Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
|
|
|
- Rosalia Di Lorenzo
- 8 anni fa
- Visualizzazioni
Transcript
1 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 31

2 Lipidi I lipidi si dividono in due gruppi principali: I gruppo: comprende composti a catena aperta con una testa polare e lunghe code apolari (acidi grassi, triacilgliceroli, fosfolipidi, sfingolipidi e glicolipidi) II gruppo: comprende composti ad anelli fusi, gli steroidi (tra cui il colesterolo). Esistono poi lipidi associati a proteine (lipoproteine) e polisaccaridi (lipopolisaccaridi).
.")
3 Acidi grassi Gli acidi grassi sono acidi carbossilici con una catena idrocarburica composta da 4 a 36 atomi di carbonio. Sono composti anfipatici perché contengono il gruppo carbossilico idrofilo e una catena idrocarburica idrofobica. Il gruppo carbossilico può ionizzare. La catena idocarburica può contenere solo legami singoli (acidi grassi saturi) o può contenere uno (acidi grassi insaturi) o più (acidi grassi polinsaturi) doppi legami. Gli acidi grassi insaturi la stereochimica del doppio legame è spesso cis con una torsione nella coda idrocarburica.
 o può contenere uno (acidi grassi insaturi) o più (acidi")
4 Trigliceridi I grassi sono costituiti per la maggior parte da miscele di triacilgliceroli (o trigliceridi) triesteri di acidi grassi del glicerolo, un composto che contiene tre gruppi ossidrilici.

5 Triacilgliceroli I trigliceridi si accumulano nel tessuto adiposo negli adipociti e costituiscono una riserva concentrata di energia metabolica. L ossidazione completa dei grassi produce circa 9 kcal/g rispetto alle 4 kcal/g dei carboidrati e proteine. Il contenuto di calorie per grammo nei grassi è più di due volte superiore a quello dei carboidrati e delle proteine.

6 Digestione, assorbimento e trasporto dei lipidi Concetti chiave: I triacilgliceroli sono demoliti dalle lipasi e i prodotti derivanti dalla loro scissione sono assorbiti dall intestino. Le lipoproteine trasportano i lipidi tra l intestino, il fegato e gli altri tessuti.

7 Assorbimento dei grassi

8 Acidi biliari e i loro coniugati Affinché i lipidi, insolubili in acqua, siano accessibili alle lipasi che si trovano in un mezzo acquoso, vengono inglobati in micelle composte da acidi biliari. Gli acidi biliari, molecole anfipatiche sintetizzate nel fegato a partire da colesterolo e secrete dalla cistifellea, facilitano la digestione dei lipidi nell intestino.

9 Lipasi pancreatiche La maggior parte dei lipidi della dieta è costituita dai triacilgliceroli che devono però essere degradati ad acidi grassi per essere assorbiti attraverso l epitelio intestinale. Le lipasi, enzimi intestinali secreti dal pancreas, degradano i triacilgliceroli ad acidi grassi e monoacilglicerolo.

10 Lipasi pancreatiche Il legame estereo di ciascuna molecola di triacilglicerolo è orientato verso la superficie delle micelle. Il legame diventa così più suscettibile alla digestione in ambiente acquoso in quanto le lipasi agiscono a livello dell interfaccia lipide-acqua.

11 Assorbimento dei grassi I prodotti finali della digestione vengono trasportati in forma micellare nell epitelio intestinale dove vengono trasportati attraverso la membrana. I trigliceridi vengono risintetizzati ed impacchettati insieme ad altri lipidi e all apolipoproteina B-48 per formare i chilomicroni che vengono poi riversati nel sistema linfatico.

12 Lipoproteine A causa di scarsa o nulla solubilità nel plasma (mezzo acquoso), i lipidi hanno bisogno di ancorarsi a proteine per poter circolare nel sangue. I lipidi vengono trasportati nel sangue sottoforma di aggregati micellari lipoproteici (Lipoproteine) capaci di formare sospensioni stabili. Le lipoproteine sono particelle complesse, ad alto peso molecolare, che trasportano lipidi apolari (soprattutto trigliceridi ed esteri del colesterolo) e proteine definite apolipoproteine.

13 Struttura molecolare di un chilomicrone I chilomicroni hanno struttura globulare nella quale apolipoproteine, fosfolipidi e colesterolo formano un involucro polare entro il quale sono racchiusi, segregati dall ambiente esterno acquoso, triacilgliceroli ed esteri del colesterolo.

14 Le lipoproteine del plasma umano
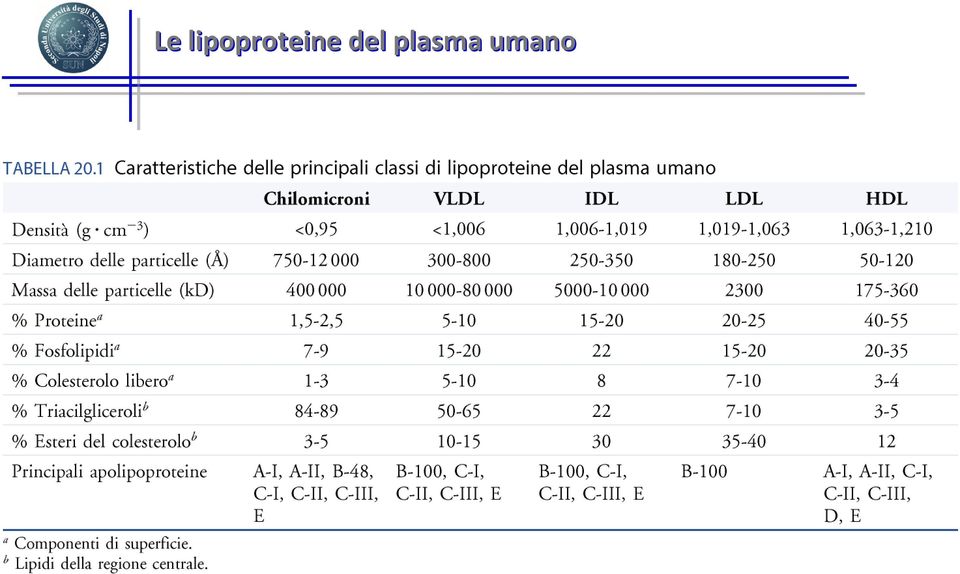
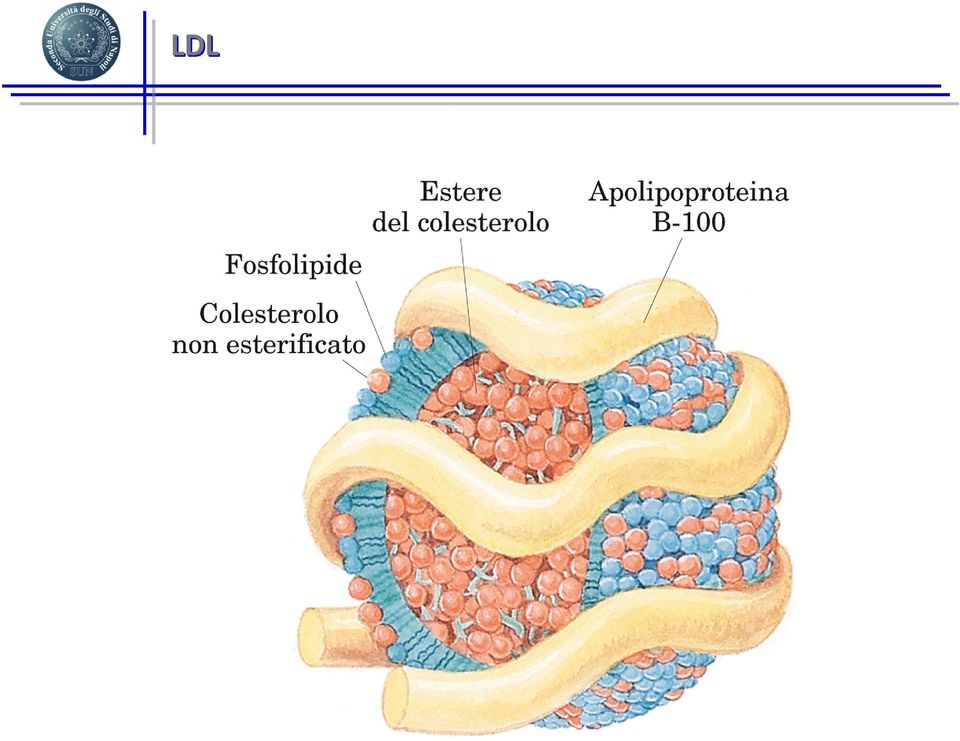
15 LDL
16 Il trasporto dei triacilgliceroli e del colesterolo

17 Colesterolo e aterosclerosi Le cellule periferiche non sono in grado di degradare il colesterolo in eccesso. Le HDL costituiscono un sistema che sottrae colesterolo libero ai tessuti periferici. Questo processo è ritenuto ANTI-ATEROGENO, ed un alto rapporto HDL/colesterolo conferisce all individuo un rischio ridotto di malattie coronariche. Le HDL prelevano il colesterolo dalle pareti delle arterie, ostacolando la formazione delle placche aterosclerotiche. Ecco perché il colesterolo HDL è comunemente detto buono. Le LDL, al contrario, depositano il colesterolo in eccesso sulle pareti delle arterie, favorendo così la formazione delle placche.

18 L ossidazione degli acidi grassi Concetti chiave: Gli acidi grassi da degradare sono legati al CoA tramite una reazione dipendente dall ATP. I gruppi acilici grassi sono trasportati nei mitocondri tramite il sistema navetta della carnitina per essere ossidati. A ogni giro della β-ossidazione mitocondriale sono prodotti FADH 2, NADH e acetil-coa. Per ossidare gli acidi grassi insaturi sono necessari ulteriori enzimi. Il propionil-coa prodotto dall ossidazione degli acidi grassi a catena dispari di atomi di carbonio è convertito a succinil-coa. I perossisomi ossidano gli acidi grassi a lunga catena producendo H 2 O 2.

19 L ossidazione degli acidi grassi L utilizzo degli acidi grassi come combustibili richiede tre stadi di trasformazione 1. I lipidi devono essere mobilizzati. In questo processo i triacilgliceroli vengono degradati ad acidi grassi e glicerolo, che vengono rilasciati dal tessuto adiposo e trasportati ai tessuti che richiedono energia. 2. Gli acidi grassi devono quindi essere attivati e trasportati nei mitocondri per la degradazione. 3. Nei mitocondri gli acidi grassi vengono degradati in modo graduale in acetil CoA, che poi viene ossidato nel ciclo dell acido citrico.

20 1. Mobilizzazione dei lipidi Affinché i grassi possano essere utilizzati come combustibili, i trigliceridi devono essere idrolizzati dalla lipasi per dare acidi grassi liberi. Lipasi Glicerolo Trigliceride Acidi grassi

21 1. Mobilizzazione dei lipidi L attività della lipasi è regolata dall AMP ciclico in risposta a stimoli ormonali. Glucagone e adrenalina stimolano i recettori 7TM che attivano l adenilato ciclasi che stimola la PKA a fosforilare due proteine, la perlipina e la lipasi, che portano alla mobilizzazione degli acidi grassi. ATGL: triacilglicerolo lipasi del tessuto adiposo Lipasi HS: lipasi ormone sensibile
22 1. Mobilizzazione dei lipidi Il glicerolo che deriva dall azione della lipasi viene captato nel fegato dove viene: Fosforilato a glicerolo 3-fosfato Ossidato a diidrossiacetone fosfato Isomerizzato a gliceraldeide 3-fosfato, intermedio della via glicolitica e gluconeogenetica
23 1. Mobilizzazione dei lipidi Il glicerolo può essere convertito in piruvato o a glucosio nel fegato. Gli acidi grassi vengono utilizzati come combustibili da molti tessuti mediante ossidazione.
24 2. Attivazione degli acidi grassi Prima di essere ossidati gli acidi grassi vengono attivati ossia legati al coenzima A. L adenosina trifosfato (ATP) favorisce la formazione di un legame tioestere tra il gruppo carbossilico di un acido grasso e il gruppo sulfidrilico del CoA. Acido grasso + CoA + ATP + H 2 O AcilCoA + AMP + 2 Pi + 2H La reazione avviene in due tappe, avviene sulla membrana mitocondriale esterna ed è catalizzata dall enzima acilcoa sintetasi. La reazione è irreversibile grazie all idrolisi del pirofosfato inorganico (equivalente all idrolisi di 2 ATP).
25 2. Attivazione degli acidi grassi Gli acidi grassi vengono attivati sulla membrana mitocondriale esterna mentre vengono ossidati nella matrice mitocondriale. La carnitina, un alcol zwitterionico, trasporta gli acidi grassi attivati nella matrice mitocondriale. Il gruppo acile viene trasferito allo zolfo del CoA formando l acil carnitina dalla carnitina pamitil trasferasi I (CPTI o Carnitina aciltransferasi I), enzima localizzato sulla membrana mitocondriale esterna.
26 2. Attivazione degli acidi grassi L acil carnitina attraversa la membrana mitocondriale interna servendosi di una traslocasi. Il gruppo acilico viene trasferito di nuovo al Coenzima A sul lato interno della membrana dalla carnitina pamitil trasferasi II (CPTII o Carnitina aciltransferasi II).
27 Trasporto dei gruppi acile attraverso la membrana mitocondriale
28 Fasi dell ossidazione degli acidi grassi
29 β-ossidazione degli acidi grassi Gli acidi grassi vengono degradati mediante la ripetizione di una sequenza di quattro reazioni: 1. Formazione di un doppio legame mediante ossidazione FAD dipendente 2. Idratazione del doppio legame per aggiungere un atomo di ossigeno 3. Ossidazione dell alcol a gruppo chetonico NAD + dipendente 4. L acido grasso viene scisso mediante tiolisi dal CoA per formare AcetilCoA e una catena di acido grasso di 2 atomi più corta
30 β-ossidazione degli acidi grassi La prima reazione di ciascun ciclo è l ossidazione dell acil CoA catalizzata dalla acil CoA deidrogenasi per formare un doppio legame tra il C2 ed il C3 utilizzando il FAD come accettore di elettroni. Gli elettroni provenienti dal gruppo prostetico FADH 2 dell acil CoA deidrogenasi vengono trasferiti a una seconda flavoproteina detta flavoproteina di trasferimento degli elettroni (ETF) che li trasferisce alla proteina ferro-zolfo ETF:ubichinone riduttasi che li convoglia attraverso la riduzione dell ubichinone ad ubichinolo alla catena di trasporto degli elettroni.
31 β-ossidazione degli acidi grassi La seconda reazione consiste nell idratazione del doppio legame tra il C2 ed il C3 catalizzata dalla enoil CoA idratasi. L idratazione è stereospecifica.
32 β-ossidazione degli acidi grassi La terza reazione consiste nell ossidazione che converte il gruppo ossidrilico in C3 in un gruppo chetonico. La reazione è catalizzata dalla L-3-idrossiacil CoA deidrogenasi NAD + dipendente.
33 β-ossidazione degli acidi grassi La quarta ed ultima reazione, catalizzata dalla β-chetotiolasi, consiste nella scissione del 3- chetoacil CoA ad opera del gruppo tiolico di una seconda molecola di CoA che produce una molecola di acetil CoA e una di acil CoA accorciata di due atomi di carbonio.
34 β-ossidazione degli acidi grassi
35 β-ossidazione degli acidi grassi L acil CoA accorciato subisce poi un altro ciclo di ossidazione a partire dalla reazione catalizzata dalla acil CoA deidrogenasi per cui le unità bicarboniose vengono rimosse sequenzialmente a partire dalla terminazione carbossilica dell acido grasso.
36 Il sito attivo dell acil-coa deidrogenasi specifica per una catena di media lunghezza Le acil CoA deidrogenasi possono essere specifiche per gli acidi grassi a catena lunga (C12-C18), media (C14-C4) e corta (C4-C6).
37 Primi tre cicli della degradazione del palmitato L ossidazione completa di una molecola di palmitato richiede 7 cicli di reazione durante i quali si formano 7 NADH, 7 FADH 2 e 8 Acetil CoA che vengono ossidati nel ciclo di Krebs.
38 Resa energetica per l ossidazione completa del palmitato Durante la fosforilazione ossidativa si producono 1.5 ATP per ogni FADH 2 e 2.5 per ogni NADH. Reazione Acil CoA deidrogenasi β-idrossiacil CoA deidrogenasi Isocitrato deidrogenasi α-chetoglutarato deidrogenasi Succinil CoA sintetasi Succinato deidrogenasi Malato deidrogenasi Totale Numero di NADH o FADH 2 Numero di ATP 7 FADH NADH 8 NADH 8 NADH 8 GTP 8 FADH 2 8 NADH L equivalente di 2 molecole di ATP viene consumato nell attivazione del palmitato (ATP scisso a AMP e 2 molecole di Pi), pertanto: L ossidazione completa di una molecola di palmitato produce 106 molecole di ATP
39 Gli orsi ossidano i loro grassi durante il letargo
40 β-ossidazione degli acidi grassi a numero dispari di atomi di C Con un acido grasso con numero pari di atomi di carbonio, durante l ultima tappa della β- ossidazione un acetoacetil CoA è scisso in 2 acetil CoA dalla tiolasi. La maggior parte dei lipidi presenti in natura contiene acidi grassi con un numero pari di atomi di carbonio. Gli acidi grassi a catena dispari sono presenti in quantità significative nei lipidi delle piante e in alcuni organismi marini. Il propionil CoA e l acetil CoA sono i prodotti finali della β-ossidazione quando viene ossidato un acido grasso a numero dispari di atomi di carbonio.
41 β-ossidazione degli acidi grassi a numero dispari di atomi di C Esiste una via enzimatica che converte il propionil CoA in succinil CoA che entra nel ciclo di Krebs. Il propionil CoA viene carbossilato dalla propionil CoA carbossilasi e subisce poi un riarrangiamento per formare succinil CoA. Poiché il propionil CoA è anche un prodotto del catabolismo degli amminoacidi tale processo ha un ruolo nel metabolismo degli amminoacidi.
42 β-ossidazione degli acidi grassi a numero dispari di atomi di C L ossidazione degli acidi grassi a catena dispari ha bisogno di altri tre enzimi: Una propionil CoA carbossilasi biotina-dipendente Una metilmalonil CoA racemasi Una metilmalonil CoA mutasi che richiede vitamina B12 (cobalammina) La metilmalonil CoA mutasi forma il succinil CoA mediante un riordinamento intramolecolare in cui il gruppo R può essere un gruppo amminico, ossidrilico o un carbonio sostituito.
43 Riarrangiamento catalizzato dalla metilmalonil-coa mutasi
44 Struttura del coenzima B12 Quella del coenzima B12 appartiene ad una classe di molecole variabili a seconda dei sostituenti (X). La 5 -deossiadenosilcobalammina è la forma presente nella metilmalonil CoA mutasi.
45 Formazione di un radicale 5 -deossiadenosilico La reazione della metilmalonil CoA mutasi comincia con la scissione omolitica del legame che unisce lo ione Co 3+ a un atomo di C del ribosio della porzione adenosinica dell enzima. La scissione genera un radicale 5 deossiadenosilico e determina la riduzione del Co 3+ a Co 2+. Il ruolo del coenzima B12 nelle migrazioni intramolecolari è quello di fungere da sorgente di radicali liberi per l estrazione di atomi di idrogeno. Una proprietà essenziale del coenzima è la debolezza del legame cobalto-carbonio, la cui facile scissione genera un radicale.
46 Formazione di succinil CoA mediante riarrangiamento Il radicale libero generato dal coenzima B12 estrae un atomo di idrogeno nel riarrangiamento del metilmalonil CoA a succinil CoA.
47 β-ossidazione degli acidi grassi monoinsaturi L ossidazione degli acidi grassi monoinsaturi richiede un altro enzima, l enoil CoA isomerasi, che modifica la posizione del doppio legame convertendo l isomero cis nell isomero trans, l intermedio normale della β ossidazione. Il doppio legame cis tra il C3 ed il C4 viene convertito il un doppio legame trans tra il C2 e C3, substrato ordinario della enoil CoA idratasi che prosegue nella β-ossidazione. L ossidazione degli acidi grassi poliinsaturi richiede oltre all enoil-coa isomerasi, un secondo enzima ausiliario la 2,4-dienol-CoA reduttasi NADPH dipendente.
48 β-ossidazione degli acidi grassi poliinsaturi Questi due enzimi convertono il 2,4-dienoil CoA in trans- 3-enoil CoA, che prosegue nella β ossidazione.
49 β-ossidazione degli acidi grassi poliinsaturi Queste strategie catalitiche sono eleganti ed economiche. Per l ossidazione di qualsiasi acido grasso polinsaturo sono sufficienti solo enzimi addizionali. I doppi legami in posizione dispari sono convertiti dall isomerasi. I doppi legami in posizione pari dalla riduttasi e dall isomerasi.
50 β-ossidazione perossisomiale Gli acidi grassi a lunga catena carboniosa sono ossidati anche nei perossisomi, piccoli organelli cellulari dotati di membrana presenti nelle cellule eucariotiche. L ossidazione perossisomiale differisce da quella mitocondriale a livello della reazione di deidrogenazione.
51 β-ossidazione perossisomiale Nei perossisomi l acil CoA deidrogenasi trasferisce gli elettroni dal FADH 2 all O 2 formando H 2 O 2 mentre nella βossidazione mitocondriale gli elettroni catturati dal FADH2 sono utilizzati nella catena di trasporto degli elettroni.
52 β-ossidazione perossisomiale
53 I corpi chetonici Concetti chiave: Nel fegato l acetil-coa può essere convertito reversibilmente a corpi chetonici che saranno utilizzati come fonte di energia dagli altri tessuti.
54 I corpi chetonici L acetil CoA che si forma nella β-ossidazione degli acidi grassi entra nel ciclo di Krebs solo se la degradazione dei grassi e quella dei carboidrati sono bilanciate. Il destino dell acetil-coa dipende dalla disponibilità di ossalacetato per formare citrato (prima reazione del ciclo dell acido citrico). ossalacetato Acetil CoA Ossalacetato (digiuno, diabete: l ossalacetato viene utilizzato per sintetizzare glucosio) Ciclo dell acido citrico Formazione di corpi chetonici
55 I corpi chetonici I corpi chetonici si formano principalmente nel fegato.
56 Formazione dei corpi chetonici dall acetil-coa L acetoacetato si forma a partire dal CoA in tre tappe: 1. Condensazione di due molecole di acetil CoA 2. Reazione con una seconda molecola di acetil CoA e H 2 O 3. Scissione in acetoacetato e acetil CoA
57 β-idrossibutirrato Il D-β-idrossibutirrato si forma per riduzione dell acetoacetato in una reazione NADH dipendente catalizzata dalla β-idrossibutirrato deidrogenasi.
58 Acetone L acetoacetato, essendo un β-chetoacido, subisce una decarbossilazione spontanea lenta ad acetone (può avvenire anche ad opera di una acetoacetato decarbossilasi).
59 Corpi chetonici come combustibile Durante il digiuno o in condizioni di diabete non trattato il fegato converte gli acidi grassi in corpi chetonici che fungono da combustibile per gli altri organi. L acetoacetato può essere considerato una forma solubile e trasportabile di unità acetile. L acetoacetato e il D-βidrossibutirrato sono trasportati dal sangue ai tessuti extraepatici dove sono ossidati nel ciclo dell acido citrico.
60 β-idrossibutirrato e acetoacetato come fonte di energia L acetoacetato e il 3-idrossibutirrato sono combustibili normali della respirazione e sono quantitativamente importanti come fonte energetica. Il muscolo cardiaco e la corteccia renale impiegano di preferenza l acetoacetato anziché il glucosio. Il glucosio, invece, è il principale combustibile del cervello e degli eritrociti negli individui ben nutriti che assumono una dieta bilanciata. Il cervello si adatta all utilizzo di acetoacetato durante il digiuno e nello stato diabetico.
61 β-idrossibutirrato e acetoacetato come fonte di energia L acetoacetato viene convertito in acetil CoA in due tappe: L acetoacetato viene attivato tramite una reazione di trasferimento del CoA dal succinil CoA Nella seconda l acetoacetil CoA viene scisso da una tiolasi per dare due molecole di acetil CoA
62 β-idrossibutirrato e acetoacetato come fonte di energia Il D-β-idrossibutirrato richiede un passaggio addizionale ovvero una ossidazione NAD + dipentente ad acetoacetato. Il NADH prodotto entra poi nella fosforilazione ossidativa.

63 Chetosi diabetica In assenza di insulina i grassi vengono rilasciati dal tessuto adiposo e il glucosio non può essere captato dal fegato o dal tessuto adiposo. Il fegato degrada i grassi attraverso la β-ossidazione, ma non può metabolizzare l acetil CoA prodotto perché i livelli di ossalacetato, metabolita che deriva dal glucosio, sono bassi. Si produce così un eccesso di corpi chetonici che vengono rilasciati nel sangue.
64 La biosintesi degli acidi grassi Concetti chiave: Il sistema di trasporto degli acidi tricarbossilici trasferisce acetil-coa nel citosol per la sintesi degli acidi grassi. La sintesi degli acidi grassi inizia con la carbossilazione dell acetil-coa che genera malonil- CoA. L acido grasso sintasi catalizza sette reazioni e allunga un acido grasso di due atomi di carbonio alla volta.
65 La biosintesi degli acidi grassi
66 La biosintesi degli acidi grassi Gli acidi grassi sono sintetizzati e degradati attraverso vie distinte: La sintesi degli acidi grassi avviene nel citosol, la degradazione nella matrice mitocondriale Gli intermedi della sintesi sono legati covalentemente ai gruppi sulfidrilici di una proteina trasportatrice di acili (ACP), mentre gli intermedi della degradazione sono legati al coenzima A La molecola attivata che cede unità bicarboniose nel processo di allungamento è il malonil- ACP al posto dell acetil CoA. La reazione di allungamento nella sintesi è favorita dal rilascio di CO 2. L agente riducente nella sintesi degli acidi grassi è il NADPH mentre nella via degradativa gli ossidanti sono il NAD + e il FAD Negli organismi superiori gli enzimi della sintesi degli acidi grassi sono uniti in una unica catena polipeptidica detta acido grasso sintasi L allungamento della catena ad opera della acido grasso sintasi si ferma alla formazione di palmitato (C16). Ulteriori allungamenti e l inserzione di doppi legami sono effettuati da altri sistemi enzimatici
67 Acetil-CoA carbossilasi La formazione di malonil CoA è la tappa di comando nella sintesi degli acidi grassi. La biosintesi degli acidi grassi inizia nel citosol con la formazione di malonil CoA catalizzata dalla acetil CoA carbossilasi, che contiene biotina. Come nel caso degli altri enzimi che contengono biotina (es. piruvato carbossilasi) si forma un intermedio carbossibiotina a spese di ATP. Il gruppo CO 2 attivato viene poi trasferito per formare malonil CoA.
68 Proteina trasportatrice di acili Gli intermedi della sintesi degli acidi grassi sono legati ad una proteina trasportatrice di acili (ACP). Il legame avviene al terminale sulfidrilico di una fosfopanteteina, lo stesso gruppo presente nel coenzima A.
69 Proteina trasportatrice di acili Sia la proteina trasportatrice di acili (ACP) che il coenzima A contengono fosfopanteteina.
70 La biosintesi degli acidi grassi La biosintesi degli acidi grassi comincia con la formazione di acetil ACP e malonil ACP ad opera degli enzimi acetil e malonil transacilasi, rispettivamente. 1. L acetil ACP e il malonil ACP reagiscono per formare acetoacetil ACP. Nella reazione viene rilasciata CO 2 Gruppo malonile Acetoacetil ACP Gruppo acetile Condensazione Acido grasso sintasi
71 La biosintesi degli acidi grassi 2. Riduzione del dell acetoacetil ACP a D-3-idrossibutirril ACP ad opera della β-chetoacil ACP riduttasi. L agente riducente è il NADPH. Acetoacetil ACP D-3-idrossibutirril ACP Riduzione
72 La biosintesi degli acidi grassi 3. Il D-3-idrossibutirril ACP viene deidratato formando il crotonil ACP ad opera della 3-idrossiacil ACP deidratasi. D-3-idrossibutirril ACP Crotonil ACP Deidratazione
73 La biosintesi degli acidi grassi 4. Il crotonil ACP viene ridotto a butirril ACP ad opera della enoil ACP riduttasi. L agente riducente è di nuovo il NADPH. Riduzione
74 Biosintesi del palmitato I cicli di allungamento si susseguono fino alla formazione di C16-acil-ACP, che non è più substrato dell enzima condensante, ma viene idrolizzato a palmitato e ACP. Quattro cicli
75 La biosintesi degli acidi grassi Il complesso dell acido grasso sintasi è mostrato in modo schematico sottoforma di un disco. Ogni segmento del disco rappresenta una delle sei attività enzimatiche del complesso. Al centro del disco si trova la proteina che trasporta gli acili (ACP), con il braccio fosfopanteteinico. Enzimi AT: acetil-coa-acp transacetilasi KS: β-chetoacil-acp sitasi MT: malonil-coa-acp transferasi KR: β-chetoacil-acp reduttasi HD: β-idrossiacil-acp deidratasi ER: enoil-acp reduttasi
76 La biosintesi degli acidi grassi Il gruppo acetilico dell acetil CoA viene trasferito al gruppo -SH di un residuo di cisteina della β-chetoacil sintasi (KS) Enzima: acetil-coa-acp transacetilasi Il gruppo malonile viene trasferito dal malonil CoA al gruppo -SH dell ACP Enzima: malonil-coa-acp transferasi
77 La biosintesi degli acidi grassi 1. Condensazione Enzima: β-chetoacil-acp sitasi (KS) 2. Riduzione Enzima: β-chetoacil-acp reduttasi (KR)
78 La biosintesi degli acidi grassi 3. Deidratazione Enzima: β-idrossiacil-acp deidratasi (HD) 4. Riduzione del doppio legame Enzima: enoil-acp reduttasi (ER)
79 La biosintesi degli acidi grassi La tappa successiva prevede la traslocazione del gruppo butirrilico sulla Cys della β-chetoacil-acp sintasi (KS). L enzima è nuovamente l acetil-coa-acp transacetilasi (AT) che può trasferire gruppi acile diversi dal gruppo acetile.
80 II ciclo delle reazioni del complesso dell acido grasso sintasi
81 Struttura delle varie acido grasso sintasi L acido grasso sintasi dei batteri e delle piante è un complesso formato da 7 polipeptidi diversi Nel lievito le sette attività sono riunite in 2 polipeptidi Nei vertebrati tutte le attività sono su di una sola catena polipeptidica
82 L acido grasso sintasi animale La proteina funzionale è un dimero. Ciascuna catena identica nel dimero contiene tre domini: Il dominio 1 contiene la acetil transferasi (AT), la malonil transferasi (MT) e l enzima condensante (CE). Il dominio 2 contiene la proteina trasportatrice di acili (ACP), la β- chetoacil riduttasi (KR), la deidratasi (DH) e la enoil reduttasi (ER). Il dominio 3 contiene la tioesterasi (TE) Il gruppo flessibile di fosfopanteteina trasporta la catena di acido grasso da un sito catalitico su una catena all altro, nonché tra le catene nel dimero.
83 Struttura dell acido grasso sintasi
84 Stechiometria della sintesi del palmitato Stechiometria della sintesi del palmitato: Acetil CoA + 7 malonil CoA + 14 NADPH + 20 H + palmitato + 7 CO NADP CoA + 6 H 2 O Equazione per la sintesi del malonil CoA: 7 Acetil CoA + 7 CO ATP 7 malonil CoA + 7 ADP + 7 Pi + 14 H + stechiometria complessiva 8 Acetil CoA + 7 ATP + 14 NADPH + 6 H + palmitato + 14 NADP CoA + 6 H 2 O + 7 ADP + 7 P i
85 Confronto tra b-ossidazione e biosintesi degli acidi grassi
86 Trasporto di acetili per la sintesi di acidi grassi Il citrato trasporta dai mitocondri al citosol i gruppi acetile che vengono usati per la sintesi degli acidi grassi. Per ogni acetil CoA trasferito dai mitocondri al citosol viene prodotta una molecola di NADPH per cui il potenziale di riduzione del NADH viene convertito in quello del NADPH.
87 Fonti di NADPH per la sintesi di acidi grassi L ossalacetato utilizzato per il trasferimento dei gruppi acetile viene restituito ai mitocondri. Poiché la membrana mitocondriale interna è impermeabile all ossalacetato, mediante una serie di reazioni, si forma piruvato che può rientrare nel mitocondrio. Durante tali reazioni viene generata una molecola di NADPH per ogni acetil CoA che viene trasferito dai mitocondri al citosol. Il NADPH è generato dall enzima malico e nella via del pentoso fosfato. 8 NADPH per gli 8 acetil CoA trasferiti per la sintesi del palmitato

88 Cooperazione di vie metaboliche nella sintesi di acidi grassi La sintesi degli acidi grassi comporta la cooperazione di diverse vie metaboliche localizzate in diversi compartimenti cellulari. Il ciclo di Krebs, la compartimentazione e la via del pentosio fosfato forniscono gli atomi di carbonio e il potere riducente. La glicolisi e la fosforilazione ossidativa l ATP per soddisfare le esigenze per la sintesi degli acidi grassi.
89 Schema riassuntivo del metabolismo lipidico
90 Allungamento ed insaturazione degli acidi grassi Il principale prodotto della sintesi degli acidi grassi è il palmitato. Negli eucarioti gli acidi grassi più lunghi vengono formati da reazioni di allungamento catalizzate da enzimi sulla faccia citosolica del reticolo endoplasmatico. Queste reazioni addizionano sequenzialmente unità bicarboniose del malonil CoA alle estremità carbossiliche dei substrati acil CoA sia saturi che insaturi.
91 Allungamento ed insaturazione degli acidi grassi I sistemi enzimatici del reticolo endoplasmatico hanno anche la funzione di introdurre doppi legami negli acil CoA a catena lunga ad opera di tre enzimi legati alla membrana che operano il trasferimento di elettroni che operano la desaturazione degli acidi grassi.
92 Ormoni eicosanoidi L arachidonato, un acido grasso 20:4 derivato dal linoleato, è il principale precursore degli ormoni eicosanoidi (dal greco eikosi, venti), molecole segnale coinvolte in svariate funzioni tra cui stimolano l infiammazione, regolano l irrorazione sanguigna. Acido acetilsalicilico
93 La regolazione del metabolismo degli acidi grassi Concetti chiave: L ossidazione e la sintesi degli acidi grassi sono regolate dagli ormoni e da fattori cellulari.
94 Regolazione del metabolismo degli acidi grassi La sintesi e la degradazione degli acidi grassi sono reciprocamente regolate in modo che non possano essere attivate simultaneamente. L acetil CoA carbossilasi svolge un ruolo chiave nella regolazione del metabolismo degli acidi grassi che viene inibita per fosforilazione dalla chinasi AMP-dipendente (AMPK) ed attivata per defosforilazione. L AMPK è un sensore dell energia attivato dall AMP e inibita dall ATP. Se il livelli di energia sono bassi la cabossilasi si inattiva ed i grassi non vengono sintetizzati quando vi è richiesta energetica.
95 Regolazione del metabolismo degli acidi grassi La carbossilasi è anche attivata allostericamente dal citrato che promuove la polimerizzazione dei dimeri inattivi formando dei filamenti attivi. La polimerizzazione rimuove inoltre parzialmente l inibizione per fosforilazione.
96 Regolazione del metabolismo degli acidi grassi Se i livelli di citrato sono alti, acetil CoA e ATP sono abbondanti ovvero vi sono precursori ed energia sufficienti per la sintesi degli acidi grassi. Al contrario, il palmitato, che abbonda quando vi è un eccesso di sintesi, provoca la disgregazione dei filamenti di carbossilasi. Il malonil CoA blocca inoltre l ingresso degli acil CoA nei mitocondri.
97 Regolazione del metabolismo degli acidi grassi L attività carbossilasica è inoltre controllata a livello ormonale essendo stimolata dall insulina ed inibita dal glucagone e dall adrenalina, ormoni che riflettono lo stato energetico dell organismo.
Attenzione : lunedì 29 aprile NON ci sarà lezione
 Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI
 LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
GLUCONEOGENESI. Sintesi (GENESI) di nuove (NEO) molecole di glucosio
 di nuove (NEO) molecole di glucosio") GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
METABOLISMO LIPIDICO. Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva. Lipidi introdotti con gli alimenti.
 METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
dotto di Wirsung che si riversa nella papilla maggiore del duodeno;
 Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Il ciclo dell acido citrico
 Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico 2014-15
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Βeta ossidazione degli acidi grassi BETA OSSIDAZIONE DEGLI ACIDI GRASSI 13/12/2013 SEMINARIO. Acidi grassi (2)
") Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Si possono suddividere in:
 Si possono suddividere in: LIPIDI DI DEPOSITO: si accumulano nelle cellule del tessuto adiposo, sono una riserva energetica, mantengono costante la temperatura del corpo (termogenesi) LIPIDI STRUTTURALI:
Si possono suddividere in: LIPIDI DI DEPOSITO: si accumulano nelle cellule del tessuto adiposo, sono una riserva energetica, mantengono costante la temperatura del corpo (termogenesi) LIPIDI STRUTTURALI:
digestione dei lipidi alimentari duodeno digiuno sali biliari fosfolipidi bile specifiche idrolasi pancreatiche colecistochinina
 La digestione dei lipidi alimentari, in prevalenza trigliceridi, avviene nel duodeno e nel digiuno per azione combinata dei sali biliari e dei fosfolipidi della bile e delle specifiche idrolasi pancreatiche
La digestione dei lipidi alimentari, in prevalenza trigliceridi, avviene nel duodeno e nel digiuno per azione combinata dei sali biliari e dei fosfolipidi della bile e delle specifiche idrolasi pancreatiche
Biosintesi dei triacilgliceroli
 Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
Da dove prendono energia le cellule animali?
 Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine
 Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine I grassi
GLICOGENOLISI & GLICOGENOSINTESI
 GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
METABOLISMO DEI LIPIDI
 METABOLISMO DEI LIPIDI Generalità I lipidi alimentari, per poter essere assorbiti dall organismo umano, devono essere preventivamente digeriti (ad eccezione del colesterolo), cioè, trasformati in molecole
METABOLISMO DEI LIPIDI Generalità I lipidi alimentari, per poter essere assorbiti dall organismo umano, devono essere preventivamente digeriti (ad eccezione del colesterolo), cioè, trasformati in molecole
Metabolismo degli aminoacidi
 Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
ANABOLISMO DEI LIPIDI
 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE. L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO
 GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
Idratazione (introduciamo un gruppo OH sul Cβ) per mezzo della enoil-coa idratasi
 per mezzo della enoil-coa idratasi") β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
«Macromolecole» Lipidi
 LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
SISTEMI ENERGETICI. L ATP privato di uno dei suoi 3 radicali fosforici diventa ADP (adenosindifosfato).
.") SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
METABOLISMO DEL Glucosio
 METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
Metabolismo degli amminoacidi. Ciclo dell urea
 Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
Regolazione del metabolismo del glucosio
 Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
FUNZIONI DEI MITOCONDRI
 FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
Metabolismo degli amminoacidi
 Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
CHIMICA BIOLOGICA. Università degli Studi della Campania Luigi Vanvitelli. DiSTABiF. Corso di Laurea in Scienze Biologiche.
 Università degli Studi della Campania Luigi Vanvitelli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2017-18 Lezione 16 Degradazione
Università degli Studi della Campania Luigi Vanvitelli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2017-18 Lezione 16 Degradazione
Niccolò Taddei Biochimica
 Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici
 I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
I L I P I D I. Lipidi complessi: fosfolipidi e glicolipidi; sono formati da CHO e altre sostanze.
 I L I P I D I ASPETTI GENERALI I Lipidi o grassi sono la riserva energetica del nostro organismo; nel corpo umano costituiscono mediamente il 17% del peso corporeo dove formano il tessuto adiposo. In generale
I L I P I D I ASPETTI GENERALI I Lipidi o grassi sono la riserva energetica del nostro organismo; nel corpo umano costituiscono mediamente il 17% del peso corporeo dove formano il tessuto adiposo. In generale
INSOLUBILI IN ACQUA E SOLUBILI NEI SOLVENTI ORGANICI NON POLARI
 LIPIDI Una delle quattro principali classi di sostanze biologicamente attive. I lipidi (la parola deriva dal greco lípos, grasso) sono costituenti delle piante e degli animali, caratterizzati da particolari
LIPIDI Una delle quattro principali classi di sostanze biologicamente attive. I lipidi (la parola deriva dal greco lípos, grasso) sono costituenti delle piante e degli animali, caratterizzati da particolari
CHETOGENESI (PRODUZIONE DI CORPI CHETONICI)
") CHETOGENESI (PRODUZIONE DI CORPI CHETONICI) Bassa glicemia (es.: digiuno) o di Diabete mellito non trattato (insufficiente captazione insulino-dipendente di glucosio) La glicolisi è inibita ma la β-ossidazione
CHETOGENESI (PRODUZIONE DI CORPI CHETONICI) Bassa glicemia (es.: digiuno) o di Diabete mellito non trattato (insufficiente captazione insulino-dipendente di glucosio) La glicolisi è inibita ma la β-ossidazione
glicogeno Glucosio 6-P Piruvato Acetil CoA Intermedi del ciclo di Krebs
 PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
>> Il malonil-coa è prodotto nel citosol a partire dall acetil-coa.
 SINTESI DEGLI ACIDI GRASSI >> AVVIENE NEL CITOSOL. Nei mammiferi principalmente negli epatociti e negli adipociti. >> per sintetizzare acidi grassi è necessario avere nel citosol: Acetil-CoA, Malonil-CoA
SINTESI DEGLI ACIDI GRASSI >> AVVIENE NEL CITOSOL. Nei mammiferi principalmente negli epatociti e negli adipociti. >> per sintetizzare acidi grassi è necessario avere nel citosol: Acetil-CoA, Malonil-CoA
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 ANABOLISMO DEI LIPIDI
 Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
I LIPIDI. Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi
 Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI Nelson Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER,
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI Nelson Cox, I PRINCIPI DI BIOCHIMICA DI LEHNINGER,
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
METABOLISMO DEI GRASSI
 Capitolo 27 METABOLISMO DEI GRASSI La maggior parte dell energia conservata in un organismo si trova nei depositi di grasso. In questi corpulenti combattenti di sumo la conservazione di energia sotto forma
Capitolo 27 METABOLISMO DEI GRASSI La maggior parte dell energia conservata in un organismo si trova nei depositi di grasso. In questi corpulenti combattenti di sumo la conservazione di energia sotto forma
Valitutti, Taddei, Kreuzer, Massey, Sadava, Hills, Heller, Berenbaum
 Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
METABOLISMO E SISTEMI ENERGETICI
 METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
Cosa sono i grassi o lipidi?
 Cosa sono i grassi o lipidi? Lipidi da LIPOS = grasso Sostanze organiche di diversa natura con una caratteristica comune: sono insolubili in acqua Importanti costituenti dei tessuti vegetali e animali
Cosa sono i grassi o lipidi? Lipidi da LIPOS = grasso Sostanze organiche di diversa natura con una caratteristica comune: sono insolubili in acqua Importanti costituenti dei tessuti vegetali e animali
METABOLISMO DEI LIPIDI
 METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
Biosintesi non ribosomiale di metaboliti peptidici bioattivi
 Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA
 CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
BIOLOGIA GENERALE. Alessandro Massolo Dip. Biologia Animale e Genetica c/o Prof. F. Dessì-Fulgheri (Via Romana 17) massolo@unifi.
 massolo@unifi.") Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE Facoltà di Scienze della Formazione Scienze della Formazione Primaria Alessandro Massolo Dip. Biologia Animale
Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE Facoltà di Scienze della Formazione Scienze della Formazione Primaria Alessandro Massolo Dip. Biologia Animale
Biosintesi degli acidi grassi
 Biosintesi degli acidi grassi NADPH 1 Acetil-CoA Carbossilasi (ACC):il primo enzima della sintesi degli acidi grassi Precursore principale della biosintesi La reazione avviene in due tappe come in tutte
Biosintesi degli acidi grassi NADPH 1 Acetil-CoA Carbossilasi (ACC):il primo enzima della sintesi degli acidi grassi Precursore principale della biosintesi La reazione avviene in due tappe come in tutte
Il metabolismo del glicogeno e la gluconeogenesi. Copyright 2013 Zanichelli editore S.p.A.
 Il metabolismo del glicogeno e la gluconeogenesi Copyright 2013 Zanichelli editore S.p.A. Capitolo 16 La demolizione del glicogeno Concetti chiave 16.1 Il glicogeno, la forma di immagazzinamento del glucosio,
Il metabolismo del glicogeno e la gluconeogenesi Copyright 2013 Zanichelli editore S.p.A. Capitolo 16 La demolizione del glicogeno Concetti chiave 16.1 Il glicogeno, la forma di immagazzinamento del glucosio,
AcetilCoA carbossilasi
 AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
La nutrizione e il metabolismo
 La nutrizione e il metabolismo 1. Le sostanze nutritive, o nutrienti 2. Definizione di metabolismo 3. Il metabolismo dei carboidrati 4. Il metabolismo dei lipidi 1 L apparato respiratorio 5. Il metabolismo
La nutrizione e il metabolismo 1. Le sostanze nutritive, o nutrienti 2. Definizione di metabolismo 3. Il metabolismo dei carboidrati 4. Il metabolismo dei lipidi 1 L apparato respiratorio 5. Il metabolismo
Il ciclo nutrizione / digiuno
 Il ciclo nutrizione / digiuno Aspetti generali La fase di assorbimento è il periodo che va dalle 2 alle 4 ore successive all assunzione di un pasto. Durante questo intervallo si verifica un transitorio
Il ciclo nutrizione / digiuno Aspetti generali La fase di assorbimento è il periodo che va dalle 2 alle 4 ore successive all assunzione di un pasto. Durante questo intervallo si verifica un transitorio
dieta vengono convertiti in composti dei corpi chetonici.
 Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Ruolo metabolico delle flavine
 Ruolo metabolico delle flavine Deidrogenasi flaviniche che trasferiscono 2 elettroni Acil CoA deidrogenasi (beta ossidazione acidi grassi) Diidrolipoildeidrogenasi (complessi piruvato dh etc.) Glutatione
Ruolo metabolico delle flavine Deidrogenasi flaviniche che trasferiscono 2 elettroni Acil CoA deidrogenasi (beta ossidazione acidi grassi) Diidrolipoildeidrogenasi (complessi piruvato dh etc.) Glutatione
Modulo 15 metabolismo dei lipidi
 Modulo 15 metabolismo dei lipidi Il metabolismo cellulare dei lipidi (AG= acido grasso) AGE= AG essenziale 7 Digestione ed assorbimento e utilizzazione dei lipidi 90% dieta triacilgliceroli 10% colesterolo,
Modulo 15 metabolismo dei lipidi Il metabolismo cellulare dei lipidi (AG= acido grasso) AGE= AG essenziale 7 Digestione ed assorbimento e utilizzazione dei lipidi 90% dieta triacilgliceroli 10% colesterolo,
Fosforilazione ossidativa
 Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Metabolismo dei lipidi
 Metabolismo dei lipidi 1 Funzione centrale dei lipidi nel metabolismo energetico 1)Assorbimento e trasporto degli acidi grassi 1) Ossidazione degli acidi grassi 2) sintesi degli acidi grassi e del colesterolo
Metabolismo dei lipidi 1 Funzione centrale dei lipidi nel metabolismo energetico 1)Assorbimento e trasporto degli acidi grassi 1) Ossidazione degli acidi grassi 2) sintesi degli acidi grassi e del colesterolo
AcetilCoA carbossilasi
 AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
BIOCHIMICA e BIOTECNOLOGIE degli ALIMENTI
 Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di
 IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
FUNZIONI DEI LIPIDI. Energetica Strutturale Precursori di biomolecole
 I LIPIDI I parte FUNZIONI DEI LIPIDI Energetica Strutturale Precursori di biomolecole CLASSIFICAZIONE E STRUTTURA DEI LIPIDI a) Lipidi di deposito: a) trigliceridi o triacilgliceroli (TAG) b) Lipidi strutturali:
I LIPIDI I parte FUNZIONI DEI LIPIDI Energetica Strutturale Precursori di biomolecole CLASSIFICAZIONE E STRUTTURA DEI LIPIDI a) Lipidi di deposito: a) trigliceridi o triacilgliceroli (TAG) b) Lipidi strutturali:
 forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
di glucosio da parte di tutte le cellule, facilitandone il trasporto transmembranario 2. aumenta l utilizzazione
 non c Ormone ipoglicemizzante, causa un forte abbassamento della glicemia (70-100 mg/100 ml) perché esalta i processi responsabili della sottrazione di glucosio dal sangue e inibisce i processi responsabili
non c Ormone ipoglicemizzante, causa un forte abbassamento della glicemia (70-100 mg/100 ml) perché esalta i processi responsabili della sottrazione di glucosio dal sangue e inibisce i processi responsabili
ENZIMI CINETICA ENZIMATICA
 ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi
 Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
1. Omogenizzazione meccanica del cibo 2. Secrezione di elettroliti 3. Secrezione di enzimi digestivi
 La dieta deve contenere, oltre all acqua, combustibile metabolico, proteine per la crescita e il ricambio delle proteine tissutali, fibre per far massa nel lume intestinale, minerali essenziali per specifiche
La dieta deve contenere, oltre all acqua, combustibile metabolico, proteine per la crescita e il ricambio delle proteine tissutali, fibre per far massa nel lume intestinale, minerali essenziali per specifiche
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) L acetil CoA carbossilasi è un enzima regolato allostericamente e covalentemente. Esso determina la velocità di biosintesi degli acidi grassi nei tessuti animali. Figura 16.7
da: Nelson & Cox (IV Ed.) L acetil CoA carbossilasi è un enzima regolato allostericamente e covalentemente. Esso determina la velocità di biosintesi degli acidi grassi nei tessuti animali. Figura 16.7
I LIPIDI. - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua
 I LIPIDI I LIPIDI - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua - ruolo fondamentale di fornitori di energia (circa 9 kcal/grammo),
I LIPIDI I LIPIDI - gruppo eterogeneo sia dal punto di vista chimico che funzionale; caratteristica comune è l insolubilità in acqua - ruolo fondamentale di fornitori di energia (circa 9 kcal/grammo),
a cura di : Gaia,Giulia, Lorenzo e Simone 2^ B ( LA MIGLIORE)
") a cura di : Gaia,Giulia, Lorenzo e Simone 2^ B ( LA MIGLIORE) I grassi nella piramide alimentare Forniscono molta ENERGIA!!! Danno gusto al cibo Trasportano alcune vitamine o LIPIDI Però se se ne mangiano
a cura di : Gaia,Giulia, Lorenzo e Simone 2^ B ( LA MIGLIORE) I grassi nella piramide alimentare Forniscono molta ENERGIA!!! Danno gusto al cibo Trasportano alcune vitamine o LIPIDI Però se se ne mangiano
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara
 Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
Mais, riso, patate, granozucchero di canna o barbabietola Latte da zucchero Polisaccaride Amido - - Disaccaride Maltosio Saccarosio Lattosio
 Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) scaricato da www.sunhope.it
 FOSFORILAZIONE OSSIDATIVA (FO) scaricato da www.sunhope.it") CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) - Decarb. ossid. piruvato - β-ossidazione acidi grassi - Vie ossidative aa - Ciclo di Krebs avvengono tutte nella matrice mitocondriale In tutte
CATENA RESPIRATORIA (CR) FOSFORILAZIONE OSSIDATIVA (FO) - Decarb. ossid. piruvato - β-ossidazione acidi grassi - Vie ossidative aa - Ciclo di Krebs avvengono tutte nella matrice mitocondriale In tutte
Degradazione degli acidi grassi
 Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Il catabolismo degli acidi grassi
 Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di
 Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di materiali per fare funzionare il nostro corpo È il carboidrato
Mangiamo perché abbiamo bisogno di energia, di materiali con cui costruire il nostro corpo, di materiali per riparare parti del nostro corpo, di materiali per fare funzionare il nostro corpo È il carboidrato
Nota dell editore Presentazione
 00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
CELLULE EUCARIOTICHE
 CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone = C 6 O 6 (H 2 O) 6
 6") Carboidrati Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone Glucosio Carboidrati modificati: membrane cellulari, acidi nucleici, Da carbonio idrato: glucosio= C 6 H 12
Carboidrati Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone Glucosio Carboidrati modificati: membrane cellulari, acidi nucleici, Da carbonio idrato: glucosio= C 6 H 12
TIAMINA (vitamina B1, aneurina)
") TIAMINA (vitamina B1, aneurina) Fonti alimentari: cereali integrali (germe e crusca), carne (maiale), lievito di birra, legumi Abbondanza nell organismo umano : 30-100 mg RDA (assunzione giornaliera di
TIAMINA (vitamina B1, aneurina) Fonti alimentari: cereali integrali (germe e crusca), carne (maiale), lievito di birra, legumi Abbondanza nell organismo umano : 30-100 mg RDA (assunzione giornaliera di
Biosintesi degli acidi grassi SCARICATO DA
 Biosintesi degli acidi grassi 1 2 3 La sintesi degli acidi grassi richiede 7 reazioni enzimatiche catalizzate dal complesso multienzimatico acido grasso sintasi e parte da acetil CoA + malonil CoA Il complesso
Biosintesi degli acidi grassi 1 2 3 La sintesi degli acidi grassi richiede 7 reazioni enzimatiche catalizzate dal complesso multienzimatico acido grasso sintasi e parte da acetil CoA + malonil CoA Il complesso
Le Biomolecole I parte. Lezioni d'autore di Giorgio Benedetti
 Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO
 Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali delle biomolecole. -
Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali delle biomolecole. -
I Composti Organici. Le Biomolecole
 I Composti Organici I composti organici sono molecole tutte contenenti carbonio. Essi comprendono. 1. composti di interesse energetico che sono gli Idrocarburi ( i derivati del petrolio), 2. composti a
I Composti Organici I composti organici sono molecole tutte contenenti carbonio. Essi comprendono. 1. composti di interesse energetico che sono gli Idrocarburi ( i derivati del petrolio), 2. composti a
La classificazione dei lipidi
 La classificazione dei lipidi Concetti chiave Le proprietà fisiche di un acido grasso sono determinate dalla sua lunghezza e dal suo grado di saturazione. I triacilgliceroli e i glicerofosfolipidi contengono
La classificazione dei lipidi Concetti chiave Le proprietà fisiche di un acido grasso sono determinate dalla sua lunghezza e dal suo grado di saturazione. I triacilgliceroli e i glicerofosfolipidi contengono
Prima di essere ossidati. gli ACIDI GRASSI devono essere attivati. (cioè legati al CoA) e poi entrare nel mitocondrio
 e poi entrare nel mitocondrio") Prima di essere ossidati gli ACIDI GRASSI devono essere attivati (cioè legati al CoA) e poi entrare nel mitocondrio L enzima acil CoA sintetasi (= acido grasso tiokinasi) sta sulla membrana mitocondriale
Prima di essere ossidati gli ACIDI GRASSI devono essere attivati (cioè legati al CoA) e poi entrare nel mitocondrio L enzima acil CoA sintetasi (= acido grasso tiokinasi) sta sulla membrana mitocondriale
 1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
Catena di Trasporto degli elettroni
 Chimica Biologica A.A. 2010-2011 Catena di Trasporto degli elettroni Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Respirazione - il catabolismo di tutti i combustibili
Chimica Biologica A.A. 2010-2011 Catena di Trasporto degli elettroni Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Respirazione - il catabolismo di tutti i combustibili
Fasi dell ossidazione degli acidi grassi
 Fasi dell ossidazione degli acidi grassi REGOLAZIONE DELLA OSSIDAZIONE DEGLI ACIDI GRASSI La velocità di trasporto nella matrice mitocondriale decide il destino (ossidazione) CARNITINA ACIL-TRANSFERASI
Fasi dell ossidazione degli acidi grassi REGOLAZIONE DELLA OSSIDAZIONE DEGLI ACIDI GRASSI La velocità di trasporto nella matrice mitocondriale decide il destino (ossidazione) CARNITINA ACIL-TRANSFERASI
ghiandole salivari bocca amilasi stomaco HCl (ph 1) pepsinogeno pepsina
 pepsinogeno pepsina") Metabolismo lipidico Digestione I lipidi della dieta sono Trigliceridi Fosfolipidi Colesterolo processi digestivi ghiandole salivari amilasi bocca HCl (ph 1) pepsinogeno pepsina stomaco processi digestivi
Metabolismo lipidico Digestione I lipidi della dieta sono Trigliceridi Fosfolipidi Colesterolo processi digestivi ghiandole salivari amilasi bocca HCl (ph 1) pepsinogeno pepsina stomaco processi digestivi
STOMACO. A livello dello stomaco si verifica un limitato assorbimento di alimenti, soprattutto di acqua, di alcuni sali, di alcool e di alcuni farmaci
 STOMACO Nel loro insieme i secreti delle ghiandole formano il succo gastrico, fortemente acido (ph circa 1). Nella mucosa gastrica sono disposte tra le ghiandole cellule mucose, che producono un muco che
STOMACO Nel loro insieme i secreti delle ghiandole formano il succo gastrico, fortemente acido (ph circa 1). Nella mucosa gastrica sono disposte tra le ghiandole cellule mucose, che producono un muco che