Endomembrane e Smistamento delle proteine
|
|
|
- Michelina Foti
- 5 anni fa
- Visualizzazioni
Transcript

1 Endomembrane e Smistamento delle proteine La traduzione degli mrna inizia sempre nel citoplasma ma in dipendenza di specifici segnali può continuare e ultimare nel citoplasma stesso o sui ribosomi legati al Reticolo Endoplasmatico. Sequenze segnale Lo smistamento delle proteine ai comparti cellulari dipende da specifiche sequenze di amminoacidi presenti sulla proteina stessa. Tali sequenze sono dette sequenze segnale. Pertanto, il destino della proteina è noto già al momento della trascrizione del gene corrispondente
2 Struttura delle sequenze segnale La sequenza (peptidica) segnale può essere presente: alla estremità amino-terminale come ad esempio la sequenza per l importo al RER; alla estremità carbossi-terminale come ad esempio la sequenza per l importo nei perossisomi;intersperse come ad esempio nell importo o esporto nucleare. La presenza delle sequenze segnali prevede che esse siano riconosciute da specifiche proteine (recettori di smistamento) che le indirizzano al compartimento bersaglio; qui si legano a specifiche proteine e/o a canali di traslocazione presenti sulla membrana del comparto bersaglio in modo da consentire l inserimento della proteina o all interno del comparto o alla sua membrana
3 Smistamento proteine: importazione post-traduzionale le proteine sintetizzate in dipendenza di specifici segnali possono: restare nel citoplasma; oppure essere trasferite: nel nucleo; nei mitocondri; nei plastidi; nei perossisomi. Il trasferimento delle proteine in tali comparti avviene dopo la traduzione ed è pertanto detta: importazione posttraduzionale.

4 Tre proteine traslocatrici mitocondriali
5 Smistamento di proteine mitocondriali Se viene traslocata nella matrice il peptide segnale si lega ad un recettore sulla membrana mitocondriale esterna e la proteina viene traslocata grazie alla presenza di un secondo traslocatore sulla membrana interna: si crea un punto di contatto tra le due membrane. Le proteine sono traslocate non ripiegate. Intervengono in questo processo le proteine chaperonine, come hsp70, che mantengono le proteine srotolate. L idrolisi dell ATP viene usata per staccare l hsp70 e fornire energia motrice per importare la proteina attraverso il traslocatore.

6 Importazione di proteine nei mitocondri, la sequenza segnale è riconosciuta dalla proteina TOM.
 l hsp mitocondriale si lega alla catena polipetidica in regioni strategiche trascinandola Nella Matrice ed è poi rimossa grazie ad")
7 Ruolo dell energia nell importazione di proteine nella matrice mitocondriale 1) La proteina hsp70 citosolica attaccata è rilasciata dalla proteina in un passaggio che Dipende dall idrolisi di ATP 2) Traslocazione nella matrice grazie ad un gradiente protonico 3) l hsp mitocondriale si lega alla catena polipetidica in regioni strategiche trascinandola Nella Matrice ed è poi rimossa grazie ad idrolisi di ATP

8 La traslocazione delle proteine nella membrana mitocondriale interna e nello spazio intermembra na richiede: la sequenza segnale una sequenza di aa idrofobica dopo il peptide segnale, che funziona da nuovo peptide segnale quando il primo è stato tagliato via dalla peptidasi (meccanismo simile alla traslocazione nell ER)

9 Endomembrane della cellula eucariotica
10 Membrane biologiche e loro interazioni

11 Nucleo Poro Nucleare Reticolo Endoplasmatico

12 Traffico di Materiale attraverso I Pori Nucleari

13

14 Organizzazione del poro nucleare

15

16




17

18 Meccanismo di trasporto attivo attraverso i pori nucleari

: i tratti del RE che portano i ribosomi legati sul versante citoplasmatico.")
19 Reticolo Reticolo endoplasmatico Il Reticolo Endoplasmatico (RE) è presente in tutte le cellule eucariotiche. E una intricata rete di cisterne e tubuli intercomunicanti tra loro. Endoplasmatico E compartimento unico con porzioni morfologicamente e funzionalmente differenti, distinte in: Reticolo endoplasmatico Rugoso (RER): i tratti del RE che portano i ribosomi legati sul versante citoplasmatico. Reticolo endoplasmatico liscio (REL): i tratti di RE privi di ribosomi.
.")
20 Reticolo endoplasmatico Rugoso struttura Il Reticolo Endoplasmatico Rugoso è costituito da cisterne appiattite intercomunicanti, con i ribosomi legati alla superficie citosolica. Il RER è molto sviluppato nelle cellule che attivamente secernono proteine o glicoproteine (ad esempio i neuroni, le cellule del pancreas esocrino, le plasmacellule). Reticolo endoplasmatico Liscio struttura Il reticolo endoplasmatico liscio (REL o SER da Smooth Endoplasmatic Reticulum) si differenzia dal RER soprattutto per la mancanza di ribosomi sul versante citosolico. E costituito essenzialmente da tubuli interconnessi tra loro. Il REL è molto sviluppato nelle cellule steroidogenetiche, quali ad esempio le cellule di Leydig o quelle mineralcoticoidi del surrene, nelle fibre muscolari scheletriche dove è denominato Reticolo sarcoplasmatico

21

22 Microfotografia al microscopio elettronico
23 Reticolo Endoplasmatico Liscio (SER): funzioni Il REL assolve a numerose funzioni; le principali sono di seguito elencate: Biosintesi delle membrane. Partecipa alla biosintesi degli ormoni stereoidei. Metabolismo dei carboidrati. Immagazzinamento del Calcio. Detossificazione dei farmaci.
24 IL SER E RESPONSABILE DELLA SINTESI DI TRIACILGLICEROLI CHE VENGONO ACCUMULATI NEL LUME,CON PRODUZIONE DI GOCCIOLINE LIPIDICHE. NEGLI ADIPOCITI LE GOCCIOLINE LIPIDICHE OCCUPANO QUASI TUTTO LO SPAZIO DEL CITOPLASMA IL SER SINTETIZZA I FOSFOLIPIDI DELLE MEMBRANE CELLULARI IL FOSFOLIPIDE PRINCIPALE PRODOTTO E FOSFATIDILCOLINA O LECITINA PUO ESSERE FORMATO IN TRE PASSAGGI DA COLINA, DUE ACIDI GRASSI E GLICEROLO FOSFATO SER: sintesi di membrane Nel RE gli enzimi della sintesi dei fosfolipidi hanno il sito attivo sul lato citoplasmatico. Durante la sintesi si ha pertanto la crescita asimmetrica della membrana plasmatica (sintesi di un monostrato). Particolari enzimi (le scramblasi- dall inglese to scramble = mescolare) operano il mescolamento casuale dei fosfolipidi, ripristinando le dimensioni corrette dei due foglietti. Successivamente, altri enzimi ( le flippasi) operano il trasferimento selettivo dei fosfolipidi da un monostrato all altro, ripristinando l asimmetria, la composizione chimica e l organizzazione del doppio starto lipidico.
25 IL SER PARTECIPA ALLA SINTESI DI STEROLI COME IL COLESTEROLO IL COLESTEROLO VIENE SINTETIZZATO ATTRAVERSO UNA VIA METABOLICA A PIU STADI ALLA QUALE PARTECIPANO ANCHE ENZIMI PRESENTI NEL CITOSOL, OLTRE A QUELLI DEL SER LA SPECIE UMANA SINTETIZZA UNA GRAN PARTE DEL SUO COLESTEROLO TUTTE LE CELLULE SONO CAPACI DI PRODURLO, MA LA MAGGIOR PARTE DI ESSO VIENE SINTETIZZATO DALLE CELLULE EPATICHE IL FEGATO UTILIZZA IL COLESTEROLO PER FORMARE GLI ACIDI BILIARI
26 ORMONI STEROIDEI NELLA CORTECCIA SURRENALE E NELLE GONADI (REGIONE INTERSTIZIALE DEL TESTICOLO) IL COLESTEROLO E UTILIZZATO NELLA PRODUZIONE DEGLI ORMONI STEROIDEI NEI MITOCONDRI VIENE SCISSA LA CATENA LATERALE DEL COLESTEROLO, IMPORTATO DAL CITOSOL IL PRODOTTO COSI FORMATO LASCIA I MITOCONDRI E PASSA NEL SER DOVE VIENE DI NUOVO MODIFICATO. INFINE, TALE PRODOTTO DAL SER TORNA DI NUOVO NEI MITOCONDRI PER LE M0DIFICAZIONI FINALI COSI QUESTI DUE ORGANELLI GIOCANO UNA SORTA DI PALLA A VOLO PER PRODURRE L ORMONE STEROIDEO

27
28 IL SER CONTIENE UN ENZIMA GLUCOSIO-6-FOSFATASI G6Pase ESSENZIALE PER L OMEOSTASI DEL GLUCOSIO NEL SANGUE, L ENZIMA E PRESENTE SOLO NELLE CELLULE PARENCHIMALI DEL FEGATO
29 LA DETOSSIFICAZIONE E SVOLTA DA UN SISTEMA DI ENZIMI CHE TRASFERISCONO L OSSIGENO (MONOSSIGENASI) E IL CUI COMPONENTE PRINCIPALE E IL CITOCROMO P-450 IL SISTEMA DELLE MONOSSIGENASI P-450 E FORMATO DA NUMEROSI ISOENZIMI (50-100) DEL CITOCROMO P-450 (EMO-PROTEINA DI TIPO b), DA UNA SINGOLA REDUTTASI E FOSFOLIPIDE.

30 QUESTI ENZIMI SONO PROTEINE INTEGRALI DI MEMBRANA E SONO PREMINENTI NEL SER, MA PRESENTI ANCHE NEI MITOCONDRI IL CITOCROMO P-450 E UBIQUITARIO, MA SI TROVA IN CONCENTRAZIONI MOLTO ELEVATE NEL SER DELLE CELLULE EPATICHE RAPPRESENTA CIRCA IL 20% DELLE PROTEINE DEL ER E IL 2-3% DELLE PROTEINE TOTALI DELLA CELLULA
31 QUESTI ENZIMI SONO CAPACI DI OSSIDARE MIGLIAIA DI COMPOSTI IDROFOBICI DIFFERENTI CHE CONVERTONO IN DERIVATI PIU IDROFILICI E, QUINDI, PIU FACILMENTE SECRETI I SUBSTRATI DI TALI ENZIMI SONO IN GRADO DI INDURRE PROLIFERAZIONE DEL SER E CONSEGUENTE AUMENTO DELL ATTIVITA DEGLI ENZIMI SONNIFERI A BASE DI FENOBARBITAL (C OSI COME CENTINAIA DI ALTRE SOSTANZE CHIMICHE) INDUCONO PROLIFERAZIONE DEL SER E AUMENTO DELL ATTIVITA DELLE MONOSSIDASI NELLE CELLULE EPATICHE IL CONSEGUENTE AUMENTO DELLA CAPACITA DI DEGRADARE IL FENOBARBITAL SPIEGA PER QUALE MOTIVO I CONSUMATORI DI SONNIFERO DEVONO ASSUMERNE DOSI SEMPRE PIU ELEVATE
32 PERTANTO L ASSUNZIONE CRONICA DI FENOBARBITAL (O ALTRE SOSTANZE) AUMENTA L EFFICIENZA DI DEGRADAZIONE, DA PARTE DEL FEGATO, DI MOLTI ALTRI FARMACI, TRA CUI SOSTANZE TERAPEUTICAMENTE UTILI, COME ANTIBIOTICI, STEROIDI, ANTICOAGULANTI E NARCOTICI COMUNQUE L AUMENTATO SVILUPPO DEL SER NON COMPORTA AUMENTO EQUIVALENTE DI TUTTI GLI ENZIMI MENTRE L AZIONE DEL COMPLESSO P-450 E NEL COMPLESSO FAVOREVOLE DAL PUNTO DI VISTA FISIOLOGICO, CERTI EVENTI METABOLICI BASATI SUL P- 450 POSSONO PORTARE A SERIE PATOLOGIE

33 Reticolo endoplasmatico Rugoso funzioni: sintesi proteine Sul RER vengono sintetizzate le proteine che penetrate nel lume del RER, passano al Golgi e da qui smistate alla membrana plasmatica, ai lisosomi e alla vescicola di secrezione. L importazione delle proteine nel lume del RER è co-traduzionale.
.")
34 Nel caso che il peptide nascente presenti una particolare sequenza di amminoacidi, detta peptide segnale, la sintesi proteica è momentaneamente bloccata per continuare dopo la smistamento dei polisomi sulla membrana del Reticolo Endoplasmatico (RE). Le proteine sintetizzate entrano lume del RE, da qui passano nell apparato di Golgi, per essere poi inviate: alla membrana plasmatica; a vescicole di secrezione; ai lisosomi. Lo smistamento delle proteine al RE avviene durante la traduzione ed è, pertanto, detta: importazione co-traduzionale.

35 legame tra il peptide segnale e la proteina solubile SRP con blocco della sintesi proteica; legame tra il complesso mrna-ribosomi-srp e il recettore dell SRP (una proteina transmembrana del RE); legame del ribosoma al traslocone e distacco della SRP dal suo recettore; ripresa del sintesi della proteina che tramite il canale del traslocone penetra nel lume del RER; la peptidasi del segnale taglia il peptide segnale dalla proteina; rilascio della proteina nel lume del RER.


36 RER funzioni: sintesi proteine transmembrana Le proteine sintetizzate sul RER in dipendenza di assenza o presenza nelle proteine in sintesi di una specifica sequenza di arresto di trasferimento possono sia passare nello spazio luminale o restare inserite nella membrana, ovvero diventare proteine transmembrana: A in assenza del sequenza di arresto di trasferimento, eliminato il peptide segnale la proteina passa nel lume del RER; B in presenza del segnale di arresto di trasferimento (una serie di aminoacidi apolari), eliminato il peptide segnale, la proteina resta inserita nella membrana, come proteina transmembrana monopasso; C in presenza di due o più segnali di arresto di trasferimento, si avranno proteine transmembrana di-, tri- o multipasso

37 Le modalità di inserzione delle proteine nei bilayer fosfolipidici vengono già definite nel RER.

 al gruppo aminico delle asparagine presenti nella catena polipeptidica in crescita (la gicosilazione a livello delle asparagine è detta N- glicosilazione.")
38 Reticolo endoplasmatico Rugoso funzioni: glicosilazione Nel lume del RER inizia la glicosilazione delle proteine. Tale processo continuerà nell apparato di Golgi ed è importante per l indirizzamento delle proteine alla membrana plasmatica, ai lisosomi e alle vescicole di secrezione. La glicosilazione consiste nel trasferimento ad opera di un enzima (la oligosaccaridedolicol-transferase) di un oligosaccaride, di 14 residui zuccherini, ricco in mannosio, da un glicolipide di membrana (il dolicolo) al gruppo aminico delle asparagine presenti nella catena polipeptidica in crescita (la gicosilazione a livello delle asparagine è detta N- glicosilazione. Nel citoplasma la (eventuale) glicosilazione delle proteine avviene a livello del gruppo OH delle serine o delle treonine ed è, pertanto, detta O- glicosilazione).
.")

39 Reticolo endoplasmatico Rugoso e folding delle proteine Nel reticolo endoplasmatico inizia il processamento dell oligosaccaride aggiunto alle asparagine: ad opera di glicosidasi vengono eliminati quattro residui zuccherini (tre glucosio e un mannosio). Avviene anche il folding (ripiegamento) delle glico-proteine, tramite specifiche chaporonine, le bip. Le proteine correttamente folded vengono smistate all apparato di Golgi. Quelle non correttamente folded vengono traslocate nel citoplasma e degradate da specifici complessi enzimatici (i proteosomi).
 e intermedi di trasporto tubulo vescicolari.")
40 Nelle cellule eucariotiche è possibile individuare un sistema di membrane interne organizzate in un compartimento secretorio che permette a proteine, lipidi e carboidrati neosintetizzati di raggiungere la superficie cellulare. Questo sistema è costituito da organelli (reticolo endoplasmatico, apparato di Golgi e membrana plasmatica) e intermedi di trasporto tubulo vescicolari.

41 Apparato di Golgi L apparato del Golgi è costituito da una serie di cisterne (pile golgiane o, nelle cellule vegetali, dittiosiomi) ciascuna contenente un distintivo set di enzimi, in maggioranza glicosidasi. Le cisterne sono slargate in periferia, impilate l una sull altra e non intercomunicanti tra loro. Nelle pile golgiane si distinguono: faccia CIS o prossimale contigua al Reticolo endoplasmatico, da cui riceve le microvescicole, le quali fondono tra a costituire una rete di vescicole interconnesse detta CIS Golgi Network (CGN). faccia trans o distale lontana dal reticolo endoplasmatico Dalla cisterna si staccano vescicole,interconnesse tra loro a costituire il Trans Golgi Network (TGN), da cui si staccano vescicole che diventano lisosomi, oppure vescicole di secrezione, o che si fondono con la membrana plasmatica cisterne mediane Le cisterne interposte tra le CIS e TRANS.

42 Le cisterne golgiane differiscono sia strutturalemnte sia funzionalmente. Le membrane, infatti, presentano una differente composizione lipida e proteica che rendono ciascuna cisterna funzionalmente differenziata dalle altre. La componente proteica è costituita particolarmente da enzimi, glicosidasi e glicotrasferasi, che modificano l oligosaccaride, costituito da 10 residui zuccherini legati alle asparagine, che il Golgi riceve dal RE. Gli enzimi contenuti nelle cisterne conducono a due tipi due tipi di oligosaccaridi: oligosaccaridi complessi o oligosaccaridi ad alto contenuto di mannosio.

43 L apparato del Golgi riceve, elabora e smista ad altri compartimenti il materiale (le glicoproteine) che riceve dal Reticolo endoplasmatico tramite microvescicole di trasporto. Nel Golgi l elaborazione del materiale può avvenire secondo due modalità: progressione delle vesciole progressione delle cisterne Secondo il modello della progressione delle vescicole il materiale, elaborato in un cisterna, viene in incluso in microvescicole di trasporto che gemmano lateralmente dai margini della cisterna per fondersi con la membrana della cisterna immediatamente successiva. Il processo continua fino alla cisterna TRANS. Dal TRANS si staccano vescicole che possono costituire i lisosomi primari (1) oppure le vescicole di secrezione (2) o anche fondersi con la membrana plasmatica (3). Nel modello della progressione delle vescicole si ha il movimento delle vescicole in direzione Cis-Trans. Tale movimento costituisce il flusso vescicolare anterogrado.
, troppo grandi per essere contenute nelle micovescicole di trasporto, prevede che il materiale permanga nelle cisterne le quali avanzano maturando progressivamente in")
44 Apparato del Golgi traffico vescicolare- progressione delle cisterne Il modello della progressione delle cisterne, valido soprattutto per le molecole di notevole dimensioni (quali ad esempio la cellulosa, la lattoalbumina), troppo grandi per essere contenute nelle micovescicole di trasporto, prevede che il materiale permanga nelle cisterne le quali avanzano maturando progressivamente in Cis, Mediana e Trans. Lateralmente, dalle cisterne gemmano delle microvescicole che trasportano nella cisterna precedente gli enzimi caratteristici di quella cisterna. Il materiale lascia il Golgi tramite vescicole si staccano dal TGN per formare i lisosomi, oppure le vescicole di secrezione o anche fondersi con la membrana plasmatica. Secondo tale modello le cisterne, progredendo in direzione CIS-TRANS, realizzerebbero il Flusso delle cisterne anterogrado, mentre le vescicole, scorrendo nella direzione opposta TRANS-CIS, darebbero origine al flusso vescicolare retrogrado.

45 Proteine, lipidi e carboidrati neosintetizzati, sono smistati ai vari distretti cellulari da vescicole di Trasporto.
 che ne segnala il compartimento di destinazione.")
46 Indirizzamento delle vescicole di trasporto Le vescicole di trasporto presentano sulla membrana un particolare recettore per il materiale da trasportare. A ciascuno di tale recettore è associato dal lato citosolico una specifica proteina v-snare (vscicular-snare) che ne segnala il compartimento di destinazione. Su tale compartimento è presente una t-snare, che interagisce con la v-snare. Coppie specifiche di proteine transmembarna v-snare e t-snare determinano la destinazione delle vescicole.
 e t- SNARE (nella membrana del comparto")

47 Fusione di vescicole di trasporto La specifica segnalazione e fusione delle vescicole è medita dall interazione di coppie complementari di proteine SNAREs: v-snares (nella membrana della vescicola) e t- SNARE (nella membrana del comparto bersaglio). L adesione delle vescicole al comparto di destinazione è mediato dal legame tra le SNAREs, mentre la fusione delle vescicole e lo scarico del materiale nel comparto di destinazione è mediato da specifiche proteine di fusione (Rab e SNAPs,).

48 Le proteine Rab assicurano la specificitàdell ancoraggio. Si tratta di una famiglia di GTPasi monomeriche la cui funzione è di verificare che l adattamento tra una v- SNARE e una t-snare sia corretto. Quando una vescicola incontra la membrana bersaglio corretta il legame v-snare a t-snare fa rimanere la vescicola legata abbastanza a lungo in maniera che Rab possa idrolizzare il GTP legato e bloccare la vescicola. la fusione delle vescicole e lo scarico del materiale nel comparto di destinazione è mediato da specifiche proteine di fusione (Rab e SNAPs,).

49 Ruolo delle proteine di fusione nell internalizazione di virus

, il cui recettore Vescicole è presente di trasporto sulla membrana e proteine del associate RE.")
50 I vari tipi di proteine di rivestimento implicate nel traffico vescicolare tra il RE, l apparato del Golgi e la membrana plasmatica. La gemmazione delle vescicole dal Reticolo endoplasmatico è mediato da specifiche proteine di rivestimento le COP-II, COat Proteins type II. Una volta che la vescicola si stacca dal RE le COP-II ritornano al RE. Le vescicole arrivano al CIS- Golgi e scaricano il materiale. Dal CIS vengono riciclate al RE le vescicole che presentano sulla membrana una proteina con il segnale di ritenzione al RE. Tale segnale è costituito da 4 a.a. lisina, asparagina, glucina, leucina (K,D,E, L), il cui recettore Vescicole è presente di trasporto sulla membrana e proteine del associate RE. La gemmazione di tali vescicole è mediata dalle proteine di rivestimento del tipo COP-I.

51 Trasporto selettivo mediato da vescicole rivestite Il trasporto vescicolare non è aspecifico: ciascuna vescicola di trasporto contiene proteine selezionate e si fonde con un compartimento specifico: questo permette di mantenere l identità di ogni tipo di organello. Via biosintetica-secretoria: trasporto all esterno di proteine di nuova sintesi: ER-Golgisuperficie cellulare (lisosomi) Via endocitica: assunzione di macromolecole, passaggio in endosomi e lisosomi.

52 Compartimentalizzazione funzionale del Golgi Le cisterne del Gogi presentano uno specifico set di enzimi che modificano le glicoproteine durante il loro transito nelle cisterne in direzione Cis-Trans. Nella immagine a fianco sono riportate le principali modifiche che in ciascuna cistena possono essere apportate ai residui oligosaccaridici. In dipendenza delle modifiche apportate ai residui oligasaccaridi le glicoproteine vengono indirizzate ai lisosomi o alle vescicole di secrezione o alla membrana plasmatica.
53 lisosomi I lisosomi furono scoperti dapprima biochimicamente da De Duve su celllule di ratto, da cui riuscì ad isolare ed purificare i lisosomi attraverso delle semplici tecniche di centrifugazione. De Duve osservò che nella frazione lisosomiale l attività delle idrolasi acide da lui studiati (fosfatasi acida, DNasi, RNAsi, glucosidasi, ect) era molto alta nel sedimento e bassa nel supernatante quando utilizzava metodi di isolamento dolci (preservanti le membrane cellulari). Tuttavia, dopo il congelamento della frazione lisosomiale o il suo isolamento con metodi drastici (destabilizzanti le membrane cellulari) l attività delle idrolasi risultava alta nel supertante e bassa nel sedimento. Da tali esperimenti De Duve dedusse che le idrolasi acide dovevano essere contenute in organelli delimitati membrana che furono denominati lisosomi in base alle definizione data da Duve a tali organelli: This Small Particle acts as the digestive tract of the living cell. Its enzymes dissolve the substances ingested by the cell and under certain circumstances can dissolve the cell itself.
54 I lisosomi: punti di incontro ove convergono le sostanze che devono essere digerite. Enzimi lisosomali: idrolasi lisosomali Alcune proteine sono trasportate nel lisosoma direttamente con l uso di un segnale lisosomale: KFERQ (Lys, Phe, Glut, Arg, glutammina) Il modello proposto per la maturazione e smistamento delle proteine lisosomiali prevede che esse siano sintetizzate e traslocate nel lume del RER, dove avviene la glicosilazione delle asparagine. Le glicoproteine poi passano nella cisterna Cis dell apparato del Golgi. In tale cisterna si ha la sforilazione dei residui del mannosio (M-6-P). Le proteine così marcate, senza subire altre modifiche, arrivano al TGN, dove sono riconosciute dal recettore del M-6-P ed incluse in vescicole rivestite da clatrina. Le vescicole successivamente trasportano le proteine lisosomiali ad un endosoma tardivo da cui gemmano le vescicole che riciclano al TGN i recettori del M-6-P.
 Fagocitosi Endosoma precoci e")
55 Vie per arrivare al lisosoma: Autofagia (ER si trasforma in lisosoma) Fagocitosi Endosoma precoci e tardivi

56 Le idrolasi lisosomiali sono caratterizzate dalla presenza di un gruppo specifico, il mannosio- 6-fosfato (M6P), che serve a farle riconoscere da un recettore specifico presente sul trans- Golgi. L M6P è aggiunto agli oligosaccaridi legati all asparagina, nel reticolo cis del Golgi. Il recettore è una proteina transmembrana localizzata sul trans Golgi. L unione con l idrolasi inizia la formazione di vescicole e si verifica a ph 7. Le idrolasi si dissociano dai recettori negli endosomi tardivi a ph 6. I recettori sono riciclati verso il trans Golgi.
57 La GlcNAc-fosfotransferasi, l enzima che marca gli enzimi lisosomiali nel Golgi
58 Malattie da accumulo lisosomale: le idrolasi lisosomali sono alterate per difetti genetici recessivi. Malattia di Hurler I-cell disease I substrati delle idrolasi si accumulano non digeriti nel lisosomi, per cui si formano inclusioni nelle cellule. Le idrolasi alterate sfuggono ai lisosomi, sono secrete dalla via di default e si accumulano nel sangue: il difetto è dovuto alla mancanza o al difetto di una fosfotransferasi GlcNAc. Le idrolasi sono secrete. Non funziona la via di scavenger (raccolta dei rifiuti) di endocitosi e si accumulano. Nel fegato via alternativa.
59 Endocitosi: Dalla superficie cellulare verso i lisosomi. Fagocitosi di grandi particelle (250 nm) svolta da cellule specializzate tramite fagosomi. Le particelle si devono legare ai fagociti: recettori attivati scatenano la risposta (vd. Immunologia). Normalmente sono gli anticorpi che riconoscono le particelle estranee e le presentano ai fagociti. Proteine Fc o complemento.

60 Endocitosi: la fagocitosi è una forma specializzata nei macrofagi e neutrofili (10 all 11 globuli rossi al giorno)
61 Fosse rivestite di clatrina: 2% dell area di membrana La clatrina forma un cesto o una gabbia sotto la membrana e dà origine ad una vescicola rivestita di clatrina: è un processo rapido e continuo in molti tipi cellulari. Le vescicole si fondono poi negli endosomi precoci. Servono per l endocitosi in fase fluida e per l assunzione di specifiche macromolecole dall esterno. Questo processo è chiamato endocitosi mediata da recettori: processo selettivo che permette di assumere grandi quantità di macromolecole. Es. trasporto del colesterolo.
62 Le cellule assumono colesterolo che serve per assemblare le membrane. Se si blocca l assunzione, il colesterolo si accumula nel sangue e può formare placche aterosclerotiche. Il colesterolo è trasportato nel sangue associato a proteine LDL (lipoproteine a basse densità).
63 Ogni LDL contiene 1500 mol colesterolo esterificato ad acidi grassi
64 Le cellule presentano dei recettori per le LDL, che si associano alle zone di membrana ricche in clatrina. L LDL si lega al recettore e si formano vescicole rivestite che sono internalizzate. Le vescicole perdono il rivestimento in clatrina e si fondono negli endosomi precoci. Le LDL sono portate agli endosomi tardivi ed ai lisosomi, dove gli esteri di colesterolo sono idrolizzati ed il colesterolo liberato per l incorporazione nelle membrane. Se c è troppo colesterolo, viene spenta la sintesi di colesterolo e anche di recettori per LDL, per non importarlo più e così il colesterolo si accumula nel sangue. Geni difettosi per le LDL: mancata assunzione del colesterolo, aterosclerosi in età giovanile.

65 Struttura della clatrina

66

67 In alcune patologie i recettori per LDL sono situati al di fuori delle Recettori delle LDL normali e mutanti fossette rivestite da clatrina. Tale diversa localizzazione dei recettori per LDL impedisce la normale importazione di colesterolo.

68 Endocitosi di LDL mediata da recettori

69 I recettori per le LDL sono riciclati dagli endosomi e riportati in membrana per essere riutilizzato.

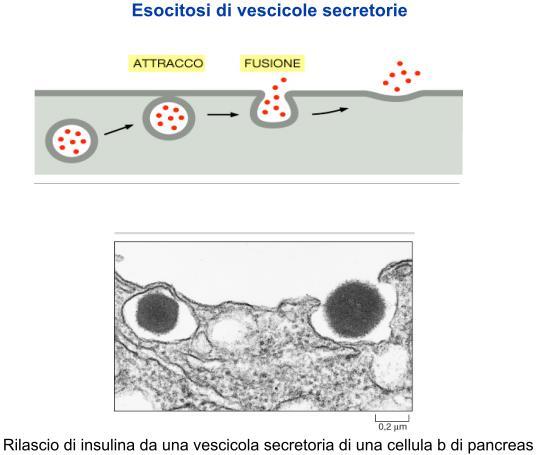
70 Esocitosi di vescicole secretorie
 nella matrice extracellulare le glicoproteine contenute nel lume.")
 plasmacellule (esocitosi costitutiva di immunoglobuline).")
71 Le proteine di secrezione vengono immesse in due tipi di vescicole di secrezione: - Vesciscole non rivestite da clatrina ma da coatomeri che ininterrottamente gemmano dal TGN, fondono con la membrana plasmatica e riversano (Esocitosi) nella matrice extracellulare le glicoproteine contenute nel lume. Tale modalità di esocitosi rappresenta la secrezione costitutiva che è generalizzata in tutti i tipi cellulari, ad esempio i fibroblasti (esocitosi costitutiva di fibre collagene, la fibronectina, GAG, ect) plasmacellule (esocitosi costitutiva di immunoglobuline). - Vescole rivestite da clatrina che si staccano dal TGN, si accumulano nel citoplasma e vengono successivamente rilasciate in seguito a specifici segnali provenienti dalla matrice extracellulare. Tale modalità di Esocitosi, denominata secrezione regolata, è tipica delle cellule ghiandolari endocrinie (secrezione regolata di ormoni) e soprattutto delle cellule degli epiteli ghiandolari esocrini ( esocitosi di secreti mucosi, sierosi, misti, ect). Da notare che la clatrina media anche la gemmazione delle vescicole contenenti le proteine lisosomiali.

72 Una via di secrezione
COMPARTIMENTI INTRACELLULARI
 COMPARTIMENTI INTRACELLULARI ogni organello è delimitato da una, o due membrane: ciascuna di esse è costituita da un doppio strato fosfolipidico con stessa struttura della membrana plasmatica ma composizione
COMPARTIMENTI INTRACELLULARI ogni organello è delimitato da una, o due membrane: ciascuna di esse è costituita da un doppio strato fosfolipidico con stessa struttura della membrana plasmatica ma composizione
PASSAGGIO DAL GOLGI E DESTINI
 PASSAGGIO DAL GOLGI E DESTINI Apparato di Golgi lisosomi Apparato di Golgi membrana plasmatica Apparato di Golgi secrezione Apparato di Golgi reticolo (trasporto retrogrado) lisosomi idrolasi acide qual
PASSAGGIO DAL GOLGI E DESTINI Apparato di Golgi lisosomi Apparato di Golgi membrana plasmatica Apparato di Golgi secrezione Apparato di Golgi reticolo (trasporto retrogrado) lisosomi idrolasi acide qual
I ribosomi liberi nel citoplasma sintetizzano le proteine destinate alla via citoplasmatica, cioè quelle destinate a:
 I ribosomi liberi nel citoplasma sintetizzano le proteine destinate alla via citoplasmatica, cioè quelle destinate a: filmato Rimanere nel citoplasma Essere trasportate dal citoplasma al nucleo Essere
I ribosomi liberi nel citoplasma sintetizzano le proteine destinate alla via citoplasmatica, cioè quelle destinate a: filmato Rimanere nel citoplasma Essere trasportate dal citoplasma al nucleo Essere
Dischi piatti (cisterne) impilate con una polarità. Microvescicole (trasporto) Macrovescicole (secrezione) Tre regioni: Cis Mediale Trans
 impilate con una polarità. Microvescicole (trasporto) Macrovescicole (secrezione) Tre regioni: Cis Mediale Trans") Apparato di Golgi Aspetto dell apparato di Golgi Dischi piatti (cisterne) impilate con una polarità. Microvescicole (trasporto) Macrovescicole (secrezione) Tre regioni: Cis Mediale Trans Funzioni dell
Apparato di Golgi Aspetto dell apparato di Golgi Dischi piatti (cisterne) impilate con una polarità. Microvescicole (trasporto) Macrovescicole (secrezione) Tre regioni: Cis Mediale Trans Funzioni dell
TRAFFICO INTRACELLULARE E COMUNICAZIONE TRA LE CELLULE
 TRAFFICO INTRACELLULARE E COMUNICAZIONE TRA LE CELLULE Carboidrati proteine Glicolipide Regione ad α-elica di proteina transmembrana Acido grasso Doppio strato Fosfolipidico 5-8 nm Proteina integrale di
TRAFFICO INTRACELLULARE E COMUNICAZIONE TRA LE CELLULE Carboidrati proteine Glicolipide Regione ad α-elica di proteina transmembrana Acido grasso Doppio strato Fosfolipidico 5-8 nm Proteina integrale di
8. Lo smistamento delle proteine
 8. Lo smistamento delle proteine contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale Tre sono i meccanismi con i quali le proteine
8. Lo smistamento delle proteine contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale Tre sono i meccanismi con i quali le proteine
FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE
 FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE Carboidrati proteine Glicolipide Regione ad a-elica di proteina transmembrana Acido grasso Doppio strato Fosfolipidico 5-8 nm Proteina integrale di membrana
FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE Carboidrati proteine Glicolipide Regione ad a-elica di proteina transmembrana Acido grasso Doppio strato Fosfolipidico 5-8 nm Proteina integrale di membrana
Incubazione PULSE-CHASE di pezzi di tessuto pancreatico in solux. contenente AA radioattivi. Gli AA venivano assimilati e incorporati
 Nobel nel 1974 a James JAMIESON e George PALADE Incubazione PULSE-CHASE di pezzi di tessuto pancreatico in solux. contenente AA radioattivi. Gli AA venivano assimilati e incorporati Reticolo endoplasmatico
Nobel nel 1974 a James JAMIESON e George PALADE Incubazione PULSE-CHASE di pezzi di tessuto pancreatico in solux. contenente AA radioattivi. Gli AA venivano assimilati e incorporati Reticolo endoplasmatico
Smistamento delle proteine neosintetizzate
 Smistamento delle proteine neosintetizzate Principi di Citologia e Istologia. Prof. Pucci, 2003 La sintesi continua sul RER La sintesi continua su ribosomi liberi Proteine citoplasmatiche Proteine lisosomali,
Smistamento delle proteine neosintetizzate Principi di Citologia e Istologia. Prof. Pucci, 2003 La sintesi continua sul RER La sintesi continua su ribosomi liberi Proteine citoplasmatiche Proteine lisosomali,
organismi pluricellulari Differenziamento compartimentalizzazione
 Negli organismi pluricellulari ogni unità partecipa all insieme, incapace di vita autonoma ma unità di base della struttura e funzione dell organismo. Gli organismi pluricellulari sono caratterizzati dalla
Negli organismi pluricellulari ogni unità partecipa all insieme, incapace di vita autonoma ma unità di base della struttura e funzione dell organismo. Gli organismi pluricellulari sono caratterizzati dalla
Le due subunità ribosomali sono formate nel nucleolo dall associazione tra rrna e proteine
 Le due subunità ribosomali sono formate nel nucleolo dall associazione tra rrna e proteine 5S 28S 18S 5.8S 60S 40S I poliribosomi Numerosi ribosomi possono legarsi in sequenza su una stessa molecola di
Le due subunità ribosomali sono formate nel nucleolo dall associazione tra rrna e proteine 5S 28S 18S 5.8S 60S 40S I poliribosomi Numerosi ribosomi possono legarsi in sequenza su una stessa molecola di
Smistamento delle proteine nella cellula
 SMISTAMENTO DELLE PROTEINE LE PROTEINE VENGONO SMISTATE A PEROSSISOMI, NUCLEO, MITOCONDRI E RE IN MODO INDIPENDENTE E AUTONOMO. DAL RE, INVECE, SI DIPARTE LA VIA DI SMISTAMENTO CHE PORTA LE PROTEINE A
SMISTAMENTO DELLE PROTEINE LE PROTEINE VENGONO SMISTATE A PEROSSISOMI, NUCLEO, MITOCONDRI E RE IN MODO INDIPENDENTE E AUTONOMO. DAL RE, INVECE, SI DIPARTE LA VIA DI SMISTAMENTO CHE PORTA LE PROTEINE A
MEMBRANE INTERNE. nm)
") MEMBRANE INTERNE Nella cellula eucariotica,, membrane circoscrivono cavità chiuse di varia forma: i compartimenti citoplasmatici. In base alla forma, le strutture delimitate da membrana possono essere
MEMBRANE INTERNE Nella cellula eucariotica,, membrane circoscrivono cavità chiuse di varia forma: i compartimenti citoplasmatici. In base alla forma, le strutture delimitate da membrana possono essere
TRASPORTO VESCICOLARE
 TRASPORTO VESCICOLARE ORIGINE Dalle membrane degli organelli DESTINAZIONE Verso altri organelli (trasporto intracellulare) Verso l ambiente extracellulare (esocitosi) Dalla membrana plasmatica Verso l
TRASPORTO VESCICOLARE ORIGINE Dalle membrane degli organelli DESTINAZIONE Verso altri organelli (trasporto intracellulare) Verso l ambiente extracellulare (esocitosi) Dalla membrana plasmatica Verso l
Biologia generale Prof.ssa Bernardo
 Cellula procariotica cellula eucariotica CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica,
Cellula procariotica cellula eucariotica CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica,
FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE
 FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE Meccanismi e vie dello smistamento di molecole Smistamento delle proteine nei compartimenti cellulari ed endocitosi Le vie principali La membrana nucleare Continuità
FUNZIONE CELLULARE E TRAFFICO INTRACELLULARE Meccanismi e vie dello smistamento di molecole Smistamento delle proteine nei compartimenti cellulari ed endocitosi Le vie principali La membrana nucleare Continuità
materiale didattico, vietata la riproduzione e la vendita 1
 1 I lisosomi hanno funzione di sistema digestivo della cellula e degradano sia materiale trasportato dall esterno della cellula tramite endocitosi e che componenti cellulari non più utili frutto di autofagocitosi.
1 I lisosomi hanno funzione di sistema digestivo della cellula e degradano sia materiale trasportato dall esterno della cellula tramite endocitosi e che componenti cellulari non più utili frutto di autofagocitosi.
Compartimenti intracellulari
 Compartimenti intracellulari Endosomi: smistamento di materiali assunti per endocitosi Perossisomi: sede di reazioni ossidative per la demolizione di lipidi e di molecole tossiche Lisosomi: contengono
Compartimenti intracellulari Endosomi: smistamento di materiali assunti per endocitosi Perossisomi: sede di reazioni ossidative per la demolizione di lipidi e di molecole tossiche Lisosomi: contengono
- In che modo questi segnali dirigono il trasporto della proteina al compartimento di destinazione?
 PRINCIPI GENERALI DELLO SMISTAMENTO DELLE PROTEINE - Dove risiede l informazione per la corretta localizzazione di una proteina? - In che modo questi segnali dirigono il trasporto della proteina al compartimento
PRINCIPI GENERALI DELLO SMISTAMENTO DELLE PROTEINE - Dove risiede l informazione per la corretta localizzazione di una proteina? - In che modo questi segnali dirigono il trasporto della proteina al compartimento
06_citologia_SER_golgi 1
 1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
Il traffico vescicolare
 Il traffico vescicolare Gemmazione di vescicole rivestite di clatrina dalla membrana plasmatica e dal TGN: Come fa una vescicola di trasporto a scegliere il proprio carico specifico? ci riesce grazie alle
Il traffico vescicolare Gemmazione di vescicole rivestite di clatrina dalla membrana plasmatica e dal TGN: Come fa una vescicola di trasporto a scegliere il proprio carico specifico? ci riesce grazie alle
LE BASI DELL ORGANIZZAZIONE BIOLOGICA
 LE BASI DELL ORGANIZZAZIONE BIOLOGICA TABELLA: Le caratteristiche delle cellule procariotiche ed eucariotiche PROCARIOTI EUCARIOTI Organismi batteri e cianobatteri protisti, funghi, piante, animali Diametro
LE BASI DELL ORGANIZZAZIONE BIOLOGICA TABELLA: Le caratteristiche delle cellule procariotiche ed eucariotiche PROCARIOTI EUCARIOTI Organismi batteri e cianobatteri protisti, funghi, piante, animali Diametro
RE granuloso (RER) 1- Taglio proteolitico del peptide segnale. vedi lezione precedente. MFN0366-A1 (I. Perroteau) - Reticolo endoplasmatico
 1- Taglio proteolitico del peptide segnale. vedi lezione precedente. MFN0366-A1 (I. Perroteau) - Reticolo endoplasmatico") MFN0366-A1 (I. Perroteau) - Reticolo endoplasmatico RE granuloso (RER) Sintesi (sul lato citoplasmatico del RER) e maturazione delle proteine di membrana, delle proteine secretorie e di quelle destinate
MFN0366-A1 (I. Perroteau) - Reticolo endoplasmatico RE granuloso (RER) Sintesi (sul lato citoplasmatico del RER) e maturazione delle proteine di membrana, delle proteine secretorie e di quelle destinate
La cellula eucariotica contiene una varietà di organelli circondati da membrane.
 La cellula eucariotica contiene una varietà di organelli circondati da membrane. Reticolo endoplasmatico ruvido Reticolo endoplasmatico liscio Nucleo Assenti nella maggior parte delle cellule vegetali
La cellula eucariotica contiene una varietà di organelli circondati da membrane. Reticolo endoplasmatico ruvido Reticolo endoplasmatico liscio Nucleo Assenti nella maggior parte delle cellule vegetali
La cellula e il sistema membranoso
 Genesi Sistema Membranoso La cellula e il sistema membranoso * Sistema Membranoso * 1& Citoplasma: ialoplasma e organuli cellulari Fase fluida: matrice ialoplasmatica, ialoplasma o citosol (55%) Sistema
Genesi Sistema Membranoso La cellula e il sistema membranoso * Sistema Membranoso * 1& Citoplasma: ialoplasma e organuli cellulari Fase fluida: matrice ialoplasmatica, ialoplasma o citosol (55%) Sistema
MECCANISMI DI ESOCITOSI ED ENDOCITOSI. Meccanismi di Trasporto mediati da vescicole
 MECCANISMI DI ESOCITOSI ED ENDOCITOSI Meccanismi di Trasporto mediati da vescicole ENDOCITOSI ENDOCITOSI Forma di trasporto che consente l ingresso in cellula di varie sostanze mediante la formazione di
MECCANISMI DI ESOCITOSI ED ENDOCITOSI Meccanismi di Trasporto mediati da vescicole ENDOCITOSI ENDOCITOSI Forma di trasporto che consente l ingresso in cellula di varie sostanze mediante la formazione di
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita
 -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita") MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione.
 Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione. Questo sistema di membrane sintetizza le proteine che sono destinate alla secrezione
Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione. Questo sistema di membrane sintetizza le proteine che sono destinate alla secrezione
materiale didattico, vietate riproduzione e vendita 1
 1 L apparato del Golgi è un organello a più compartimenti, che contengono un ordinata serie di enzimi, che sequenzialmente modificano le glicoproteine e i lipidi quando essi transitano dalle cisterne cis
1 L apparato del Golgi è un organello a più compartimenti, che contengono un ordinata serie di enzimi, che sequenzialmente modificano le glicoproteine e i lipidi quando essi transitano dalle cisterne cis
Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione.
 Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione. Questo sistema di membrane sintetizza le proteine che sono destinate alla secrezione
Tutte le membrane del sistema sono connesse da un traffico vescicolare anterogrado e retrogrado denominato via di secrezione. Questo sistema di membrane sintetizza le proteine che sono destinate alla secrezione
Plasma membrane. Endoplasmic reticulum. Nucleus. Golgi apparatus. Mitochondrion Lysosome. Ribosome
 Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
11/12/2015. Apparato di Golgi. Camillo Golgi. Eventi post traduzionali. Córteno, 1843 Pavia, : Cattedra di Istologia, UniPV
 Camillo Golgi Apparato di Golgi Córteno, 1843 Pavia, 1926 1876: Cattedra di Istologia, UniPV 1893: Rettore, UniPV 1906: Premio Nobel per la Medicina Disegno eseguito da Camillo Golgi dell'apparato reticolare
Camillo Golgi Apparato di Golgi Córteno, 1843 Pavia, 1926 1876: Cattedra di Istologia, UniPV 1893: Rettore, UniPV 1906: Premio Nobel per la Medicina Disegno eseguito da Camillo Golgi dell'apparato reticolare
Apparato di Golgi. centro di maturazione e smistamento delle proteine della via secretoria. Complesso di Golgi RER. Faccia di uscita (TRANS)
") Apparato di Golgi centro di maturazione e smistamento delle proteine della via secretoria Faccia di uscita (TRANS) Complesso di Golgi Faccia di entrata (CIS) RER Il complesso di Golgi è stato scoperto
Apparato di Golgi centro di maturazione e smistamento delle proteine della via secretoria Faccia di uscita (TRANS) Complesso di Golgi Faccia di entrata (CIS) RER Il complesso di Golgi è stato scoperto
COME E ORGANIZZATA UNA CELLULA?
 COME E ORGANIZZATA UNA CELLULA? CITOPLASMA Citoplasma è tutta la regione compresa tra l Involucro Nucleare e la Membrana Plasmatica Comprende gli organelli cellulari CITOSOL Citosol è la sostanza semifluida
COME E ORGANIZZATA UNA CELLULA? CITOPLASMA Citoplasma è tutta la regione compresa tra l Involucro Nucleare e la Membrana Plasmatica Comprende gli organelli cellulari CITOSOL Citosol è la sostanza semifluida
Sistema membranoso del citoplasma. Sistema vacuolare del citoplasma
 Sistema membranoso del citoplasma O Sistema vacuolare del citoplasma Sistema membranoso del citoplasma La grande maggioranza degli organuli cellulari è formata da sistemi di membrane intracitoplasmatiche
Sistema membranoso del citoplasma O Sistema vacuolare del citoplasma Sistema membranoso del citoplasma La grande maggioranza degli organuli cellulari è formata da sistemi di membrane intracitoplasmatiche
I trasportatori citoplasmatici sono vescicole di trasporto
 I trasportatori citoplasmatici sono vescicole di trasporto Via biosintetica o via secretoria (secr. regolata, secr. costitutiva) Via endocitica ALCUNI APPROCCI ALLO STUDIO DELLE ENDOMEMBRANE: autoradiografia
I trasportatori citoplasmatici sono vescicole di trasporto Via biosintetica o via secretoria (secr. regolata, secr. costitutiva) Via endocitica ALCUNI APPROCCI ALLO STUDIO DELLE ENDOMEMBRANE: autoradiografia
3. Citologia i. Strutture cellulari comuni tra cellule animali e vegetali
 Strutture cellulari comuni tra cellule animali e vegetali: CITOPLASMA CITOSCHELETRO RIBOSOMI RETICOLO ENDOPLASMATICO APPARATO DEL GOLGI MITOCONDRI NUCLEO PEROSSISOMI CITOPLASMA materiale gelatinoso incolore
Strutture cellulari comuni tra cellule animali e vegetali: CITOPLASMA CITOSCHELETRO RIBOSOMI RETICOLO ENDOPLASMATICO APPARATO DEL GOLGI MITOCONDRI NUCLEO PEROSSISOMI CITOPLASMA materiale gelatinoso incolore
7a. COMPARTIMENTI INTRACELLULARI
 7a. COMPARTIMENTI INTRACELLULARI contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale molte delle reazioni chimiche che avvengono
7a. COMPARTIMENTI INTRACELLULARI contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale molte delle reazioni chimiche che avvengono
COME E ORGANIZZATA UNA CELLULA?
 COME E ORGANIZZATA UNA CELLULA? CITOPLASMA Citoplasma è tutta la regione compresa tra l Involucro Nucleare e la Membrana Plasmatica Comprende gli organelli cellulari CITOSOL Citosol è la sostanza semifluida
COME E ORGANIZZATA UNA CELLULA? CITOPLASMA Citoplasma è tutta la regione compresa tra l Involucro Nucleare e la Membrana Plasmatica Comprende gli organelli cellulari CITOSOL Citosol è la sostanza semifluida
Membrane Biologiche. Barriere per confinare sostanze o attività in ambienti specifici. Costituite da lipidi e proteine. Confini Cellulari Organelli
 Membrane Biologiche Membrane Biologiche Barriere per confinare sostanze o attività in ambienti specifici. Confini Cellulari Organelli Costituite da lipidi e proteine. Sistema di Endomembrane Delimitano
Membrane Biologiche Membrane Biologiche Barriere per confinare sostanze o attività in ambienti specifici. Confini Cellulari Organelli Costituite da lipidi e proteine. Sistema di Endomembrane Delimitano
La membrana nucleare come invaginazione della membrana citoplasmatica.
 Relazioni topologiche tra i compartimenti cellulari. La membrana nucleare come invaginazione della membrana citoplasmatica. Nucleo e citosol topologicamente continui. Mitocondrio come batterio simbionte.
Relazioni topologiche tra i compartimenti cellulari. La membrana nucleare come invaginazione della membrana citoplasmatica. Nucleo e citosol topologicamente continui. Mitocondrio come batterio simbionte.
- organuli di nm costituenti essenziali di tutti gli organismi - possono essere liberi nel citoplasma o adesi alla superficie esterna del RER
 Ribosoma - organuli di 15-20 nm costituenti essenziali di tutti gli organismi - possono essere liberi nel citoplasma o adesi alla superficie esterna del RER Funzione: - sono la sede della sintesi di proteine
Ribosoma - organuli di 15-20 nm costituenti essenziali di tutti gli organismi - possono essere liberi nel citoplasma o adesi alla superficie esterna del RER Funzione: - sono la sede della sintesi di proteine
MFN0366-A1 (I. Perroteau) - indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita.
 - indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita.") 10_bct_2011 1 E la proteina nascente a determinare se il ribosoma che catalizza la sua sintesi deve rimanere libero oppure essere associato alla membrana del RER. 1 2 Proteine che hanno destinazione finale
10_bct_2011 1 E la proteina nascente a determinare se il ribosoma che catalizza la sua sintesi deve rimanere libero oppure essere associato alla membrana del RER. 1 2 Proteine che hanno destinazione finale
Vescicole trasferite da una membrana all altra mantengono la loro asimmetria. Orientamento porzione glicidica
 Vescicole trasferite da una membrana all altra mantengono la loro asimmetria Orientamento porzione glicidica L USCITA DAL RER AVVIENE MEDIANTE TRAFFICO VESCICOLARE Sintesi e modificazione nel RER di proteine
Vescicole trasferite da una membrana all altra mantengono la loro asimmetria Orientamento porzione glicidica L USCITA DAL RER AVVIENE MEDIANTE TRAFFICO VESCICOLARE Sintesi e modificazione nel RER di proteine
Permeabilità Di Membrana Cellulare
 Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Permeabilità Di Membrana Cellulare
 Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
Permeabilità Di Membrana Cellulare Le membrane biologiche rappresentano una barriera per le molecole polari La membrana cellulare controlla il volume, la componente ionica e molecolare della cellula Una
INIZIO DELLA TRADUZIONE. Proteine citoplasmatiche (ed anche nucleari,mitocondriali ecc. Proteine integrali di membrana. Proteine di secrezione
 INIZIO DELLA TRADUZIONE Proteine citoplasmatiche (ed anche nucleari,mitocondriali ecc. Proteine integrali di membrana Proteine di secrezione RER MITOCONDRI / CLOROPLASTI proteine di secrezione PROTEINE
INIZIO DELLA TRADUZIONE Proteine citoplasmatiche (ed anche nucleari,mitocondriali ecc. Proteine integrali di membrana Proteine di secrezione RER MITOCONDRI / CLOROPLASTI proteine di secrezione PROTEINE
Fisiologia cellulare e Laboratorio di colture cellulari
 Corso di laurea in Biotecnologie UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di colture cellulari Prof.ssa Luisa Gioia possibili sistemi di trasporto della cellula TRASPORTO TRANS-MEMBRANA
Corso di laurea in Biotecnologie UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di colture cellulari Prof.ssa Luisa Gioia possibili sistemi di trasporto della cellula TRASPORTO TRANS-MEMBRANA
Plasma membrane. Endoplasmic reticulum. Nucleus. Golgi apparatus. Mitochondrion Lysosome. Ribosome
 Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
Endoplasmic reticulum Plasma membrane Nucleus Golgi apparatus Ribosome Mitochondrion Lysosome Funzioni 1- Compartimentazione 2- Localizzazione per attività biochimiche 3- Barriera selettiva 4- Trasporto
La cellula eucariotica e i suoi organuli (seconda parte)
") Università di Ferrara Corso di Laurea in Scienze Motorie Primo anno di corso Corso di Biologia Applicata Lezione di Biologia Cellulare La cellula eucariotica e i suoi organuli (seconda parte) Dott.ssa
Università di Ferrara Corso di Laurea in Scienze Motorie Primo anno di corso Corso di Biologia Applicata Lezione di Biologia Cellulare La cellula eucariotica e i suoi organuli (seconda parte) Dott.ssa
Il sistema endocrino
 Il sistema endocrino Indice delle lezioni: i LEZIONE 1 : Il sistema endocrino LEZIONE 2: Il sistema neuroendocrino: asse ipotalamo ipofisi Le ghiandole surrenali L asse ipotalamo-ipofisi-gonadi LEZIONE
Il sistema endocrino Indice delle lezioni: i LEZIONE 1 : Il sistema endocrino LEZIONE 2: Il sistema neuroendocrino: asse ipotalamo ipofisi Le ghiandole surrenali L asse ipotalamo-ipofisi-gonadi LEZIONE
Trasporto/ diffusione/osmosi
 Trasporto/ diffusione/osmosi 1 La membrana plasmatica è una struttura dinamica Si tratta di un doppio strato fosfolipidico con proteine integrate. La consistenza oleosa del doppio strato consente i movimenti
Trasporto/ diffusione/osmosi 1 La membrana plasmatica è una struttura dinamica Si tratta di un doppio strato fosfolipidico con proteine integrate. La consistenza oleosa del doppio strato consente i movimenti
STRUTTURA E FUNZIONI
 STRUTTURA E FUNZIONI Typical Cell Membrana plasmatica Chiamata anche membrana cellulare Circonda ogni cellula Separa il contenuto cellulare dal cio che la circonda Separa il LIC dal LEC Controlla il movimento
STRUTTURA E FUNZIONI Typical Cell Membrana plasmatica Chiamata anche membrana cellulare Circonda ogni cellula Separa il contenuto cellulare dal cio che la circonda Separa il LIC dal LEC Controlla il movimento
Dr.ssa Mauro ricevimento studenti
 Dr.ssa Mauro ricevimento studenti il lunedì dalle ore 12.00 alle ore 13.30 o previo appuntamento telefonico 0861.266888 E-mail: amauro@unite.it N.d.A.: Il materiale contenuto nel presente file è inteso
Dr.ssa Mauro ricevimento studenti il lunedì dalle ore 12.00 alle ore 13.30 o previo appuntamento telefonico 0861.266888 E-mail: amauro@unite.it N.d.A.: Il materiale contenuto nel presente file è inteso
Struttura e funzione delle membrane biologiche
 La membrana plasmatica delimita la cellula e separa l ambiente interno da quello esterno. Non impedisce però tutti gli scambi Struttura e funzione delle membrane biologiche Figure'11)1'!Essen&al!Cell!Biology!(
La membrana plasmatica delimita la cellula e separa l ambiente interno da quello esterno. Non impedisce però tutti gli scambi Struttura e funzione delle membrane biologiche Figure'11)1'!Essen&al!Cell!Biology!(
Lezione 4 - La cellula eucariotica ed i suoi organuli
 Lezione 4 - La cellula eucariotica ed i suoi organuli Membrana nucleare Citoscheletro Pori nucleari Citosol Centrioli Ribosomi Perossisomi Vantaggi della compartimentalizzazione: 1. Creazione di un microambiente
Lezione 4 - La cellula eucariotica ed i suoi organuli Membrana nucleare Citoscheletro Pori nucleari Citosol Centrioli Ribosomi Perossisomi Vantaggi della compartimentalizzazione: 1. Creazione di un microambiente
Reticolo Endoplasmatico Liscio, REL
 Reticolo Endoplasmatico Liscio Reticolo Endoplasmatico Liscio, REL E più tubulare del RER e forma un subcompartimento reticolare interconnesso del RE. E distribuito in un modo abbastanza omogeneo nel citoplasma.
Reticolo Endoplasmatico Liscio Reticolo Endoplasmatico Liscio, REL E più tubulare del RER e forma un subcompartimento reticolare interconnesso del RE. E distribuito in un modo abbastanza omogeneo nel citoplasma.
La cellula vegetale. Scienze e Tecnologie Agrarie (STAg) Tecnologie Forestali e Ambientali (TFA) Biotecnologie Agrarie (BA) Biologia Vegetale
 Tecnologie Forestali e Ambientali (TFA) Biotecnologie Agrarie (BA) Biologia Vegetale") Scienze e Tecnologie Agrarie (STAg) Tecnologie Forestali e Ambientali (TFA) Biotecnologie Agrarie (BA) Biologia Vegetale 1 La cellula vegetale 2 1 Il protoplasto 3 La cellula vegetale + o come negli animali
Scienze e Tecnologie Agrarie (STAg) Tecnologie Forestali e Ambientali (TFA) Biotecnologie Agrarie (BA) Biologia Vegetale 1 La cellula vegetale 2 1 Il protoplasto 3 La cellula vegetale + o come negli animali
Trasporto di sostanze in e out dalla cellula
 Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Fisiologia cellulare e Laboratorio di Colture cellulari
 Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
Corso di laurea BIOTECNOLOGIE UNIVERSITA DEGLI STUDI DI TERAMO Fisiologia cellulare e Laboratorio di Colture cellulari Prof.ssa Luisa Gioia Corso di laurea BIOTECNOLOGIE Fisiologia cellulare e Laboratorio
Università degli Studi del Sannio. Facoltà di Scienze MM.FF.NN. - Corso di Laurea in Biotecnologie a.a Programma di Biologia Cellulare
 Università degli Studi del Sannio Facoltà di Scienze MM.FF.NN. - Corso di Laurea in Biotecnologie a.a. 2010-2011 Programma di Biologia Cellulare (Prof Massimo Mallardo, I semestre, I anno) massimo.mallardo@unina.it
Università degli Studi del Sannio Facoltà di Scienze MM.FF.NN. - Corso di Laurea in Biotecnologie a.a. 2010-2011 Programma di Biologia Cellulare (Prof Massimo Mallardo, I semestre, I anno) massimo.mallardo@unina.it
CITOSOL
 www.fisiokinesiterapia.biz CITOSOL Il citosol (detto anche matrice citoplasmatica) è una sostanza che si presenta fisicamente gelatinosa contenente molecole di varie dimensioni e costituisce la porzione
www.fisiokinesiterapia.biz CITOSOL Il citosol (detto anche matrice citoplasmatica) è una sostanza che si presenta fisicamente gelatinosa contenente molecole di varie dimensioni e costituisce la porzione
Lezione 1: Atomi e molecole:
 Lezione 1: Atomi e molecole: La materia è costituita da elementi chimici in forma pura o in combinazioni dette composti. La vita richiede circa 25 elementi chimici. La struttura atomica determina il comportamento
Lezione 1: Atomi e molecole: La materia è costituita da elementi chimici in forma pura o in combinazioni dette composti. La vita richiede circa 25 elementi chimici. La struttura atomica determina il comportamento
Vescicole extracellulari
 Vescicole extracellulari Endocitosi e corpi multivescicolari Il sistema endosomi/lisosomi Huotari J, Helenius A. Endosome maturation. EMBO J. 2011 Aug 31;30(17):3481 500. 1 Didascalia figura Huotari (Sistema
Vescicole extracellulari Endocitosi e corpi multivescicolari Il sistema endosomi/lisosomi Huotari J, Helenius A. Endosome maturation. EMBO J. 2011 Aug 31;30(17):3481 500. 1 Didascalia figura Huotari (Sistema
Imprinting. Meccanismo epigenetico di controllo della espressione genica. repressione della trascrizione espressione mono-allelica
 Imprinting Meccanismo epigenetico di controllo della espressione genica. Metilazione repressione della trascrizione espressione mono-allelica Esistono determinati geni in cui l espressione di un allele
Imprinting Meccanismo epigenetico di controllo della espressione genica. Metilazione repressione della trascrizione espressione mono-allelica Esistono determinati geni in cui l espressione di un allele
Lezione 3. Le membrane cellulari: struttura e funzioni
 Lezione 3 Le membrane cellulari: struttura e funzioni FUNZIONI DELLA MEMBRANA PLASMATICA Barriera di permeabilità Barriera selettiva per determinate sostanze Divisione in compartimenti funzionali FUNZIONI
Lezione 3 Le membrane cellulari: struttura e funzioni FUNZIONI DELLA MEMBRANA PLASMATICA Barriera di permeabilità Barriera selettiva per determinate sostanze Divisione in compartimenti funzionali FUNZIONI
membrana cellulare membrana nucleare Nucleoloplasma nucleolo citoplasma reticolo endoplasmatico liscio
 Il secondo carattere distintivo delle cellule eucariote è costituito dalla presenza nel citoplasma di una serie eterogenea di organuli cellulari con strutture e funzioni specifiche relativamente costanti
Il secondo carattere distintivo delle cellule eucariote è costituito dalla presenza nel citoplasma di una serie eterogenea di organuli cellulari con strutture e funzioni specifiche relativamente costanti
 Hsp90 complessi con recettori per ormoni glucocorticoidi Hsp70 BiP in RE eucarioti, DnaK E. coli Hsp40 DnaJ in E. coli Hsp60 GroEL in E. coli Hsp10 GroES in E. coli H2O Pi C(ADP) Uprot C(ATP) Uprot-C(ADP)
Hsp90 complessi con recettori per ormoni glucocorticoidi Hsp70 BiP in RE eucarioti, DnaK E. coli Hsp40 DnaJ in E. coli Hsp60 GroEL in E. coli Hsp10 GroES in E. coli H2O Pi C(ADP) Uprot C(ATP) Uprot-C(ADP)
1. Fisiologia del trasporto delle lipoproteine
 I INTRODUZIONE 1. Fisiologia del trasporto delle lipoproteine Le lipoproteine sono aggregati lipido-proteici, eterogenei per dimensioni e composizione ma simili per struttura che assicurano un efficace
I INTRODUZIONE 1. Fisiologia del trasporto delle lipoproteine Le lipoproteine sono aggregati lipido-proteici, eterogenei per dimensioni e composizione ma simili per struttura che assicurano un efficace
Base cellulare della vita
 Base cellulare della vita La cellula è l unità strutturale e funzionale degli organismi viventi. Struttura minima in grado di compiere tutte le attività minime della vita. Teoria cellulare (Schleiden e
Base cellulare della vita La cellula è l unità strutturale e funzionale degli organismi viventi. Struttura minima in grado di compiere tutte le attività minime della vita. Teoria cellulare (Schleiden e
Caratteristiche generali dei sistemi viventi
 Caratteristiche generali dei sistemi viventi 1 Unicità chimica 2 Complessità ed organizzazione gerarchica 3 Metabolismo 4 Interazione ambientale: Regolazione e omeostasi 5 Riproduzione 6 Sviluppo 7 Evoluzione
Caratteristiche generali dei sistemi viventi 1 Unicità chimica 2 Complessità ed organizzazione gerarchica 3 Metabolismo 4 Interazione ambientale: Regolazione e omeostasi 5 Riproduzione 6 Sviluppo 7 Evoluzione
Sistema di Endomembrane
 RETICOLO ENDOPLASMATICO & INVOLUCRO NUCLEARE Sistema di Endomembrane Biotecnologie http://en.wikipedia.org/wiki/endomembrane_system http://www.microscopy.fsu.edu/cells/endoplasmicreticulum/endoplasmicreticulum.html
RETICOLO ENDOPLASMATICO & INVOLUCRO NUCLEARE Sistema di Endomembrane Biotecnologie http://en.wikipedia.org/wiki/endomembrane_system http://www.microscopy.fsu.edu/cells/endoplasmicreticulum/endoplasmicreticulum.html
A FRONTE DI UNA NOTEVOLE VARIETA DI FORME E FUNZIONI, DIFFERENTI TIPI DI CELLULE SONO ACCOMUNATI DAL POSSEDERE TUTTE: 1) CITOPLASMA; 2) NUCELOIDE O NU
 CITOPLASMA; 2) NUCELOIDE O NU") TEORIA CELLULARE (Virchow; Schleiden; Schwann. 1850) 1)Tutti gli organismi sono formati da una o più cellule; 2)La cellula è la più piccola unità che presenta le proprietà tipiche della vita; 3)Le cellule
TEORIA CELLULARE (Virchow; Schleiden; Schwann. 1850) 1)Tutti gli organismi sono formati da una o più cellule; 2)La cellula è la più piccola unità che presenta le proprietà tipiche della vita; 3)Le cellule
5. Trasporto di membrana
 5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
Cos è la Fisiologia Scienza che studia la vita e le funzioni organiche dei vegetali, degli animali e dell uomo
 Cos è la Fisiologia Scienza che studia la vita e le funzioni organiche dei vegetali, degli animali e dell uomo Cosa studia? Studia l omeostasi, ovvero tutti i processi con i quali è mantenuto costante
Cos è la Fisiologia Scienza che studia la vita e le funzioni organiche dei vegetali, degli animali e dell uomo Cosa studia? Studia l omeostasi, ovvero tutti i processi con i quali è mantenuto costante
COMUNICAZIONE INTERCELLULARE
 COMUNICAZIONE INTERCELLULARE TRASFERIMENTO DIRETTO DI SEGNALI CHIMICI E ELETTRICI ATTRAVERSO GIUNZIONI COMUNICANTI COMUNICAZIONE CHIMICA LOCALE (SOSTANZE PARACRINE E AUTOCRINE) COMUNICAZIONE A LUNGA DISTANZA
COMUNICAZIONE INTERCELLULARE TRASFERIMENTO DIRETTO DI SEGNALI CHIMICI E ELETTRICI ATTRAVERSO GIUNZIONI COMUNICANTI COMUNICAZIONE CHIMICA LOCALE (SOSTANZE PARACRINE E AUTOCRINE) COMUNICAZIONE A LUNGA DISTANZA
FUNZIONI delle MODIFICAZIONI delle proteine per GLICOSILAZIONE
 FUNZIONI delle MODIFICAZIONI delle proteine per GLICOSILAZIONE - Specificare la struttura tridimensionale: La glicosilazione permette alla proteina di assumere conformazioni tridimensionali complesse.
FUNZIONI delle MODIFICAZIONI delle proteine per GLICOSILAZIONE - Specificare la struttura tridimensionale: La glicosilazione permette alla proteina di assumere conformazioni tridimensionali complesse.
Reticolo endoplasmatico Apparato del Golgi Lisosomi
 Reticolo endoplasmatico Apparato del Golgi Lisosomi NELLA CELLULA eucariotica: Il Nucleo controlla le funzioni cellulari Il Citoplasma è sede di attività sintetiche e metaboliche della cellula Citoplasma
Reticolo endoplasmatico Apparato del Golgi Lisosomi NELLA CELLULA eucariotica: Il Nucleo controlla le funzioni cellulari Il Citoplasma è sede di attività sintetiche e metaboliche della cellula Citoplasma
 http://digilander.libero.it/glampis64 I composti organici di interesse biologico sono: Carboidrati Proteine Lipidi Acidi nucleici Le macromolecole sono composti di grandi dimensioni, unione di piccole
http://digilander.libero.it/glampis64 I composti organici di interesse biologico sono: Carboidrati Proteine Lipidi Acidi nucleici Le macromolecole sono composti di grandi dimensioni, unione di piccole
Il volume delle cellule è approssimativamente costante per ciascun tipo cellulare nell ambito della stessa specie o di specie affini ed è
 Introduzione allo studio della citologia Introduzione allo studio della citologia Legge di Driesch Il volume delle cellule è approssimativamente costante per ciascun tipo cellulare nell ambito della stessa
Introduzione allo studio della citologia Introduzione allo studio della citologia Legge di Driesch Il volume delle cellule è approssimativamente costante per ciascun tipo cellulare nell ambito della stessa
La cellula. Unità fondamentale di tutti gli organismi viventi.
 La cellula Unità fondamentale di tutti gli organismi viventi. Gli organismi sono tutti costituiti da cellule. La cellula è considerata l unità fondamentale di tutti i viventi. Ogni cellula nasce, cresce,
La cellula Unità fondamentale di tutti gli organismi viventi. Gli organismi sono tutti costituiti da cellule. La cellula è considerata l unità fondamentale di tutti i viventi. Ogni cellula nasce, cresce,
LISOSOMI. Vescicole (0.2 e 0.6 µm) rivestite da una membrana, contenenti enzimi idrolitici per la digestione intracellulare di macromolecole
 rivestite da una membrana, contenenti enzimi idrolitici per la digestione intracellulare di macromolecole") LISOSOMI Vescicole (0.2 e 0.6 µm) rivestite da una membrana, contenenti enzimi idrolitici per la digestione intracellulare di macromolecole Gli enzimi sono delle idrolasi acide, (circa 40 tipi diversi)
LISOSOMI Vescicole (0.2 e 0.6 µm) rivestite da una membrana, contenenti enzimi idrolitici per la digestione intracellulare di macromolecole Gli enzimi sono delle idrolasi acide, (circa 40 tipi diversi)
LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.
 LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.il modello a mosaico fluido descrive la struttura comune a tutte
LE MEMBRANE Le membrane sono composte da lipidi e proteine in composizioni che variano in base alla specie,al tipo cellulare e all organello.il modello a mosaico fluido descrive la struttura comune a tutte
Trasporto di sostanze in e out dalla cellula
 Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Trasporto Trasporto di sostanze in e out dalla cellula Tutte le sostanze che devono entrare o uscire dalla cellula devono passare la barriera della membrana plasmatica Le membrane biologiche sono selettivamente
Traffico di vescicole, endocitosi ed esocitosi. Biotecnologie Agro- Industriali. Biologia Cellulare. M.E. Miranda Banos
 IL TRASPORTO VESCICOLARE Le vescicole di trasporto collegano i diversi organuli membranosi e l esterno della cellula, entrando per fusione e uscendo per gemmazione con@nuamente. Trasportano proteine, solubili
IL TRASPORTO VESCICOLARE Le vescicole di trasporto collegano i diversi organuli membranosi e l esterno della cellula, entrando per fusione e uscendo per gemmazione con@nuamente. Trasportano proteine, solubili
RETICOLO ENDOPLASMATICO LISCIO, APPARATO DEL GOLGI, LISOSOMI E PEROSSISOMI
 Corso di Biologia Anno Accademico 2015/2016 Corso di Laurea in Scienze Motorie RETICOLO ENDOPLASMATICO LISCIO, APPARATO DEL GOLGI, LISOSOMI E PEROSSISOMI Dott. Andrea Puozzo Membrana nucleare Ribosomi
Corso di Biologia Anno Accademico 2015/2016 Corso di Laurea in Scienze Motorie RETICOLO ENDOPLASMATICO LISCIO, APPARATO DEL GOLGI, LISOSOMI E PEROSSISOMI Dott. Andrea Puozzo Membrana nucleare Ribosomi
Lezione 2. La cellula eucariote: organelli e citoscheletro
 Lezione 2 La cellula eucariote: organelli e citoscheletro IL RETICOLO ENDOPLASMATICO IL RETICOLO ENDOPLASMATICO Reticolo di membrane organizzate in strutture tubulari e cisterne appiattite che si estendono
Lezione 2 La cellula eucariote: organelli e citoscheletro IL RETICOLO ENDOPLASMATICO IL RETICOLO ENDOPLASMATICO Reticolo di membrane organizzate in strutture tubulari e cisterne appiattite che si estendono
Lezione 3. Dentro la cellula eucariote. Bibliografia. I colori della biologia. Giusti Gatti Anelli. Ed. Pearson
 Lezione 3 Dentro la cellula eucariote Bibliografia I colori della biologia Giusti Gatti Anelli Ed. Pearson Quali sono la struttura e le funzioni della membrana plasmatica? Qual è la funzione del nucleo?
Lezione 3 Dentro la cellula eucariote Bibliografia I colori della biologia Giusti Gatti Anelli Ed. Pearson Quali sono la struttura e le funzioni della membrana plasmatica? Qual è la funzione del nucleo?
La cellula. Teoria cellulare. Cellula. Organizzazione cellulare Come si studia la cellula
 La cellula Cellula Teoria cellulare Teoria cellulare Organizzazione e dimensioni della cellula Metodi di studio della cellula Cellule procariote ed eucariote Nucleo cellulare Organuli citoplasmatici Citoscheletro
La cellula Cellula Teoria cellulare Teoria cellulare Organizzazione e dimensioni della cellula Metodi di studio della cellula Cellule procariote ed eucariote Nucleo cellulare Organuli citoplasmatici Citoscheletro
Dischi piatti (cisterne) impilate con una polarità. Microvescicole Macrovescicole Tre regioni: Cis Mediale Trans
 impilate con una polarità. Microvescicole Macrovescicole Tre regioni: Cis Mediale Trans") Apparato di Golgi Aspetto dell apparato di Golgi Dischi piatti (cisterne) impilate con una polarità. Microvescicole Macrovescicole Tre regioni: Cis Mediale Trans Funzioni dell Apparato di Golgi Stazione
Apparato di Golgi Aspetto dell apparato di Golgi Dischi piatti (cisterne) impilate con una polarità. Microvescicole Macrovescicole Tre regioni: Cis Mediale Trans Funzioni dell Apparato di Golgi Stazione
Lisosomi (solo cellule animali) (x piante vd vacuolo; Biol. Cell Veg)
 (x piante vd vacuolo; Biol. Cell Veg)") Lisosomi (solo cellule animali) (x piante vd vacuolo; Biol. Cell Veg) Biotecnologie Christian de Duve http://www.nytimes.com/2013/05/07/us/christian de duve nobel winning biochemist dies at 95.html 1 http://micro.magnet.fsu.edu/cells/lysosomes/lysosomes.html
Lisosomi (solo cellule animali) (x piante vd vacuolo; Biol. Cell Veg) Biotecnologie Christian de Duve http://www.nytimes.com/2013/05/07/us/christian de duve nobel winning biochemist dies at 95.html 1 http://micro.magnet.fsu.edu/cells/lysosomes/lysosomes.html
16/12/2011. Citosol Ribosomi Sintesi proteica Introduzione al Sistema di Endomembrane CITOSOL (1) CITOSOL (2) CITOSOL (3) Biotecnologie_2011
 CITOSOL (2) CITOSOL (3) Biotecnologie_2011") Citosol Ribosomi Sintesi proteica Introduzione al Sistema di Endomembrane Biotecnologie_2011 CITOSOL (1) Tutta la porzione non strutturata che costituisce la parte liquida del citoplasma. In esso si trovano
Citosol Ribosomi Sintesi proteica Introduzione al Sistema di Endomembrane Biotecnologie_2011 CITOSOL (1) Tutta la porzione non strutturata che costituisce la parte liquida del citoplasma. In esso si trovano
LIPIDI COMPLESSI E LIPOPROTEINE
 LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre
, dall interazione con altre") Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre cellule (contatto cellula-cellula) o con strutture extracellulari
Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre cellule (contatto cellula-cellula) o con strutture extracellulari
IL SISTEMA ENDOMEMBRANOSO
 IL SISTEMA ENDOMEMBRANOSO L intenso"traffico" cellulare è costituito dalle vescicole che attraversano la cellula in direzioni opposte: dall'interno verso l'esterno o viceversa. Le proteine che si trovano
IL SISTEMA ENDOMEMBRANOSO L intenso"traffico" cellulare è costituito dalle vescicole che attraversano la cellula in direzioni opposte: dall'interno verso l'esterno o viceversa. Le proteine che si trovano
Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori
 Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori I trasportatori somigliano agli enzimi: catalizzano la traslocazione dei substrati da una parte all altra della membrana
Diffusione facilitata Trasporti mediati da proteine di membrana: i trasportatori I trasportatori somigliano agli enzimi: catalizzano la traslocazione dei substrati da una parte all altra della membrana
LEZIONE 1_TRASPORTI DI MEMBRANA
 LEZIONE 1_TRASPORTI DI MEMBRANA 1 2 I TRASPORTI DI MEMBRANA Diffusione semplice Diffusione facilitata Trasporto attivo 3 La diffusione semplice Lezione_1_trasporti 4 La diffusione semplice Relazione tra
LEZIONE 1_TRASPORTI DI MEMBRANA 1 2 I TRASPORTI DI MEMBRANA Diffusione semplice Diffusione facilitata Trasporto attivo 3 La diffusione semplice Lezione_1_trasporti 4 La diffusione semplice Relazione tra
Le MEMBRANE in biologia
 Le MEMBRANE in biologia Le membrana plasmatica Delimitazione delle cellule Genesi del potenziale elettrico trans-membrana Mantenimento delle differenze tra l ambiente intra ed extracellulare Trasferimento
Le MEMBRANE in biologia Le membrana plasmatica Delimitazione delle cellule Genesi del potenziale elettrico trans-membrana Mantenimento delle differenze tra l ambiente intra ed extracellulare Trasferimento
Le MEMBRANE in biologia
 Le MEMBRANE in biologia Le membrana plasmatica Delimitazione delle cellule Genesi del potenziale elettrico trans-membrana Mantenimento delle differenze tra l ambiente intra ed extracellulare Trasferimento
Le MEMBRANE in biologia Le membrana plasmatica Delimitazione delle cellule Genesi del potenziale elettrico trans-membrana Mantenimento delle differenze tra l ambiente intra ed extracellulare Trasferimento
Cell Signaling. Qualsiasi tipo di comunicazione tra le cellule
 Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La
Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La