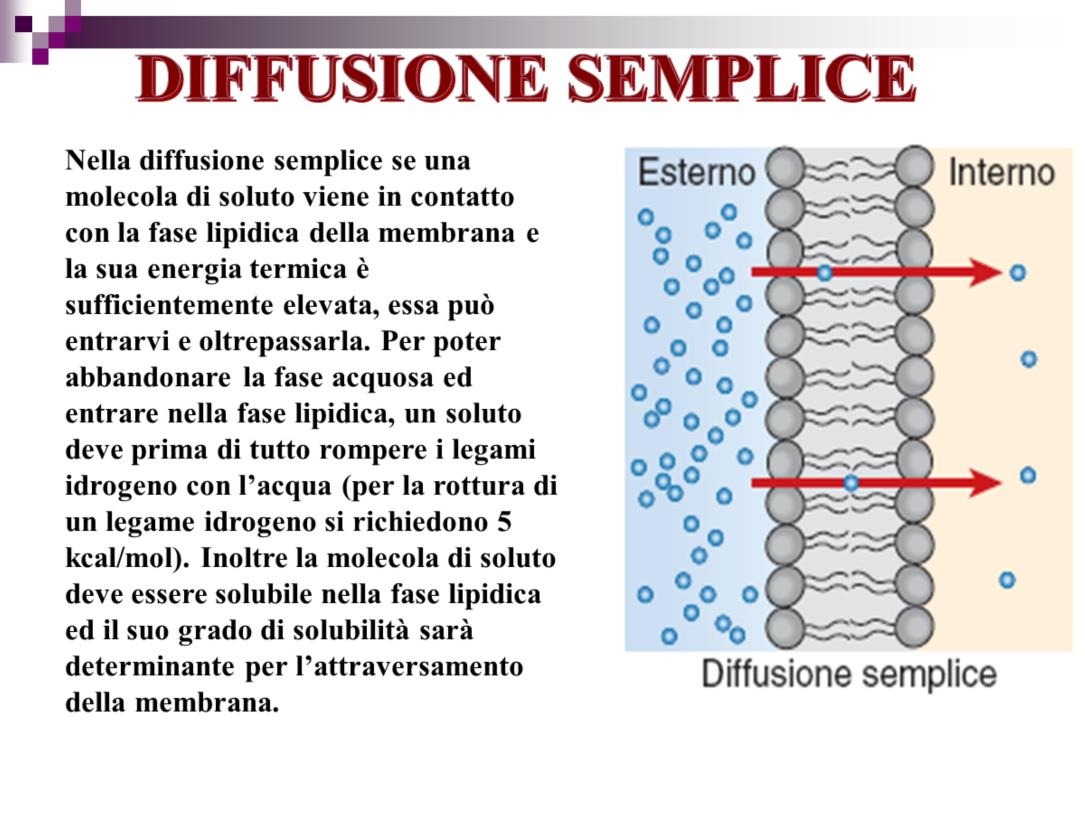
9 9

10 Se una membrana cellulare è permeabile a una molecola, tale molecola può entrare o uscire dalla cellula per diffusione. È necessario però che tali molecole siano in grado di passare attraverso il centro lipidico di una membrana cellulare. La diffusione attraverso attraverso il doppio strato lipidico della membrana va sotto il nome di diffusione semplice e possiede, oltre a tutte le proprietà elencate prima anche quelle seguenti: 1. la velocità di diffusione dipende dalla capacità della molecola che diffonde di sciogliersi nello strato lipidico della membrana. E cioè di quanto la membrana sia permeabile alle molecole che diffondono. Di regola solo i lipidi, gli steroidi e le piccole molecole lipofile possono attraversare le membrane per diffusione semplice. Ci sono però delle eccezioni che poi vedremo. 2. il tasso di diffusione attraverso una membrana è direttamente proporzionale all area della superficie della membrana. Quanto più grande è l area della superficie della membrana tanto più molecole possono diffondere attraverso di essa nell unità di tempo. È un concetto che può sembrare ovvio ma che ha profonde implicazioni fisiologiche. Per es. nella malattia polmonare denominata enfisema il tessuto polmonare collassa e viene distrutto questo determina una drastica riduzione dell area della superficie di scambio per la diffusione dell ossigeno e di conseguenza meno ossigeno arriva ai tessuti. 3. il tasso di diffusione attraverso una membrana è inversamente proporzionale allo spessore della membrana. Quanto più la membrana è spessa tanto più lenta e la velocità alla quale la diffusione ha luogo. 10

11 La permeabilità della molecola influenza chiaramente il flusso di un soluto attraverso la membrana. Maggiore è il valore di P, maggiore è il flusso di soluto tra i due compartimenti 11
 quando sono note le concentrazioni iniziali nei due compartimenti (C1 e C2), e la permeabilità di una molecola attraverso la")
12 Le regole della diffusione semplice attraverso le membrane possono essere espresse matematicamente da una equazione nota come Legge di Fick sulla diffusione che permette di calcolare la velocità di diffusione J (flusso) quando sono note le concentrazioni iniziali nei due compartimenti (C1 e C2), e la permeabilità di una molecola attraverso la membrana. 12

13 La legge di Fick suggerisce che nel caso di una diffusione condizionata esclusivamente da processi termodinamici c è una stretta linearità tra il gradiente di concentrazione del soluto e la velocità di diffusione. 13
. Inoltre la molecola di soluto deve essere solubile nella fase lipidica ed il suo grado di solubilità sarà determinante per l attraversamento della membrana.")
14 Per poter abbandonare la fase acquosa ed entrare nella fase lipidica, un soluto deve prima di tutto rompere i legami idrogeno con l acqua (per la rottura di un legame idrogeno si richiedono 5 kcal/mol). Inoltre la molecola di soluto deve essere solubile nella fase lipidica ed il suo grado di solubilità sarà determinante per l attraversamento della membrana. (la concentrazione in membrana aumenta all aumentare del coefficiente di partizione olio/acqua, K) 14

15 Quindi tutte le sostanze che formano pochi legami idrogeno con l acqua possono entrare più facilmente nel doppio strato lipidico mentre gli ioni inorganici non avranno quasi mai la possibilità di farlo. In genere molecole piccole e non polari come Ossigeno (32 dalton) e anidride carbonica (44 dalton) e molecole fortemente idrofobiche (o lipofiliche) diffondono rapidamente attraverso i doppi strati lipidici. Piccole molecole polari come acqua (18 dalton), etanolo (46 dalton) e urea (60 dalton) attraversano rapidamente; il glicerolo (92 dalton) diffonde meno rapidamente mentre il glucosio, che è una grossa molecola polare, praticamente non diffonde. I doppi strati lipidici sono praticamente impermeabili a molecole cariche e a ioni per quanto piccole esse siano; la carica e l alto grado di idratazione di queste molecole ne impedisce l ingresso nella fase idrocarburica del doppio strato. 15

16 Il grado di diffusione di una molecola attraverso una membrana, ovvero la sua permeabilità relativa, dipenderà dal suo coefficiente di ripartizione olio/acqua (K). Salvo poche eccezione, il grafico dimostra una correlazione quasi lineare tra permeabilità e coefficiente di ripartizione olio/acqua, indipendentemente dal peso molecolare. La permeabilità dell acqua attraverso le membrane cellulari è molto maggiore rispetto a quanto si possa dedurre dal suo coefficiente di ripartizione. Ciò è dovuto al fatto che l acqua può passare attraverso canali selettivi che penetrano il doppio strato lipidico. Canali denominati acquaporine sono stati riscontrati in molti epiteli e sono responsabili della permeabilità dell acqua a quel livello. Anche nei doppi strati lipidici artificiali, privi di canali, la permeabilità all acqua è superiore a quanto ci si dovrebbe aspettare in base alla sua solubilità nella fase idrocarburica. La spiegazione risiede nel passaggio di acqua o di altre piccole molecole polari non cariche attraverso dei canali temporanei che si formano tra le molecole lipidiche. Molecole parzialmente cariche come il glicerolo hanno bassi valori di K mentre molecole molto lipofiliche come l uretano hanno alti K. Non dipende invece dalle dimensioni delle molecole in quanto proprio il glicerolo e l uretano hanno dimensioni paragonabili. Gli ioni e altre molecole cariche non diffondono per diffusione semplice 16

17 Ioni e piccole molecole cariche e molecole polari come zuccheri, amminoacidi ecc non possono attraversare il doppio strato lipidico a causa della loro forte polarità. Il loro trasporto è mediato da proteine di membrana che possono essere di due tipi: le proteine trasportatrici che traslocano una sostanza da un ambiente all altro e le proteine canale che formano dei pori attraverso la membrana consentendo il passaggio di ioni della dimensione e carica appropriata. Nel caso della diffusione facilitata la relazione è lineare a basse concentrazioni per poi saturare in maniera diversa ad alte concentrazioni. 17

18 In particolare gli ioni e le piccole molecole attraversano la membrana plasmatica attraverso canali ionici. Le proteine canale sono costituite da una o più subunità proteiche i cui componenti attraversano ripetutamente il doppio strato lipidico e si distribuiscono in modo da formare un poro acquoso all interno della membrana. Quando il canale è aperto mette in comunicazione diretta il compartimento intra- ed extracellulare e permette il passaggio rapido e senza ostacoli di decine di milioni di ioni al secondo. Il movimento degli ioni avviene secondo gradiente elettrochimico ed è strettamente condizionato dalla selettività del canale. In assenza di un gradiente elettrochimico un canale ha uguali probabilità di trasportare uno ione in entrambe le direzioni. 18

19 In generale un canale è strutturalmente costituito da un vestibolo piuttosto largo dove lo ione idratato può accedere facilmente per diffusione e da un filtro di selettività, più stretto, che forma il poro vero e proprio e seleziona gli ioni in base alla carica degli aminoacidi che ne rivestono la parete e al diametro dell apertura. Gli ioni devono velocemente deidratarsi per poter permeare e, nel caso di più siti di legame, gli ioni deidratati si spostando saltando da un sito all altro in fila indiana. 19
 e canali con selettività intermedia permeabili a Na+ e K+ oppure a Na+, K+ e Ca2+.")
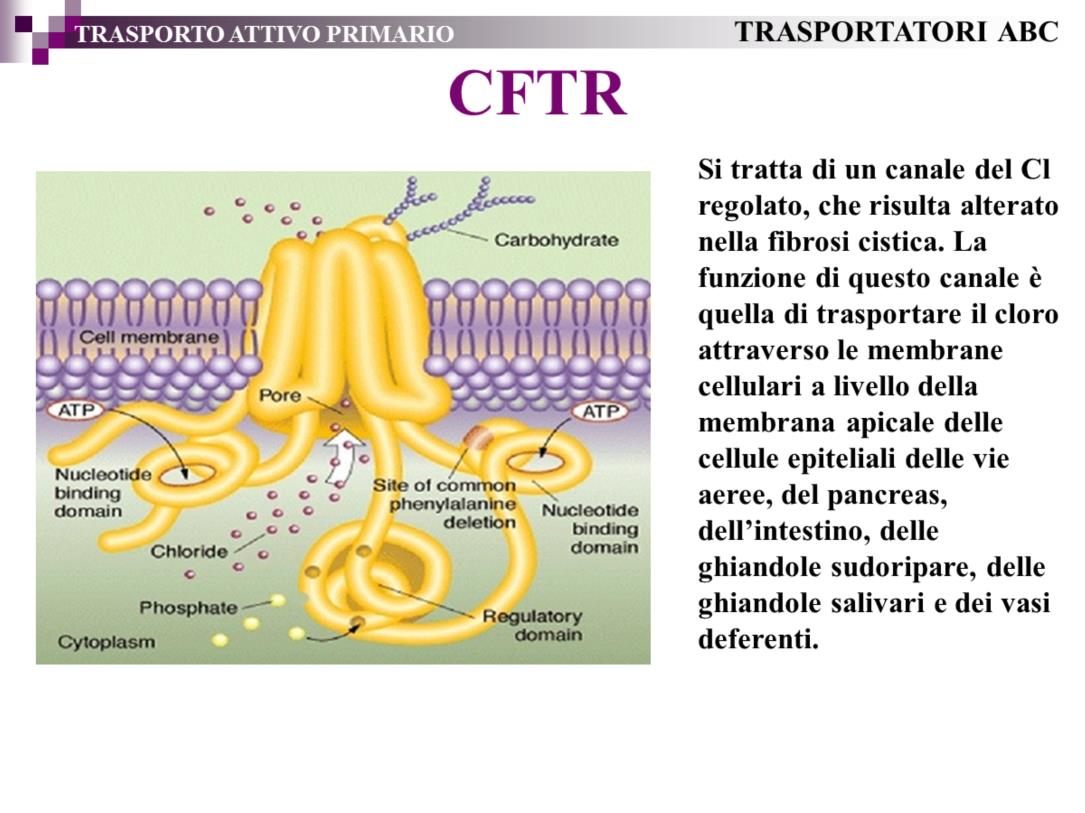
20 Dal punto di vista funzionale i canali si distinguono in base alla loro selettività e in base alla modalità di attivazione. In base alla selettività si distinguono canali per un solo ione (Na+, K+, Ca2+, Cl-) e canali con selettività intermedia permeabili a Na+ e K+ oppure a Na+, K+ e Ca2+. La selettività di un canale è strettamente determinata dal tipo di cariche nette (positive o negative) che formano le pareti del foro di permeazione. Per quanto riguarda i canali del Na+, K+ e Ca2+, i gruppi carichi delle pareti sono costituiti da gruppi carbossilici (COO-) delle catene laterali dell acido glutammico e aspartico a carica netta negativa, mentre per i canali permeabili a Cl- le pareti sono costituite da gruppi amminici (NH3+) degli aminoacidi arginina e lisina a carica netta positiva. 20
 e una a canale aperto (permeabile).")
21 In base alla loro modalità di attivazione i canali si suddividono in tre categorie. In tutti e tre i casi il canale è strutturato in modo tale da avere due conformazioni: una conformazione a canale chiuso (non permeabile) e una a canale aperto (permeabile). Si parla anche di canali a porta (gate) che differiscono in base al meccanismo che determina l apertura o la chiusura del gate: 1. Canali voltaggio dipendenti: la cui probabilità di apertura dipende strettamente dal potenziale elettrico della membrana. Il cambiamento di conformazione del canale da chiuso ad aperto è dovuto allo spostamento di cariche elettriche. 2. canali attivati dai ligandi: la cui probabilità di apertura e chiusura è determinata dall azione di particolari molecole messaggere che agiscono extracellularmente (neurotrasmettitori) e inducono cambi conformazionali capaci di aprire il poro di permeazione. 3. canali attivati da secondi messaggeri la cui probabilità di apertura è condizionata dalla presenza nel liquido intracellulare di ioni e molecole comunenmente indicati come secondi messaggeri intracellulari come Ca2+, ATP, camp, cgmp, IP3). 21

22 Alcune molecole polari come zuccheri, aminoacidi, nucleotidi e alcuni metaboliti diffondono attraverso la membrana utilizzando specifiche proteine trasportatrici (carrier) attraverso una diffusione facilitata. Si sviluppa secondo gradiente in entrambe le direzioni. Le proteine carrier si legano a substrati specifici e li trasportano attraverso la membrana cambiando conformazione. La molecola da trasportare si lega al carrier da un lato della membrana. Questo legame cambia la conformazione del carrier così che il cancello aperto si chiude. Dopo una breve fase di transizione in cui entrambi i cancelli sono chiusi, il lato opposto del carrier si apre dall altro versante della membrana. Il carrier allora rilascia la molecola e il suo rilascio determina una variazione conformazionale che fa ritornare il carrier nella sua conformazione iniziale. La diffusione facilitata mediante carrier ha carattere di uniporto in quanto viene trasportato solo un tipo di molecola e si distingue nettamente dal trasporto accoppiato indicato come cotrasporto che utilizza anche esso proteine trasportatrici ma si tratta di un trasporto attivo secondario. 22

23 23

24 La retta in basso corrisponde alle velocità del trasporto corrispondente ad un processo di diffusione semplice attraverso il doppio strato lipidico. La curva in alto corrisponde ad un processo di diffusione facilitata dalla presenza di un vettore sulla membrana. I due processi (diffusione facilitata e semplice) si distinguono per diverse caratteristiche: 1) Come si può osservare la velocità di trasporto tramite diffusione facilitata è ad ogni concentrazione di substrato notevolmente maggiore di quanto preveda l equazione di Fick. 2) Il processo è altamente specifico in quanto ogni carrier trasporta solo una singola specie di molecole o un singolo gruppo di molecole strettamente affini. 3) il processo è saturabile poiché ad elevati gradienti di concentrazione del substrato si raggiunge una Vmax di trasporto che è un valore limite al quale tende la cinetica della reazione di trasporto a valori di concentrazione di substrato infiniti. 4) La cinetica della diffusione facilitata è descritta dall equazione di Michaelis- Menten. Assumendo che una data sostanza S sia inizialmente presente solo all esterno della membrana e che la velocità sia valutata a tempi iniziali tale equazione descrive numericamente la velocità di trasporto V in funzione della velocità massima Vmax che si raggiunge a valori infiniti del substrato in cui tutte le molecole del trasportatore sono legate ad 1 molecola di substrato; Vmax è proporzionale sia al numero di trasportatori per unità di superficie cellulare che alla velocità di trasporto di ogni singolo trasportatore. 24

25 Km corrisponde alla concentrazione della sostanza alla quale si raggiunge la velocità semimassimale; S è la concentrazione della sostanza in esame. Km, cioè la costante di Michaelis-Menten è un indice dell affinità della sostanza in esame per la proteina carrier, ed in particolare è inversamente correlata all affinità di legame substrato-trasportatore; di conseguenza quanto più basso è il valore di Km tanto maggiore è l affinità del substrato per il trasportatore. Un altra caratteristica dei processi di diffusione facilitata che li differenzia dalla diffusione semplice è che essi possono essere inibiti. Nel caso più semplice cioè l inibizione competitiva si verifica una riduzione del trasporto di una molecola A, es. D-glucosio in presenza di una molecola B, ed. D- galattosio che le somiglia dal punto di vista strutturale e compete con il D-glucosio per il legame al trasportatore. Cineticamente questo tipo di inibizione è facilmente distinguibile da inibizioni di tipo non-competititvo o acompetitivo in quanto si verifica un variazione della costante di Michalis-Menten verso valori maggiori, e di conseguenza in direzione di una riduzione dell affinità enzima-substrato. 25

26 I trasportatori GLUT sono l esempio più studiato di diffusione facilitata a carattere di uniporto. 26

27 Nell ambiente intracellulare le concentrazioni di glucosio sono molto inferiori a quelle vigenti all esterno e quindi il substrato si stacca dal suo sito di legame. Le concentrazioni intracellulari di D-glucosio si mantengono a bassi livelli in quanto una esochinasi lo trasforma in glucosio-6-fosfato, che non è riconosciuto dal trasportatore. Di conseguenza il gradiente di concentrazione favorevole all ingresso di D- glucosio rimane inalterato. Una volta che lo zucchero si è staccato il sistema di trasporto scarico può riconvertirsi nella conformazione E2 e riprendere così il ciclo di trasporto. 27

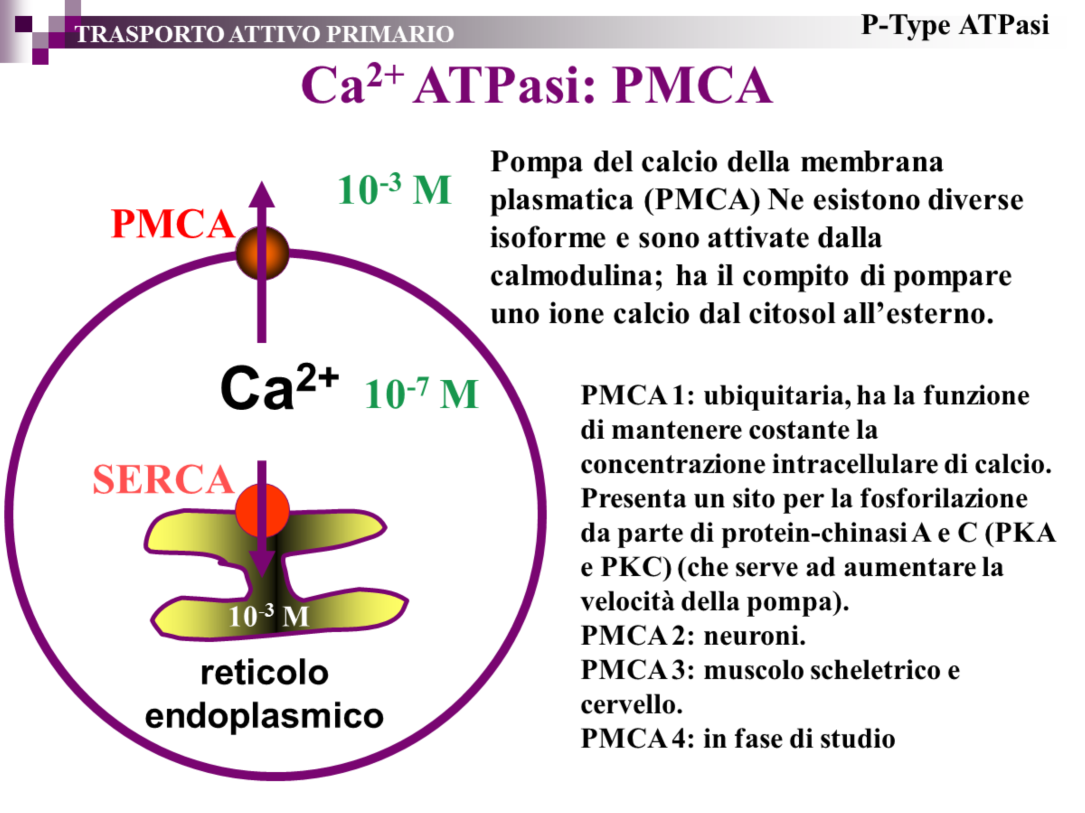
28 Al momento si conoscono 6 isoforme di trasportatori GLUT con diversa affinità per il glucosio. Con l eccezione di GLUT 5 che trasporta principalmente fruttosio, tutte le altre isoforme trasportano glucosio e sono distribuite eterogeneamente nei vari tessuti. La prima isoforma denominata GLUT1 è presente in molti tessuti, compresa la membrana eritrocitaria, ed è denominato costitutivo; l isoforma 2 è maggiormente espressa negli epiteli intestinale e renale dove facilità il flusso trans-epiteliale del glucosio; l isoforma 3 è caratteristica del tessuto nervoso; mentre la 4 maggiormente espressa negli epatociti è estremamente importante nelle patologia del diabete mellito, in quanto è l isoforma soggetta al controllo dell insulina; infine il GLUT5 espresso a livello intestinale, ha una discreta affinità per il fruttosio favorendo il trasporto trans-membrana di questo zucchero. 28

29 A seconda di come viene utilizzato l ATP, il trasporto attivo può essere suddiviso in trasporto attivo primario che utilizza direttamente l energia che deriva dal legame fosfato dell ATP e il trasporto attivo secondario che utilizza invece l energia di un gradiente di concentrazione che è stato precedentemente creato da un trasporto attivo primario. È ovvio quindi che tutti i trasporti attivi secondari dipendono strettamente dai trasporti attivi primari che utilizzano ATP per mantenere i gradienti di concentrazioni necessari per il trasporto attivo secondario. I meccanismi delle proteine responsabili del trasporto attivo assomigliano per alcuni aspetti a quelli della diffusione facilitata. I substrati, ioni o molecole, si legano alla proteina carrier, che cambia conformazione rilasciando il substrato nel lato opposto. Esistono però due sostanziali differenze per quanto riguarda le proteine del trasporto attivo primario: 1) il cambio di conformazione avviene grazie all apporto di energia dell ATP, mentre nel caso della diffusione facilitata non è richiesta alcuna energia metabolica e 2) l affinità della proteina carrier per il ligando si modifica nel passaggio da un lato all altro della membrana. Solo in questo modo possono essere legati e ceduti ioni contro gradiente 29

30 Il trasporto attivo primario utilizza proteine di membrana che cambiano conformazione utilizzando l energia chimica derivata dall idrolisi dell ATP in ADP e fosfato. In questo senso tali proteine sono dei veri e propri enzimi e vengono comunamente definite ATPasi. Possiamo distinguere quattro classi di proteine di trasporto ATP-dipendenti. Le pompe della classe p (P-type ATPasi) sono costituite da due diversi peptidi, alfa e beta, e passano in uno stato fosforilato durante il ciclo di trasporto. La sequenza attorno al residuo che viene fosforilato, situato nelle subunità alfa è omologa nelle diverse pompe. Le pompe delle classi F e V non danno origine ad intermedi fosforilati. Le loro strutture sono simili e contengono proteine simili, ma le loro subunità non hanno alcuna relazione con quelle delle pompe della classe P. Tutti i membri dell'ampia superfamiglia di proteine ABC (ATP-binding cassette transporter) contengono quattro domini essenziali: due domini transmembrana (T) e due domini citosolici leganti ATP (A) che accoppiano l'idrolisi di ATP al trasporto di un dato soluto. In alcune proteine ABC, questi domini sono presenti come subunità separate (come nel caso illustrato) mentre, in altre, risultano fusi in un singolo polipeptide. 30

31 31

32 La pompa Na + /K + ATPasi è stata la prima pompa ad essere studiata. 32

33 Ha il compito principale di mantenere stabili i gradienti di concentrazione di sodio e potassio ai lati della membrana. Oltre a possedere tutti i requisiti dei sistemi di trasporto facilitato precedentemente analizzati (velocità, specificità, saturabilità, etc..), in analogia a tutti i sistemi di trasporto attivo primario è caratterizzata funzionalmente dal lavorare contro gradiente di concentrazione sia dello ione sodio che del potassio in quanto trasporta il Na dall ambiente interno dove è meno concentrato a quello esterno dove è più concentrato; viceversa il potassio viene traslocato dall ambiente extracellulare a minor concentrazione a quello intracellulare a concentrazione maggiore. 33
. Si legano quindi 3 ioni Na.")
34 La pompa dal versante intracellulare ha un alta affinità per il Na e una bassa affinità per il K. Ciò facilita l attacco degli ioni Na ai propri siti di legame a discapito della loro bassa concentrazione all interno della cellula (circa 9 mm). Si legano quindi 3 ioni Na. A questo punto avviene la fosforilazione dei siti di legame intracellulari dell ATP (step 2) che induce il cambio di conformazione necessario per permettere il cambio di direzione della pompa e la liberazione del Na nel liquido extracellulare (step 3 e 4). Nello stato fosforilato, la pompa ha un alta affinità per il K e una bassa affinità per il Na sul lato extracellulare. Quindi il Na viene rilasciato, nonostante la sua concentrazione all esterno della cellula sia elevata (145 mm) mentre si legano due ioni K pur essendo bassa la loro concentrazione che è intorno a 4 mm (step 5). L attacco degli ioni K sul lato extracellulare attiva la liberazione del gruppo fosfato (step 6) che fa variare la conformazione della pompa (step 7) con il rilascio degli ioni K nel lato intracellulare contro gradiente di concentrazione (140 mm). 34

35 In questo modo il ripetersi del ciclo permette di mantenere bassi i livelli di Na e alti i livelli di K all interno della cellula e una situazione opposta all esterno. L 35
 o da agenti specifici, come il glicoside cardiaco ouabaina. Il sito di legame per l ouabaina (strofantina G), un glicoside vegetale si trova nelle immediate vicinanze dei siti per il potassio.")
36 E un processo ATP-dipendente, di conseguenza è soggetto all inibizione da parte di veleni del metabolismo che limitano la disponibilità di ATP (disaccoppianti respirazione mitocondriale, etc..) o da agenti specifici, come il glicoside cardiaco ouabaina. Il sito di legame per l ouabaina (strofantina G), un glicoside vegetale si trova nelle immediate vicinanze dei siti per il potassio. Il legame della ouabaina porta all arresto della pompa quando questa passa dalla configurazione E 2 a quella E 1. Ciò comporta sia l inibizione del trasporto ionico che dell idrolisi dell ATP. L inibizione è reversibile, l affinità dell inibitore per la pompa è variabile sia tra specie diverse che, nell ambito della stessa specie, a livello di tessuti diversi. Infine la > parte dei trasporti attivi primari traslocano cariche elettriche attraverso la membrana determinando una separazione di cariche tra i due lati della membrana; in questo modo la Na-K-ATPasi incide direttamente sul potenziale elettrico esistente fisiologicamente a cavallo delle membrane plasmatiche oltre a contribuirvi indirettamente creando la distribuzione asimmetrica del sodio e del potassio tra l ambiente intra ed extra-cellulare. La costante di legame per il Na è di 0,2 mm; la Km per il K è di 0,05 mm. 36

37 37
38 Sono riportati i modelli strutturali delle P-ATPasi di tipo I (batteriche) e di tipo II (degli eucarioti), ricavate da profili di idrofobicità (Kyte-Doolittle algorithm). Le 2 regioni citosoliche comuni sono denominate regione B e dominio C; questo dominio presenta una parte denominata hinge e legami di connessione con le parti intramembrana. Il dominio C è quello cataliticamente attivo, essendovi localizzato il sito di fosforilazione verso l N-terminale ed il sito di legame dell ATP nella regione centrale e carbossi terminale. E presente una regione hinge comune a molte fosfochinasi che lo connette ai domini trans-membrana. Il ruolo di questa regione dovrebbe essere quello di favorire l interazione reciproca tra il sito catalitico ed il sito di legame dei cationi durante la traslocazione. I putativi domini trans-membrana (ad alfa elica) sono 6 nel caso delle P-ATPasi di tipo I e 10 nel caso delle P- ATPasi di tipo II. Nel caso delle ATPasi di tipo I si potrebbe dedurre dall analisi del profilo di idrofobicità, la presenza di due addizionali domini trans-membrana denominati Ma e Mb. Questi addizionali motivi transmembrana sono preceduti nella regione N-terminale dalla presenza di un sito di legame per i metalli pesanti. Le ATPasi di tipo II sono distinguibili dal numero maggiore di domini trans-membrana nella regione carbossi terminale. Dal punto di vista filogenetico si suppone che le P-ATPasi derivino da un comune antenato, un trasportatore di cationi, da cui siano derivate in seguito ad una divisione ancestrale a livello batterico, le P-ATPasi di tipo I (che trasportano metalli pesanti) e quelle di tipo II presenti nei moderni procarioti come l ATPasi che trasporta magnesio del S. typhimurium e la ATPasi che trasporta calcio del Synechoccus). Nel corso dell evoluzione ulteriori diversificazioni delle P-ATPasi di tipo II ha dato origine a quelle IIA (Serca, Na-K-ATPasi; H,K-ATPAsi e di tipo IIB (PMCA; e ATPasi dei protoni predominante nelle piante e nei funghi dove è necessaria all ingresso di nutrienti e altre sostanze utili alle funzioni cellulari). Attualmente si riconoscono più di 80 membri diversi appartenenti alla famiglia delle P-ATPAsi. 38

39 La pompa Ca 2+ ATPasi è una proteina integrale di membrana (PM 100 kda) con 10 domini transmembrana. Ha le caratteristiche di un uniporto e trasporta fuori dalla cellula 2 ioni Ca per ogni molecola di ATP utilizzata in ogni ciclo. È elettrogenica ed ha l importante ruolo di mantenere bassa la concentrazione del Ca intracellulare che è una condizione vitale per la sopravvivenza e la funzionalità cellulare. La pompa del Ca è presente non solo a livello del plasmalemma ma è attiva anche sulla membrana del reticolo endoplasmatico e sarcoplasmatico e nei mitocondri e aiuta a mantenere bassi i livelli di Ca citoplasmatico trasportabto lo ione all interno di questi organuli contro un forte gradiente di concentrazione. Le due isoforme hanno diverse caratteristiche molecolari. La PMCA ha una maggiore affinità per il Ca ed è praticamente sempre in funzione per mantenere a livello basale la concentrazione di ca intracellulare. La serca entra in funzione solo quando la concentrazione di Ca aumenta di un ordine di grandezza. Non si conoscono ancora inibitori specifici per la PMCA mentre la SERCA è inibita dalla tapsigargina. 39
40 40

41 41
. In questo modo permette l acidificazione del liquido extracellulare pur essendo bassi i livelli di H+ citoplasmatici (ph 7,4, [H+] = 4*10-8 M).")
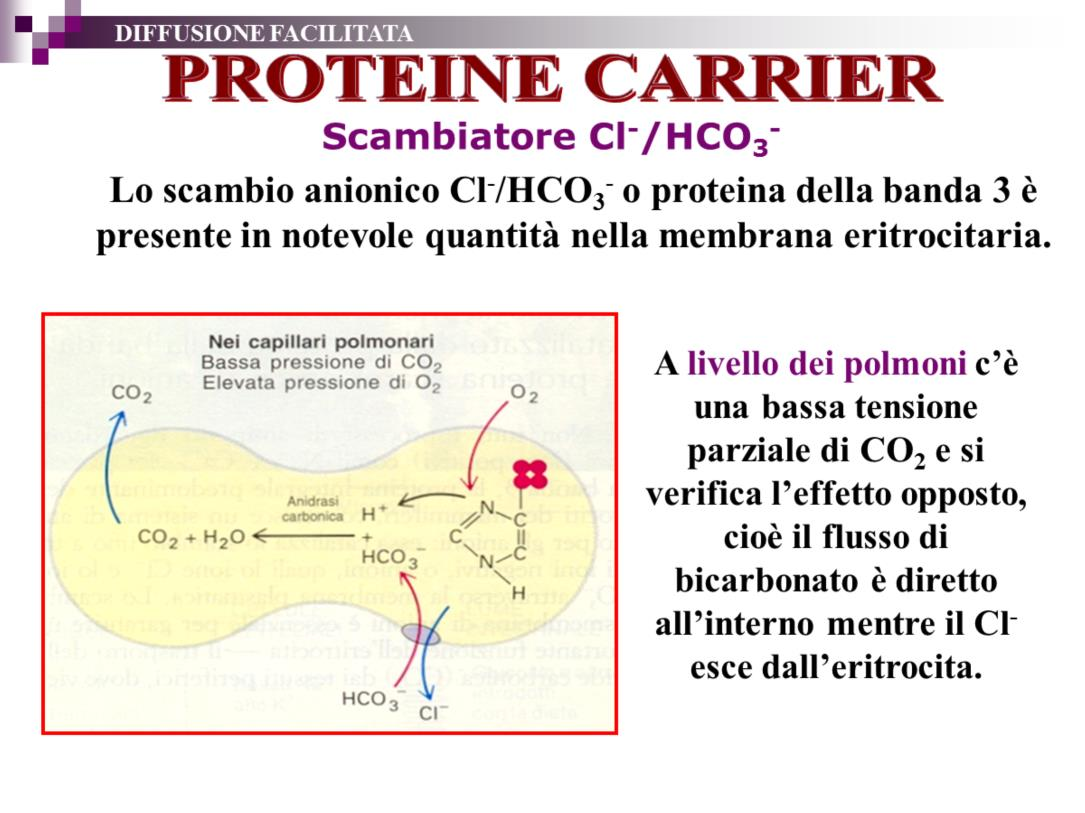
42 La pompa H + /K + ATPasi o pompa protonica è un trasportatore strutturalmente molto simile alla Na + /K + ATPasi che trasporta 4 H+ fuori dalla cellula e 4 ioni K+ all interno della cellula con stechiometria di 1:1 (quindi non è elettrogenica). In questo modo permette l acidificazione del liquido extracellulare pur essendo bassi i livelli di H+ citoplasmatici (ph 7,4, [H+] = 4*10-8 M). La pompa è localizzata sulla parete delle cellule ossintiche (o parietali) delle ghiandole gastriche e svolge un ruolo centrale nella produzione dell HCl del succo gastrico. Permette la produzione di acido a concentrazioni massime di 100 mm (ph=1) ed è quindi in grado di mantenere elevatissimi gradienti di H+ tra l interno e l esterno della cellula. La sua importanza è anche legata alla recente scoperta che derivati dell omeprazolo si legano alla subunità della pompa e ne bloccano l attività. Questi farmaci sono attualmente ampiamente utilizzati come antiulcerosi e per la cura di diverse disfunzioni gastriche. Il lansoprazolo, come tutti gli inibitori di pompa, inibisce questo processo con conseguente immediato blocco della produzione di acido cloridrico, indipendentemente dallo stimolo che l'ha prodotto. Nel lume gastrico il ph viene mantenuto forzatamente al di di sopra di 3-4. La H + -K + -ATPasi ha una struttura molto simile alla pompa sodio-potassio: anche qui si riconoscono due subunità α e due β con la α che è responsabile della funzionalità della pompa protonica contenendo i siti di legame degli ioni, di fosforilazione e di attività ATPasica. 42
43 43
 oppure in direzione opposta (antiporto).")
44 Il trasporto attivo secondario sfrutta l energia cinetica di uno ione che si muove passivamente secondo il suo gradiente di concentrazione per trasportare una molecola o un altro ione contro gradiente. Le molecole o gli ioni cotrasportati possono muoversi nella stessa direzione (simporto) oppure in direzione opposta (antiporto). I più comuni sistemi di trasporto attivo secondario sono mediati da proteine transmembrana che sfruttano il gradiente di concentrazione del Na+ creato dalla Na+-K+ ATPasi. In tal caso, l alta concentrazione extracellulare di Na+ favorisce il legame dello ione sul lato esterno del trasportatore, che a sua volta è facilitato a legare una molecola o un altro ione e a trasportarli dal lato opposto a quello in cui si trovano, senza ulteriore spesa di energia. 44

45 45

46 L antiporto Na + /Ca 2+ ha un ruolo chiave nel controllo del Ca intracellulare in tutte le cellule, dove è costantemente attivato. Trasporta 3 ioni Na dentro la cellula e 1 ione Ca fuori ed è quindi elettrogenico. È di particolare importanza nelle cellule cardiache quando la concentrazione di Ca, che aumenta durante la contrazione sistolica, deve essere velocemente riportata al suo livello normale per permettere il rilasciamento ventricolare 46

47 Un paziente con insufficienza cardiaca possiede un cuore ipodinamico caratterizzato da una bassa gittata cardiaca (volume di sangue espulso dai ventricoli al minuto) insufficiente a sostenere le richieste metaboliche del suo corpo. Il metodo più efficace per migliorare l attività cardiaca consiste nell aumentare la contrattilità miocardica che è marcatamente Ca2+ dipendente. Un aumento del Ca citoplasmatico produce un aumentata contrattilità cardiaca che a sua volta produce un aumento del volume sistolico (volume di sangue espulso durante un singolo battito cardiaco) e della gittata cardiaca. I glicosidi cardiaci come la digitossina, la digossina e l ouabaina, sono usati da moltissimi anni per la cura dell insufficienza cardiaca. Questi farmaci si legano selettivamente e inibiscono la Na+/K+-ATPasi della membrana delle cellule del miocardio. In tal modo il Na+ si accumula in cellula riducendo notevolmente l attività dello scambiatore Na+/Ca2. Come risultato finale il livello citoplasmatico di Ca aumenta e aumenta di conseguenza la contrattilità miocardica, il volume sistolico e la gittata cardiaca. 47

48 L antiporto Na + /H + è svolge un ruolo determinante nel controllo del ph intracellulare di tutte le cellule in quanto trasporta ioni H+ fuori dalla cellula e ioni Na dentro in rapporto 1:1. L antiporto Na + /H + è particolarmente importante a livello renale. La sua localizzazione sull orletto a spazzola delle cellule epiteliale del tubulo contorto prossimale permette, infatti, di arricchire il liquido del lume tubulare di ioni H+ e di acidificare le urine. 48

49 Trasportatori per gli oligopeptidi dipendenti dai protoni sono stati identificati in batteri, funghi, piante, e cellule epiteliali intestinali e renali dei mammiferi. Essi rappresentano una famiglia denominata POPT conservata nel corso dell evoluzione. Il carattere distintivo di questa classe di trasportatori è di utilizzare come driving force il gradiente elettrochimico dei protoni per trasportare peptidi in cellula contro gradiente di concentrazione. A livello intestinale e renale esiste un microambiente acido a livello luminale creato principalmente dall attività del contro-trasporto Na/H. Questo gradiente viene sfruttato dai sistemi di trasporto per dipeptidi che riconoscono inoltre altri substrati come antibiotici beta lattamici; inibitori dell enzima convertitore di angiotensina (ACE). Dal punto di vista strutturale PepT1 presenta 12 domini trans-membrana e regioni extracellulari di glicosilazione; dal lato citosolico presenta siti di fosforilazione da parte della protein chinasi C e della protein chinasi A. Per quel che riguarda il funzionamento è stato suggerito che la formazione del complesso ternario è ottenuta grazie ad un legame dipendente dal potenziale elettrico di H + e del substrato ai siti proteici del trasportatore. Nel sito proteico è stato possibile riconoscere. Il complesso va incontro ad un cambio conformazionale permettendo la traslocazione della molecola di peptide, di una molecola d acqua e un carica positiva. In questo meccanismo il potenziale di membrana (Vm) determina la velocità massima di trasporto e l affinità di legame per i soluti. Dal punto di vista delle richieste steriche per il riconoscimento del substrato, il trasportatore può riconoscere di e tri-peptidi e substrati simili a peptidi, R denota la catena laterale di amino acidi con preferenza per residui idrofobici. P1; P2; P3 indicano le tasche idrofobiche proposte essere presenti nel domino di legame del substrato del trasportatore. 49

50 50

51 Il cotrasporto Na-D-glucosio è un esempio classico di trasporto attivo secondario. La sua funzione principale è quella di mediare sia l assorbimento di glucosio a livello intestinale che il riassorbimento dello zucchero a livello renale. 51

52 Il meccanismo di traslocazione è simile a quelli riportati sino ad ora. Anche in questo caso si ha un oscillazione tra due stati conformazionali E1 e E2 con i siti rivolti alternativamente verso l ambiente extra ed intra-cellulare. In questo caso tuttavia l elemento aggiuntivo è rappresentato dal legame dello ione sodio al trasportatore che induce un cambio conformazionale nella tasca di legame per lo zucchero, aumentandone l affinità nei confronti del substrato. Una volta che il D-glucosio si è legato, si ha una traslocazione dei due substrati in direzione dell ambiente intracellulare, dove il sodio si stacca dal suo sito di legame poiché le sue concentrazioni intracellulari sono basse, il distacco del sodio determina una riduzione dell affinità per il D-glucosio che si stacca dalla sua tasca di legame, pur essendo le sue concentrazioni citosoliche abbastanza elevate. 52
53 Al momento sono conosciute due isoforme SGLT-1 e SGLT-2 del trasportatore Na-dipendente di D- glucosio. Il primo è presente sia sulle membrane apicali dell intestino tenue che nella seconda parte del tubulo contorto prossimale, è caratterizzato da una Km di circa 0.1 mm per il D-glucosio, una stechiometria di cotrasporto Na-D-glucosio di 2 ioni sodio per molecola di D-glucosio trasportata; è un trasporto elettrogenico. SGLT-2 è stato riscontrato nelle prime porzioni del tubulo contorto prossimale e si distingue cineticamente da SGLT-1 in quanto ha un affinità molto inferiore ma una velocità di trasporto molto più elevata rispetto a SGLT-1. Questa distribuzione regionale delle due isoforme di SGLT nel tubulo contorto prossimale assicura il riassorbimento totale del D-glucosio. Infatti nelle regioni adiacenti alla capsula di bowman dove predomina SGLT-2 questo trasporto capta efficientemente il D-glucosio ultrafiltrato ad alta concentrazione, grazie alla sua Km che è vicina alle concentrazioni di glucosio e alla sua alta velocità di trasporto. Allorquando le concentrazioni di D-glucosio nell ultrafiltrato si riducono, man mano che questo scorre nel tubulo prossimale, questo sistema non sarebbe più efficiente a continuare il processo di riassorbimento a causa della sua bassa affinità per il D-glucosio e di conseguenza interviene l isoforma SGLT-1 localizzata più a valle, che opera efficientemente a basse concentrazioni di D-glucosio a causa della sua elevata affinità per lo zucchero. SGLT1 è costituito da 664 residui aacidici. La struttura secondaria del trasportatore si articola in 14 alfaeliche trans-membrana (al contrario dei trasporti facilitati del glucosio che possiedono solo 12 eliche) con dei domini citosolici. La regione del trasportatore che lega e trasloca sodio, identificati tramite studi con trasportatori chimerici, comprendono la regione N-terminale sino a loop extracellulare che connette i domini trans-membrana 4 e 5; le regioni di legame e di traslocazione includono domini TM da 10 a 13. SGLT1, al contrario di altre proteine carrier manca di un grande dominio citosolico, il chè suggerisce l esistenza di addizionali sub-unità regolatorie. Recentemente è stato proposto da Rehiardt ed altri che una proteina denominata RS1, ancorata alle membrana plasmatiche apicali aumenti la velocità massima di uptake di D-glucosio e vada a modulare l attività di altri sistemi di trasporto. Un altro tipo di regolazione sembra essere effettuata dalla protein chinasi C e protein kinasi A dato che domini citosolici del trasportatore contengono regioni suscettibili a fosforilazione da parte di queste protein chinasi. Di conseguenza è un sistema di trasporto sottoposto sia a regolazione tramite AMPc, 53
54 diacil-glicerolo e calcio. Lavori recenti suggeriscono che PKA e PKC aumentino l attività del cotrasporto mediante una maggiore espressione di questo a livello delle membrane plasmatiche apicali. Il meccanismo di questa regolazione è mediato da endocitosi o esocitosi di vescicole situate sotto la membrana pronte ad essere incorporate nel momento della stimolazione da parte ormonale. SGLT-1 appartiene ad una famiglia cui appartengono più di 30 classi di trasportatori. Questi condividono caratteristiche strutturali e funzionali omologhe. Posseggono 13 o più eliche transmembrana, utilizzano il gradiente elettrochimico sia del sodio che dei protoni per trasportate una varietà di substrati che includono zuccheri, aacidi, anioni organici, urea, vitamine, etc.. Dovrebbero condividere un comune meccanismo di trasporto sebbene il substrato cotrasportato possa variare considerevolmente. 53

55 Sia il glucosio che gli aacidi sono trasportati dal lume intestinale o dei tubuli renali all interno delle cellule assorbenti rappresentate in figura e da queste al sangue. I due stadi di questo processo coinvolgono due distinti processi di trasporto: uno localizzato a livello apicale, l altro a livello basolaterale. Per quel che riguarda il trasporto basolaterale, si tratta di un sistema di trasporto facilitato per il glucosio appartenente alla famiglia GlUT, precedentemente descritta. A livello apicale è invece presente il trasportatore che sfrutta il gradiente elettrochimico favorevole allo ione sodio per traslocare in cellula, il D-glucosio contro elevati gradienti di concentrazione. Si può calcolare che un gradiente elettrochimico per il sodio esistente possa generare una concentrazione di glucosio in cellula 100 volte superiore. Il potenziale elettrochimico favorevole all ingresso del sodio è mantenuto dal trasporto attivo Na-K-dipendente localizzato sulla membrana basolaterale. Di conseguenza si parla di trasporto attivo in quanto lavora contro gradiente di concentrazione del D-glucosio, secondario perché sfrutta secondariamente l energia fornita dall ATP tramite il lavoro svolto da un trasporto attivo primario, in questo caso, la Na + /K + -ATPasi basolaterale. 54

56 Oltre al trasporto attivo secondario Na-dipendente esistono anche svariati trasportatori che non sono Na dipendenti. Tra questi, il più importante per il suo ruolo nel controllo dell equilibrio acido-base a livello cellulare è l antiporto Cl - /HCO 3- che trasporta ioni bicarbonato fuori dalla cellula e ioni Cl- dentro in un rapporto 1:1. La fuoriuscita di bicarbonato dalla cellula avviene secondo gradiente ed è accoppiata all ingresso di cloro, che può avvenire anche controgradiente. 55

57 Lo scambiatore svolge un ruolo fondamentale a livello degli eritrociti dove contribuisce all arricchimento di bicarbonato nel plasma e più in generale al trasporto della CO2 e alla regolazione della respirazione. 56
58 57

59 Il trasportatore Cl/HCO3 è particolarmente importante anche in altri distretti dove svolge importanti funzioni come: Il riassorbimento di bicarbonato a livello della membrana basolaterale del digiuno e del tubulo contorto prossimale e distale del rene L ingresso di Cl esterno nelle cellule ossintiche dell epitelio gastrico, che rappresenta la riserva di ioni Cl- utilizzati per la produzione di HCl del succo gastrico. La produzione di bicarbonato del succo pancreatico che serve per mantenere il ph del succo pancreatico a valori basici (ph 8-9) che aiuta a controbilanciare il basso ph del chimo nel lume duodenale e a ottenere ph ottimali per il funzionamento degli enzimi pancreatici. 58

60 Anche l endocitosi è un meccanismo attraverso il quale grandi molecole o particelle entrano nella cellula. L endocitosi mediata da recettore avviene in regioni specifiche della membrana cellulare note come fossette rivestite da clatrina che sono degli incavi in cui il lato citoplasmatico della membrana ha un alta concentrazione della proteina clatrina. Nella prima tappa del processo i substrati extracellulari che devono essere portati all interno della cellula si legano a specifici recettori di membrana. Il complesso recettore-ligando migra lungo la superficie cellulare fino ad incontrare una fossetta rivestita. A questo punto la membrana si ritira verso l interno (si invagina) si stacca quindi dalla membrana e diventa una vescicola citoplasmatica. Le molecole di clatrina vengono rilasciate e ritornano sulla membrana, nella vescicola il recettore e il ligando si separano, lasciano il ligando in un endosoma che viene portato a livello di un lisosoma se il ligando deve essere distrutto, oppure a livello dell apparato del Golgi se il ligando deve essere elaborato. Nel frattempo i recettori possono essere riutilizzati e riportati sulla membrana mediante un meccanismo noto come esocitosi. In questo modo vengono trasportati: ormoni proteici, fattori di crescita e proteine plasmatiche che funzionano come carrier per il ferro e il colesterolo. 59
Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori
 Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori I trasportatori somigliano agli enzimi: catalizzano la traslocazione dei substrati da una parte all altra della membrana
Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori I trasportatori somigliano agli enzimi: catalizzano la traslocazione dei substrati da una parte all altra della membrana
5. Trasporto di membrana
 5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
Permeabilità Di Membrana Cellulare
 Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Fisiologia cellulare e Laboratorio di colture cellulari
 Corso di laurea in Biotecnologie UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di colture cellulari Prof.ssa Luisa Gioia possibili sistemi di trasporto della cellula TRASPORTO TRANS-MEMBRANA
Corso di laurea in Biotecnologie UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di colture cellulari Prof.ssa Luisa Gioia possibili sistemi di trasporto della cellula TRASPORTO TRANS-MEMBRANA
Trasporto di membrana
 Trasporto di membrana CdL Tecnici di Lab Biomedico AA. 2011/12 Prof.ssa Frabetti Trasporto di membrana molecole piccole molecole grosse trasporto passivo trasporto attivo (con pompe o cotrasportatori)
Trasporto di membrana CdL Tecnici di Lab Biomedico AA. 2011/12 Prof.ssa Frabetti Trasporto di membrana molecole piccole molecole grosse trasporto passivo trasporto attivo (con pompe o cotrasportatori)
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA)
") Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Trasporto Attivo SECONDARIO. Trasporto mediato da proteine di membrana. Caratteristiche Strutturali
 Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Richiede ATP DIFFUSIONE ENDOCITOSI ESOCITOSI Diffusione Semplice Diffusione Facilitata Trasporto Attivo SECONDARIO Crea gradienti
Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Richiede ATP DIFFUSIONE ENDOCITOSI ESOCITOSI Diffusione Semplice Diffusione Facilitata Trasporto Attivo SECONDARIO Crea gradienti
Proprietà di permeabilità della membrana
 Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra ). La stabilità del ph è insidiata da
. La stabilità del ph è insidiata da") Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
2. permettere un continuo scambio di ioni e molecole fra i due compartimenti
 La membrana plasmatica ha il compito di 1. isolare il citoplasma dall ambiente extracellulare 2. permettere un continuo scambio di ioni e molecole fra i due compartimenti Trasporto passivo: passaggio di
La membrana plasmatica ha il compito di 1. isolare il citoplasma dall ambiente extracellulare 2. permettere un continuo scambio di ioni e molecole fra i due compartimenti Trasporto passivo: passaggio di
FENOMENI DI TRASPORTO. I trasporti attraverso le membrane biologiche
 FENOMENI DI TRASPORTO I trasporti attraverso le membrane biologiche 1 La struttura a doppio strato lipidico delle membrane biologiche condiziona fortemente il movimento di gas, ioni e molecole attraverso
FENOMENI DI TRASPORTO I trasporti attraverso le membrane biologiche 1 La struttura a doppio strato lipidico delle membrane biologiche condiziona fortemente il movimento di gas, ioni e molecole attraverso
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da
. La stabilità del ph è insidiata da") Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
Le variazioni di ph nei liquidi organici devono mantenute entro limiti molto ristretti (mammiferi tra 6.8 7.8). La stabilità del ph è insidiata da acidi forti (solforico, cloridrico e fosforico) prodotti
MEMBRANA CELLULRE. La membrana plasmatica svolge molteplici funzioni:
 MEMBRANA CELLULRE MEMBRANA CELLULRE La membrana plasmatica svolge molteplici funzioni: Regola gli scambi di sostanze tra interno ed esterno delle cellule e degli organismi unicellulari Protegge l integrità
MEMBRANA CELLULRE MEMBRANA CELLULRE La membrana plasmatica svolge molteplici funzioni: Regola gli scambi di sostanze tra interno ed esterno delle cellule e degli organismi unicellulari Protegge l integrità
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Il funzionamento della cellula Le cellule sono veri e propri esseri viventi: sono capaci di alimentarsi, di crescere, di produrre nuove sostanze, di riprodursi, di espellere rifiuti
METABOLISMO CELLULARE Il funzionamento della cellula Le cellule sono veri e propri esseri viventi: sono capaci di alimentarsi, di crescere, di produrre nuove sostanze, di riprodursi, di espellere rifiuti
Permeabilità di un doppio strato fosfolipidico artificiale (privo di proteine)
") Permeabilità di un doppio strato fosfolipidico artificiale (privo di proteine) Possono superare la barriera fosfolipidica: le molecole idrofobiche come i lipidi, piccole molecole polari senza carica Permeabilità
Permeabilità di un doppio strato fosfolipidico artificiale (privo di proteine) Possono superare la barriera fosfolipidica: le molecole idrofobiche come i lipidi, piccole molecole polari senza carica Permeabilità
Caratteristiche Strutturali
 Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Richiede ATP DIFFUSIONE ENDOCITOSI ESOCITOSI Diffusione Semplice Diffusione Facilitata Trasporto Attivo SECONDARIO Crea gradienti
Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Richiede ATP DIFFUSIONE ENDOCITOSI ESOCITOSI Diffusione Semplice Diffusione Facilitata Trasporto Attivo SECONDARIO Crea gradienti
Plasma membrane. Endoplasmic reticulum. Nucleus. Golgi apparatus. Mitochondrion Lysosome. Ribosome
 Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
BIOFISICA DELLE MEMBRANE
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 BIOFISICA DELLE MEMBRANE Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 BIOFISICA DELLE MEMBRANE Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Ioni fosfato e proteine (anioni) sono prevalenti nella cellula
 sono prevalenti nella cellula") Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Funzioni proteine di membrana
 Funzioni proteine di membrana Ancoraggio Le integrine svolgono un ruolo importante nella trasmissione dei segnali tra le cellule h7p://www.nature.com/fer=lity/content/images/ncb- nm- fer=litys57- f3.jpg
Funzioni proteine di membrana Ancoraggio Le integrine svolgono un ruolo importante nella trasmissione dei segnali tra le cellule h7p://www.nature.com/fer=lity/content/images/ncb- nm- fer=litys57- f3.jpg
Le membrane cellulari
 Le membrane cellulari La struttura delle membrane biologiche Le membrane cellulari sono formate da un doppio strato fosfolipidico dove si trovano varie proteine, che possono essere: periferiche: si trovano
Le membrane cellulari La struttura delle membrane biologiche Le membrane cellulari sono formate da un doppio strato fosfolipidico dove si trovano varie proteine, che possono essere: periferiche: si trovano
11/10/16. Il concetto di MOSAICO FLUIDO. Fosfolipidi Colesterolo Glicolipidi
 Il concetto di MOSAICO FLUIDO Fosfolipidi Colesterolo Glicolipidi 1 DIFFUSIONE SEMPLICE 2 Pressione osmotica Il problema della pressione osmotica 3 Diverse soluzioni al problema della pressione osmotica
Il concetto di MOSAICO FLUIDO Fosfolipidi Colesterolo Glicolipidi 1 DIFFUSIONE SEMPLICE 2 Pressione osmotica Il problema della pressione osmotica 3 Diverse soluzioni al problema della pressione osmotica
Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA
 Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA Fisiologia cellulare: Membrana cellulare. Trasporto di ioni e molecole attraverso la membrana cellulare. Canali ionici, pompe e proteine vettrici.
Fisiologia Umana - 6 CFU Docente: Loriana Castellani PROGRAMMA Fisiologia cellulare: Membrana cellulare. Trasporto di ioni e molecole attraverso la membrana cellulare. Canali ionici, pompe e proteine vettrici.
Il trasporto attraverso la membrana plasmatica LEZIONE NR PSICOBIOLOGIA
 Il trasporto attraverso la membrana plasmatica LEZIONE NR. 10 - PSICOBIOLOGIA DIFFUSIONE, OSMOSI ED EQUILIBRIO ELETTROCHIMICO L unione di un soluto in una soluzione avviene attraverso un processo denominato
Il trasporto attraverso la membrana plasmatica LEZIONE NR. 10 - PSICOBIOLOGIA DIFFUSIONE, OSMOSI ED EQUILIBRIO ELETTROCHIMICO L unione di un soluto in una soluzione avviene attraverso un processo denominato
Il potenziale di membrana a riposo
 Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti su entambe le facce della membrana; la membrana stessa;
La membrana e il trasporto di membrana
 Clostridium botulinum. Batterio strettamente anaerobio che produce una tossina termostabile che è estremamente neurotossica per l uomo. Può proliferare nelle conserve vegetali e nelle conserve sott olio.
Clostridium botulinum. Batterio strettamente anaerobio che produce una tossina termostabile che è estremamente neurotossica per l uomo. Può proliferare nelle conserve vegetali e nelle conserve sott olio.
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA
 FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
FONDAMENTI ANATOMO-FISIOLOGICI DELL ATTIVITA PSICHICA Il potenziale di membrana a riposo Per poter comprendere il potenziale di membrana a riposo dobbiamo considerare: i fluidi ricchi di sali presenti
LA MEMBRANA CELLULARE O PLASMATICA E LA STRUTTURA CHE DELIMITA ESTERNAMENTE LA CELLULA, SIA EUCARIOTICA CHE PROCARIOTICA, SEPARANDO IL COMPARTO INTRAC
 MEMBRANA CELLULARE LA MEMBRANA CELLULARE O PLASMATICA E LA STRUTTURA CHE DELIMITA ESTERNAMENTE LA CELLULA, SIA EUCARIOTICA CHE PROCARIOTICA, SEPARANDO IL COMPARTO INTRACELLULARE DA QUELLO EXTRACELLULARE.
MEMBRANA CELLULARE LA MEMBRANA CELLULARE O PLASMATICA E LA STRUTTURA CHE DELIMITA ESTERNAMENTE LA CELLULA, SIA EUCARIOTICA CHE PROCARIOTICA, SEPARANDO IL COMPARTO INTRACELLULARE DA QUELLO EXTRACELLULARE.
Il processo di ultrafiltrazione è interamente passivo e dipende sostanzialmente da tre fattori:
 Il processo di ultrafiltrazione è interamente passivo e dipende sostanzialmente da tre fattori: 1) Dalla differenza netta di pressione tra il lume del capillare ed il lume della capsula di Bowman; 2) dalla
Il processo di ultrafiltrazione è interamente passivo e dipende sostanzialmente da tre fattori: 1) Dalla differenza netta di pressione tra il lume del capillare ed il lume della capsula di Bowman; 2) dalla
Attivi (non equilibranti) Passivi (equilibranti) Migrazione attraverso canali membran. Diffusione semplice. Tasporto attivo secondario
 Passivi (equilibranti) Migrazione attraverso canali membran. Diffusione semplice. Tasporto attivo secondario") TRASPORTI Trasporti in forma libera Trasporti mediati Diffusione semplice Migrazione attraverso canali membran. Diffusione facilitata Trasporto attivo primario Tasporto attivo secondario Passivi (equilibranti)
TRASPORTI Trasporti in forma libera Trasporti mediati Diffusione semplice Migrazione attraverso canali membran. Diffusione facilitata Trasporto attivo primario Tasporto attivo secondario Passivi (equilibranti)
Membrana, trasporti, nutrizione, enzimi
 Membrana, trasporti, nutrizione, enzimi LA MEMBRANA PLASMATICA: - DETERMINA UNA COMPARTIMENTAZIONE; - FUNGE DA BARRIERA CON PERMEABILITA SELETTIVA; - TRASPORTA SOLUTI; - RISPONDE A SEGNALI ESTERNI; - PRESIEDE
Membrana, trasporti, nutrizione, enzimi LA MEMBRANA PLASMATICA: - DETERMINA UNA COMPARTIMENTAZIONE; - FUNGE DA BARRIERA CON PERMEABILITA SELETTIVA; - TRASPORTA SOLUTI; - RISPONDE A SEGNALI ESTERNI; - PRESIEDE
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Membrane Biologiche. Barriere per confinare sostanze o attività in ambienti specifici. Costituite da lipidi e proteine. Confini Cellulari Organelli
 Membrane Biologiche Membrane Biologiche Barriere per confinare sostanze o attività in ambienti specifici. Confini Cellulari Organelli Costituite da lipidi e proteine. Sistema di Endomembrane Delimitano
Membrane Biologiche Membrane Biologiche Barriere per confinare sostanze o attività in ambienti specifici. Confini Cellulari Organelli Costituite da lipidi e proteine. Sistema di Endomembrane Delimitano
Trasporto di sostanze in e out dalla cellula
 Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Membrane biologiche e trasporto cellulare
 Membrane biologiche e trasporto cellulare Le membrane biologiche di tutti gli organismi hanno una struttura e una composizione praticamente identica (fa eccezione però la membrana dei cloroplasti) Parete
Membrane biologiche e trasporto cellulare Le membrane biologiche di tutti gli organismi hanno una struttura e una composizione praticamente identica (fa eccezione però la membrana dei cloroplasti) Parete
Il rene regola l escrezione dei diversi soluti indipendentemente l uno dall altro, controllandone la velocità di riassorbimento.
 Il rene regola l escrezione dei diversi soluti indipendentemente l uno dall altro, controllandone la velocità di riassorbimento. Lungo i tubuli renali viene riassorbita la maggior parte dei soluti e il
Il rene regola l escrezione dei diversi soluti indipendentemente l uno dall altro, controllandone la velocità di riassorbimento. Lungo i tubuli renali viene riassorbita la maggior parte dei soluti e il
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido
 Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
FISIOLOGIA 9 CFU. 6 ore settimanali. Esame orale. Lun Mar Gio AULA I
 FISIOLOGIA 9 CFU 6 ore settimanali Esame orale Lun 14.30-16.30 Mar 11.30-13.30 Gio 11.30-13.30 AULA I La membrana plasmatica o cellulare avvolge ogni cellula e ne definisce l entità,separando il suo
FISIOLOGIA 9 CFU 6 ore settimanali Esame orale Lun 14.30-16.30 Mar 11.30-13.30 Gio 11.30-13.30 AULA I La membrana plasmatica o cellulare avvolge ogni cellula e ne definisce l entità,separando il suo
La pompa sodio-potassio
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 La pompa sodio-potassio 14/3/2006 Efficienza del cuore Dipende dall ordinata sequenza di eccitazione e di contrazione che procede
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni
 5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
5.10 Grazie alle proteine, la membrana plasmatica svolge molteplici funzioni Molte proteine della membrana plasmatica sono enzimi appartenenti a squadre di catalizzatori che agiscono nella catena di montaggio
LIPIDI E MEMBRANE BIOLOGICHE
 Cap.8 LIPIDI E MEMBRANE BIOLOGICE Definizione (la parola deriva dal greco lípos, grasso): MOLECOLE INSOLUBILI IN ACQUA SOLUBILI NEI SOLVENTI ORGANICI NON POLARI Molecole con struttura chimica e funzioni
Cap.8 LIPIDI E MEMBRANE BIOLOGICE Definizione (la parola deriva dal greco lípos, grasso): MOLECOLE INSOLUBILI IN ACQUA SOLUBILI NEI SOLVENTI ORGANICI NON POLARI Molecole con struttura chimica e funzioni
Le proteine si membrana si possono associare al doppio strato lipidico con modalità differenti
 Le proteine si membrana si possono associare al doppio strato lipidico con modalità differenti Proteine integrali di membrana Proteine periferiche di membrana Le proteine integrali di membrana possono
Le proteine si membrana si possono associare al doppio strato lipidico con modalità differenti Proteine integrali di membrana Proteine periferiche di membrana Le proteine integrali di membrana possono
la cellula al lavoro: cenni di termodinamica, enzimi, struttura della membrana plasmatica e il trasporto di membrana
 la cellula al lavoro: cenni di termodinamica, enzimi, struttura della membrana plasmatica e il trasporto di membrana Forme di energia L energia è la capacità di trasferire calore o compiere un lavoro.
la cellula al lavoro: cenni di termodinamica, enzimi, struttura della membrana plasmatica e il trasporto di membrana Forme di energia L energia è la capacità di trasferire calore o compiere un lavoro.
Ruolo del calcio nella cellula
 Ruolo del calcio nella cellula I meccanismi evolutivi hanno attribuito agli ioni fosfato un importanza fondamentale poiché sono necessari per sintetizzare ATP. Per questo motivo la loro concentrazione
Ruolo del calcio nella cellula I meccanismi evolutivi hanno attribuito agli ioni fosfato un importanza fondamentale poiché sono necessari per sintetizzare ATP. Per questo motivo la loro concentrazione
Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
, si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.") Membrana cellulare Proteine inserite nella membrana cellulare: Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
Membrana cellulare Proteine inserite nella membrana cellulare: Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
Riassorbimento dell acqua lungo il nefrone
 Riassorbimento dell acqua lungo il nefrone La quota escreta di acqua varia fra 0.25% (0.5 L/giorno, massima antidiuresi) ed il 13% (24 L/giorno, massima diuresi), attestandosi in condizioni normali intorno
Riassorbimento dell acqua lungo il nefrone La quota escreta di acqua varia fra 0.25% (0.5 L/giorno, massima antidiuresi) ed il 13% (24 L/giorno, massima diuresi), attestandosi in condizioni normali intorno
Lezione 1: Atomi e molecole:
 Lezione 1: Atomi e molecole: La materia è costituita da elementi chimici in forma pura o in combinazioni dette composti. La vita richiede circa 25 elementi chimici. La struttura atomica determina il comportamento
Lezione 1: Atomi e molecole: La materia è costituita da elementi chimici in forma pura o in combinazioni dette composti. La vita richiede circa 25 elementi chimici. La struttura atomica determina il comportamento
CLASSIFICAZIONE O.T.I.L.Is. Lig
 GLI ENZIMI CHE COSA SONO? Sono delle proteine altamente specializzate con attività catalitica, accelerano le reazioni chimiche rimanendo inalterati al termine della reazione stessa. CLASSIFICAZIONE O.T.I.L.Is.
GLI ENZIMI CHE COSA SONO? Sono delle proteine altamente specializzate con attività catalitica, accelerano le reazioni chimiche rimanendo inalterati al termine della reazione stessa. CLASSIFICAZIONE O.T.I.L.Is.
struttura e composizione della membrana cellulare Prof. Davide Cervia - Fisiologia Fisiologia della cellula: trasporti di membrana
 struttura e composizione della membrana cellulare 1 funzioni: separazione, regolazione, comunicazione, stabilizzazione modello a mosaico liquido 2 lipidi di membrana 3 4 Glicoproteine Glicolipidi Glicocalice
struttura e composizione della membrana cellulare 1 funzioni: separazione, regolazione, comunicazione, stabilizzazione modello a mosaico liquido 2 lipidi di membrana 3 4 Glicoproteine Glicolipidi Glicocalice
Riassorbimento tubulare
 Riassorbimento tubulare Riassorbimento Secrezione Qtr = Qf- Qe Qts = Qe Qf Qtr = VFG*P UV Qts = UV VFG*P Le urine si formano mediante tre processi: Filtrazione glomerulare Riassorbimento di acqua e soluti
Riassorbimento tubulare Riassorbimento Secrezione Qtr = Qf- Qe Qts = Qe Qf Qtr = VFG*P UV Qts = UV VFG*P Le urine si formano mediante tre processi: Filtrazione glomerulare Riassorbimento di acqua e soluti
ENZIMI. Un enzima è un catalizzatore (acceleratore) di reazioni biologiche.
 di reazioni biologiche.") ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
La compartimentalizzazione cellulare
 SISTEMA DI MEMBRANE La compartimentalizzazione cellulare LE MEMBRANE CELLULARI: UN SISTEMA IN EQUILIBRIO FUNZIONI DELLE MEMBRANE - delimitano i contorni cellulari o degli orgnelli intracellulari - consentono
SISTEMA DI MEMBRANE La compartimentalizzazione cellulare LE MEMBRANE CELLULARI: UN SISTEMA IN EQUILIBRIO FUNZIONI DELLE MEMBRANE - delimitano i contorni cellulari o degli orgnelli intracellulari - consentono
Tutta la vita cellulare ha le seguenti caratteristiche in comune. tutte le cellule hanno una membrana cellulare che separa il liquido extracellulare
 Tutta la vita cellulare ha le seguenti caratteristiche in comune. tutte le cellule hanno una membrana cellulare che separa il liquido extracellulare dal citoplasma cellulare che ha un alto grado di organizzazione.
Tutta la vita cellulare ha le seguenti caratteristiche in comune. tutte le cellule hanno una membrana cellulare che separa il liquido extracellulare dal citoplasma cellulare che ha un alto grado di organizzazione.
LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.
 LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.il modello a mosaico fluido descrive la struttura comune a tutte
LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.il modello a mosaico fluido descrive la struttura comune a tutte
Biologia generale Robert J. Brooker, Eric P. Widmaier, Linda E. Graham, Peter D. Stiling Copyright 2009 The McGraw-Hill Companies srl
 RISPOSTE AGLI SPUNTI DI RIFLESSIONE Figura 5.4 Capitolo 5 RISPOSTA : La bassa temperatura impedisce la diffusione laterale delle proteine di membrane. Quindi, dopo la fusione, tutte le proteine di topo
RISPOSTE AGLI SPUNTI DI RIFLESSIONE Figura 5.4 Capitolo 5 RISPOSTA : La bassa temperatura impedisce la diffusione laterale delle proteine di membrane. Quindi, dopo la fusione, tutte le proteine di topo
Le membrane cellulari La componente proteica: Diffusione delle proteine nella membrana
 Le membrane cellulari La componente proteica: Diffusione delle proteine nella membrana Le proteine diffondono nel piano della membrana per diffusione rotazionale e laterale Le membrane cellulari La componente
Le membrane cellulari La componente proteica: Diffusione delle proteine nella membrana Le proteine diffondono nel piano della membrana per diffusione rotazionale e laterale Le membrane cellulari La componente
Trasporto epiteliale
 Trasporto epiteliale Rivestono le cavità e le superfici corporee: interfaccia tra gli spazi interni dell organismo e l ambiente Semplice rivestimento o funzione di regolazione (es:osmoregolazione) Trasporto
Trasporto epiteliale Rivestono le cavità e le superfici corporee: interfaccia tra gli spazi interni dell organismo e l ambiente Semplice rivestimento o funzione di regolazione (es:osmoregolazione) Trasporto
Escrezione renale di Na + e sua regolazione
 Escrezione renale di e sua regolazione L escrezione di dipende dal carico filtrato (e quindi dalla VFG) e dal carico riassorbito Il riassorbimento di avviene in tutti i segmenti del nefrone e nel nefrone
Escrezione renale di e sua regolazione L escrezione di dipende dal carico filtrato (e quindi dalla VFG) e dal carico riassorbito Il riassorbimento di avviene in tutti i segmenti del nefrone e nel nefrone
TRASPORTO: qualunque tipo di passaggio di sostanze attraverso la membrana cellulare.
 Membrana citoplasmatica èsedediscambiodi materia, energia e informazione con l ambiente esterno TRASPORTO: qualunque tipo di passaggio di sostanze attraverso la membrana cellulare. Scaricato da www.sunhope.it
Membrana citoplasmatica èsedediscambiodi materia, energia e informazione con l ambiente esterno TRASPORTO: qualunque tipo di passaggio di sostanze attraverso la membrana cellulare. Scaricato da www.sunhope.it
Che ruolo ha la membrana plasmatica nei potenziali di diffusione?
 Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
Le cellule nervose e muscolari generano segnali elettrici principalmente mediante cambiamenti della permeabilità di membrana verso gli ioni sodio e potassio. Gli aumenti di permeabilità sono dovuti alla
TRASDUZIONE DEL SEGNALE CELLULARE
 TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
Struttura e funzione delle membrane biologiche
 La membrana plasmatica delimita la cellula e separa l ambiente interno da quello esterno. Non impedisce però tutti gli scambi Struttura e funzione delle membrane biologiche Figure'11)1'!Essen&al!Cell!Biology!(
La membrana plasmatica delimita la cellula e separa l ambiente interno da quello esterno. Non impedisce però tutti gli scambi Struttura e funzione delle membrane biologiche Figure'11)1'!Essen&al!Cell!Biology!(
Comunicazione tra cellula ed ambiente. FGE aa
 Comunicazione tra cellula ed ambiente FGE aa.2016-17 Fisiologia Umana La fisiologia è la scienza che studia i meccanismi di funzionamento degli organismi viventi. E una scienza integrata che utilizza principi
Comunicazione tra cellula ed ambiente FGE aa.2016-17 Fisiologia Umana La fisiologia è la scienza che studia i meccanismi di funzionamento degli organismi viventi. E una scienza integrata che utilizza principi
POTENZIALE DI MEMBRANA
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2016-17 POTENZIALE DI MEMBRANA Prof. Clara Iannuzzi Dipartimento di Biochimica, Biofisica e Patologia Generale clara.iannuzzi@unina2.it
FISIOLOGIA La fisiologia è una scienza che studia le funzioni degli organismi viventi
 FISIOLOGIA La fisiologia è una scienza che studia le funzioni degli organismi viventi 生理学 Obiettivi Al termine del corso di fisiologia lo studente dovrebbe conoscere i principi molecolari di base che regolano
FISIOLOGIA La fisiologia è una scienza che studia le funzioni degli organismi viventi 生理学 Obiettivi Al termine del corso di fisiologia lo studente dovrebbe conoscere i principi molecolari di base che regolano
TRASPORTO TRANSMEMBRANA Trasporto passivo: diffusione semplice e canali per ioni
 TRASPORTO TRANSMEMBRANA Trasporto passivo: diffusione semplice e canali per ioni Diffusione semplice: molecole lipofile attraversano la membrana spontaneamente Diffusione facilitata: i canali per ioni
TRASPORTO TRANSMEMBRANA Trasporto passivo: diffusione semplice e canali per ioni Diffusione semplice: molecole lipofile attraversano la membrana spontaneamente Diffusione facilitata: i canali per ioni
Capitolo 5 L energia e il trasporto
 Capitolo 5 L energia e il trasporto La cellula e l energia 5.1 L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità
Capitolo 5 L energia e il trasporto La cellula e l energia 5.1 L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità
Regolazione enzimatica Isoenzimi
 Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
materiale didattico, vietata la riproduzione e la vendita 1
 1 I lisosomi hanno funzione di sistema digestivo della cellula e degradano sia materiale trasportato dall esterno della cellula tramite endocitosi e che componenti cellulari non più utili frutto di autofagocitosi.
1 I lisosomi hanno funzione di sistema digestivo della cellula e degradano sia materiale trasportato dall esterno della cellula tramite endocitosi e che componenti cellulari non più utili frutto di autofagocitosi.
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici STRUTTURA TERZIARIA La struttura tridimensionale
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici STRUTTURA TERZIARIA La struttura tridimensionale
movimento di molecole e ioni tra i diversi scomparti dei sistemi biologici
 movimento di molecole e ioni tra i diversi scomparti dei sistemi biologici membrana cellulare garantisce un ambiente fisicochimico altamente stabile metabolismo riproduzione sviluppo eucarioti sviluppo
movimento di molecole e ioni tra i diversi scomparti dei sistemi biologici membrana cellulare garantisce un ambiente fisicochimico altamente stabile metabolismo riproduzione sviluppo eucarioti sviluppo
Il livello cellulare di organizzazione: la struttura cellulare. Cattedra di Fisiologia Umana
 Il livello cellulare di organizzazione: la struttura cellulare Cattedra di Fisiologia Umana 1. La diversità delle cellule nel corpo umano Le cellule del corpo hanno molte differenti forme ed una varietà
Il livello cellulare di organizzazione: la struttura cellulare Cattedra di Fisiologia Umana 1. La diversità delle cellule nel corpo umano Le cellule del corpo hanno molte differenti forme ed una varietà
STRUTTURA E FUNZIONI
 STRUTTURA E FUNZIONI Typical Cell Membrana plasmatica Chiamata anche membrana cellulare Circonda ogni cellula Separa il contenuto cellulare dal cio che la circonda Separa il LIC dal LEC Controlla il movimento
STRUTTURA E FUNZIONI Typical Cell Membrana plasmatica Chiamata anche membrana cellulare Circonda ogni cellula Separa il contenuto cellulare dal cio che la circonda Separa il LIC dal LEC Controlla il movimento
Farmacocinetica. Lo studio di questi quattro processi viene definito come FARMACOCINETICA del farmaco
 Farmacocinetica Affinché un farmaco possa raggiungere il sito d azione ad una concentrazione sufficiente per esercitare il proprio effetto terapeutico specifico deve generalmente essere: assorbito dal
Farmacocinetica Affinché un farmaco possa raggiungere il sito d azione ad una concentrazione sufficiente per esercitare il proprio effetto terapeutico specifico deve generalmente essere: assorbito dal
FADH ADP + Pi ATP...29 AG ' = -30,5 kj/mol...29 Resa di ATP per l ossidazione completa del glucosio...31
 FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
POTENZIALE DI MEMBRANA
 POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
POTENZIALE DI MEMBRANA 1 POTENZIALE DI RIPOSO Tutte le cellule (non solo le cellule eccitabili) hanno un potenziale di riposo (resting): una carica elettrica attraverso la membrana plasmatica, con l interno
Cell Signaling. Qualsiasi tipo di comunicazione tra le cellule
 Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La
Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La
Le membrane cellulari
 Le membrane cellulari Tutte le cellule, procariote ed eucariote, sono delimitate da una membrana (MEMBRANA PLASMATICA) Molti organelli intracellulari degli eucarioti sono circoscritti da membrane (MEMBRANE
Le membrane cellulari Tutte le cellule, procariote ed eucariote, sono delimitate da una membrana (MEMBRANA PLASMATICA) Molti organelli intracellulari degli eucarioti sono circoscritti da membrane (MEMBRANE
Funzioni della membrana plasmatica
 TRASPORTO M.P. -Omeostasi cellulare Funzioni della membrana plasmatica Mantenimento della conc. intracell. di ioni e molecole entro valori corretti Scambi interno - esterno Non c è consumo E Consumo E
TRASPORTO M.P. -Omeostasi cellulare Funzioni della membrana plasmatica Mantenimento della conc. intracell. di ioni e molecole entro valori corretti Scambi interno - esterno Non c è consumo E Consumo E
Caratteristiche generali dei trasporti mediati
 Caratteristiche generali dei trasporti mediati SPECIFICITA SATURAZIONE COMPETIZIONE TRASPORTO MEDIATO PASSIVO secondo gradiente di concentrazione TRASPORTO MEDIATO ATTIVO contro gradiente di concentrazione
Caratteristiche generali dei trasporti mediati SPECIFICITA SATURAZIONE COMPETIZIONE TRASPORTO MEDIATO PASSIVO secondo gradiente di concentrazione TRASPORTO MEDIATO ATTIVO contro gradiente di concentrazione
Funzioni dei nucleotidi
 Funzioni dei nucleotidi monomeri degli acidi nucleici esempi di altre funzioni ATP: moneta energetica GTP: fonte di energia nella sintesi proteica camp: secondo messaggero nella trasduzione del segnale
Funzioni dei nucleotidi monomeri degli acidi nucleici esempi di altre funzioni ATP: moneta energetica GTP: fonte di energia nella sintesi proteica camp: secondo messaggero nella trasduzione del segnale
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Membrane biologiche. Francesca Anna Scaramuzzo, PhD
 Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica Organica e Biochimica Membrane biologiche Francesca Anna Scaramuzzo, PhD Dipartimento di Scienze di Base e Applicate per l Ingegneria
Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica Organica e Biochimica Membrane biologiche Francesca Anna Scaramuzzo, PhD Dipartimento di Scienze di Base e Applicate per l Ingegneria
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I I TRASPORTI ATTRAVERSO MEMBRANE BIOLOGICHE Movimenti di piccoli soluti attraverso le membrane 2 categorie generali Trasporto
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I I TRASPORTI ATTRAVERSO MEMBRANE BIOLOGICHE Movimenti di piccoli soluti attraverso le membrane 2 categorie generali Trasporto
1 La diffusione semplice. La cellula interagisce con l ambiente. La membrana plasmatica.
 La cellula interagisce con l ambiente. La membrana plasmatica. Bibliografia I colori della biologia Giusti Gatti- Anelli Ed. Pearson La membrana plasmatica delle cellule funziona come un filtro, selezionando
La cellula interagisce con l ambiente. La membrana plasmatica. Bibliografia I colori della biologia Giusti Gatti- Anelli Ed. Pearson La membrana plasmatica delle cellule funziona come un filtro, selezionando
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
BIOCHIMICA APPLICATA e CLINICA
 BIOCHIMICA APPLICATA e CLINICA Regolazione Enzimatica Biosegnalazione Regolazione Ormonale Specializzazioni metaboliche (Biochimica d Organo) Integrazione del metabolismo BIOCHIMICA APPLICATA e CLINICA
BIOCHIMICA APPLICATA e CLINICA Regolazione Enzimatica Biosegnalazione Regolazione Ormonale Specializzazioni metaboliche (Biochimica d Organo) Integrazione del metabolismo BIOCHIMICA APPLICATA e CLINICA
I materiali della vita
 I materiali della vita I componenti chimici dei viventi Il corpo dei viventi è formato da relativamente pochi elementi chimici e in percentuale diversa da quella del mondo non vivente. Le molecole dei
I materiali della vita I componenti chimici dei viventi Il corpo dei viventi è formato da relativamente pochi elementi chimici e in percentuale diversa da quella del mondo non vivente. Le molecole dei
Corso di Laurea Magistrale in Medicina e Chirurgia. Biofisica e Fisiologia I. Muscolo liscio
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
Immagini e concetti della biologia
 Sylvia S. Mader Immagini e concetti della biologia 2 A3 Le molecole biologiche 3 Il carbonio è l elemento di base delle biomolecole Una cellula batterica può contenere fino a 5000 tipi diversi di composti
Sylvia S. Mader Immagini e concetti della biologia 2 A3 Le molecole biologiche 3 Il carbonio è l elemento di base delle biomolecole Una cellula batterica può contenere fino a 5000 tipi diversi di composti
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.
 Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
Si è ipotizzato che il potenziale di membrana fosse un potenziale di Equilibrio del K descritto dall eq. di Nerst : Em= -RT/ZF 2.3log [K]i / [K]o Registrazione del potenziale di membrana Potenziale di
COMPARTIMENTI INTRACELLULARI
 COMPARTIMENTI INTRACELLULARI ogni organello è delimitato da una, o due membrane: ciascuna di esse è costituita da un doppio strato fosfolipidico con stessa struttura della membrana plasmatica ma composizione
COMPARTIMENTI INTRACELLULARI ogni organello è delimitato da una, o due membrane: ciascuna di esse è costituita da un doppio strato fosfolipidico con stessa struttura della membrana plasmatica ma composizione
Fondamenti di Chimica Farmaceutica. Recettori
 Fondamenti di Chimica Farmaceutica Recettori Recettori In biochimica, un recettore è una proteina, transmembrana o intracellulare, che si lega ad uno specifico ligando, causando nel recettore una variazione
Fondamenti di Chimica Farmaceutica Recettori Recettori In biochimica, un recettore è una proteina, transmembrana o intracellulare, che si lega ad uno specifico ligando, causando nel recettore una variazione
Cono di crescita di un assone
 Cono di crescita di un assone cono di crescita assone Mentre il corpo di un assone mostra pochi segni esterni di attività motoria, la punta, o cono di crescita, assomiglia ad un fibroblasto strisciante,
Cono di crescita di un assone cono di crescita assone Mentre il corpo di un assone mostra pochi segni esterni di attività motoria, la punta, o cono di crescita, assomiglia ad un fibroblasto strisciante,
Rene ed equilibrio acido-base
 Rene ed equilibrio acidobase Produzione H Acidi volatili (15.000 mmol/dì) CO 2 H 2 O H 2 CO 3 H HCO 3 non costituisce guadagno H perché CO 2 (volatile) eliminata con la respirazione Acidi fissi, non volatili
Rene ed equilibrio acidobase Produzione H Acidi volatili (15.000 mmol/dì) CO 2 H 2 O H 2 CO 3 H HCO 3 non costituisce guadagno H perché CO 2 (volatile) eliminata con la respirazione Acidi fissi, non volatili
Alcune proteine vengono modificate nei dittiosomi.
 desmotubulo Reticolo endoplasmico Trasporto di macromolecole (proteine). E un sistema molto esteso di tubuli e sacculi che si ramificano e si interconnettono limitando uno spazio interno detto lume le
desmotubulo Reticolo endoplasmico Trasporto di macromolecole (proteine). E un sistema molto esteso di tubuli e sacculi che si ramificano e si interconnettono limitando uno spazio interno detto lume le
FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua
 FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua Fisiologia Generale e dell Esercizio Carlo Capelli Facoltà di Scienze Motorie Università degli Studi Verona Obiettivi Funzioni
FISIOLOGIA RENALE 2. Funzioni tubulari, riassorbimento obbligatorio dell acqua Fisiologia Generale e dell Esercizio Carlo Capelli Facoltà di Scienze Motorie Università degli Studi Verona Obiettivi Funzioni
Struttura delle membrane biologiche
 Struttura delle membrane biologiche Struttura sovramolecolare Pellicola oleosa sottilissima oligosaccaride fosfolipidi colesterolo proteina intrinseca Prof.ssa Flavia Frabetti 3 componenti la membrana
Struttura delle membrane biologiche Struttura sovramolecolare Pellicola oleosa sottilissima oligosaccaride fosfolipidi colesterolo proteina intrinseca Prof.ssa Flavia Frabetti 3 componenti la membrana
Movimenti dell acqua e dei soluti nella cellula vegetale
 Movimenti dell acqua e dei soluti nella cellula vegetale L acqua si sposta in funzione dell energia potenziale in essa contenuta o Potenziale idrico Si sposta da una regione dove il potenziale idrico è
Movimenti dell acqua e dei soluti nella cellula vegetale L acqua si sposta in funzione dell energia potenziale in essa contenuta o Potenziale idrico Si sposta da una regione dove il potenziale idrico è