Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine
|
|
|
- Sabrina Spina
- 7 anni fa
- Visualizzazioni
Transcript

1
2 Energia di ossidazione Kcal/g Kj/g Grassi 9 38 Carboidrati 4 17 Proteine 4 17 Il contenuto di calorie per grammo nei grassi è più di 2 volte superiore a quello dei carboidrati e delle proteine


3 I grassi della dieta vengono assorbiti nell intestino tenue

4 Struttura molecolare di un chilomicrone
5 Formazione dei chilomicroni
6 L utilizzo degli acidi grassi come combustibili richiede tre stadi di trasformazione 1) I lipidi devono essere mobilizzati. In questo processo i triacilgliceroli vengono degradati ad acidi grassi e glicerolo,, che vengono rilasciati dal tessuto adiposo e trasportati ai tessuti che richiedono energia. 2) Gli acidi grassi devono quindi essere attivati e trasportati nei mitocondri per la degradazione. 3) Nei mitocondri gli acidi grassi vengono degradati in modo graduale in acetil CoA,, che poi viene ossidato nel ciclo dell acido citrico.
7 Il primo passaggio nell utilizzazione dei grassi come fonte di energia è l idrolisi dei trigliceridi da parte di una lipasi: L attività della lipasi è regolata dall AMP ciclico Glucagone e adrenalina attivano le lipasi

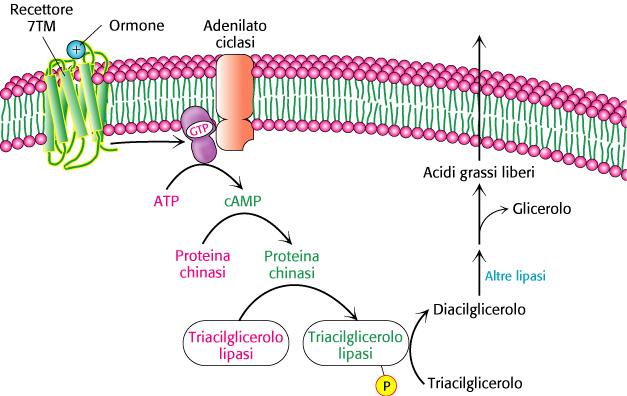
8 Mobilizzazione dei triacilgliceroli
9 Via di ingresso del glicerolo nella glicolisi
10
11
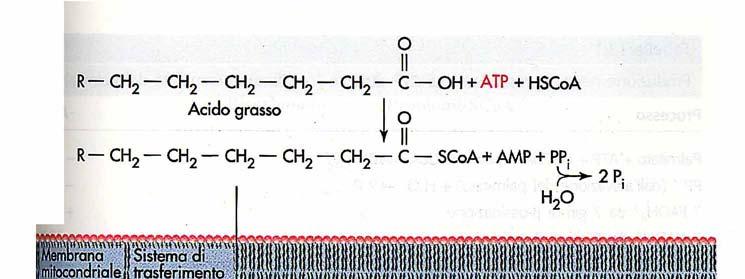
12 Prima di essere ossidati gli acidi grassi vengono legati al coenzima A L adenosina trifosfato (ATP) favorisce la formazione di un legame tioestere tra il gruppo carbossilico di un acido grasso e il gruppo sulfidrilico del CoA. La reazione avviene in due tappe:
13 L attivazione di un acido grasso avviene in due tappe Questa reazione di attivazione avviene sulla membrana mitocondriale esterna ed è catalizzata dall enzima acilcoa sintetasi. Acido grasso +CoA + ATP + H 2 O AcilCoA + AMP + 2 P i ΔG 0 = Kj/mole
14 La carnitina trasporta gli acidi grassi attivati nella matrice mitocondriale Enzimi: Carnitina aciltransferasi I e II
15

16 Ingresso degli acidi grassi nei mitocondri attraverso il trasportatore tatore acil-carnitina carnitina/carnitina

17 L acil carnitina translocasi

18 Fasi dell ossidazione degli acidi grassi
19 Formazione di un doppio legame Il doppio legame viene idratato per aggiungere un atomo di ossigeno L alcol viene ossidato a un chetone L acido grasso viene scisso dal CoA per formare AcetilCoA e una catena di acido grasso di 2 atomi più corta
20 La via di ossidazione degli acidi grassi (β( ossidazione)
21 La via di ossidazione degli acidi grassi (β( ossidazione)
22

23


24


25
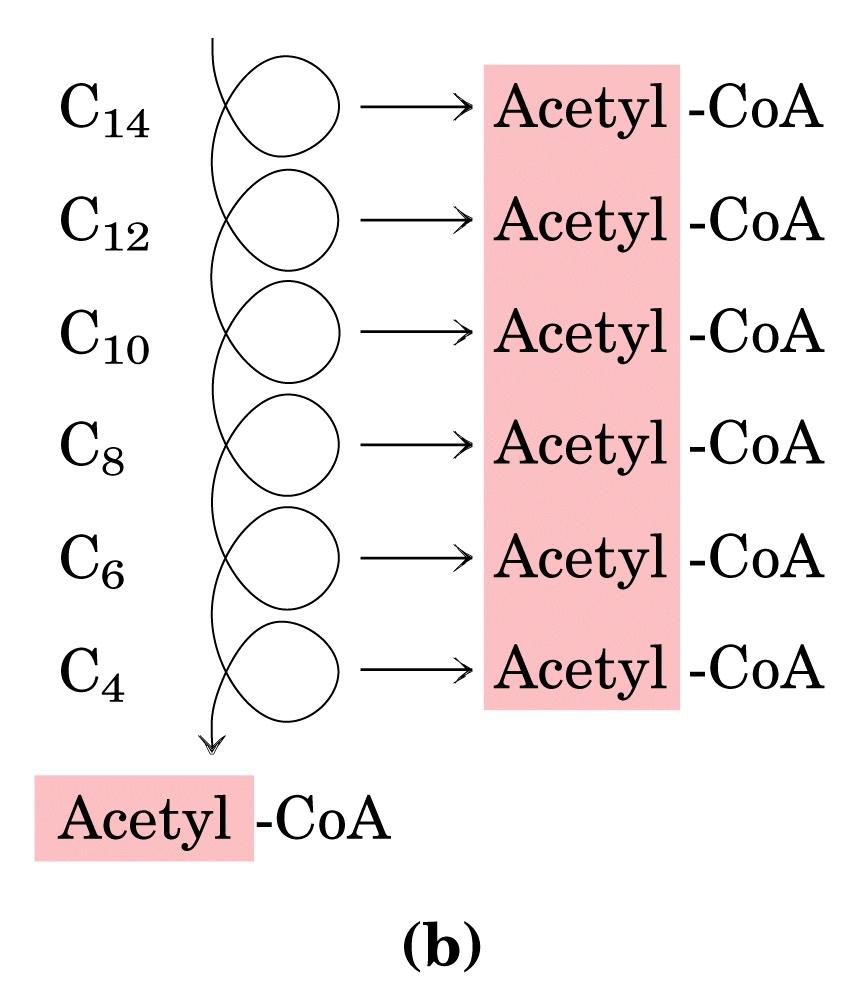
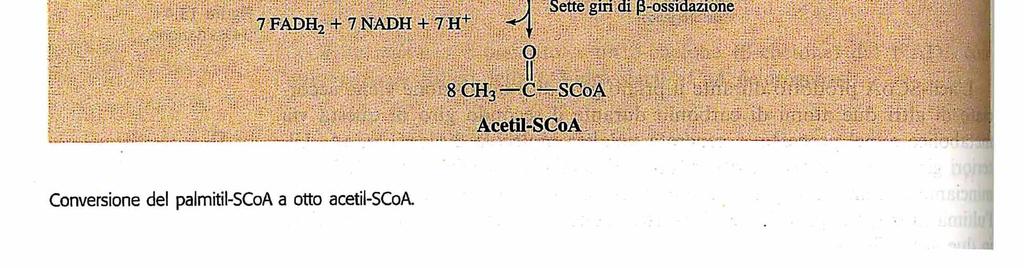
, pertanto: L ossidazione completa di una molecola di palmitato produce 106 molecole di")
26 L equivalente di 2 molecole di ATP viene consumato nell attivazione del palmitato (ATP scisso a AMP e 2 molecole di P i ), pertanto: L ossidazione completa di una molecola di palmitato produce 106 molecole di ATP

27 Gli orsi ossidano i loro grassi durante il letargo
28 Con un acido grasso con numero pari di atomi di carbonio, durante l ultima l tappa della β-ossidazione un acetoacetil-scoa è scisso in 2 Acetil-SCoA O O O CH 3 - C - CH 2 - C - SCoA + CoASH 2 CH3 - C- SCoA Acetoacetil-CoA Acetil-CoA Enzima: tiolasi La maggior parte dei lipidi presenti in natura contiene acidi grassi con un numero pari di atomi di carbonio
29 Gli acidi grassi a catena dispari sono presenti in quantità significative nei lipidi delle piante e in alcuni organismi marini Il propionil-scoa e l acetill acetil-scoa sono i prodotti finali della β-ossidazione quando viene ossidato un acido grasso a numero dispari di atomi di carbonio. O CH 3 - CH 2 O - CH 2 - CH 2 - C - SCoA CH 3 - CH 2 - C - SCoA Propionil-CoA O CH 3 - C - SCoA Acetil-CoA
30 L ossidazione degli acidi grassi a catena dispari ha bisogno di altri tre enzimi
31
32
33
34
35 Struttura del coenzima B 12
36 Formazione di un radicale 5 -deossiadenosilico5 La reazione della metilmalonil CoA mutasi comincia con la scissione omolitica del legame che unisce lo ione Co 3+ a un atomo di C del ribosio della porzione adenosinica. La scissione genera un radicale 5 deossiadenosilico e determina la riduzione del Co 3+ a Co 2+. Il ruolo del coenzima B 12 nelle migrazioni intramolecolari è quello di fungere da sorgente di radiclai liberi per l estrazione l di atomi di idrogeno. Una proprietà essenziale del coenzima è la debolezza del legame cobalto-carbonio, carbonio, la cui facile scissione genera un radicale
37 Formazione di succinil CoA mediante una reazione di riarrangiamento Il radicale libero generato dal coenzima B12 estrae un atomo di idrogeno nel riarrangiamento del metilmalonil CoA a succinil CoA
38
39 Ossidazione di un acido grasso monoinsaturo L ossidazione richiede un altro enzima, l enoil-coa isomerasi, che modifica la posizione del doppio legame convertendo l isomero cis nell isomero trans. L intermedio normale della β ossidazione.
40 Ossidazione di un acido grasso poliinsaturo L ossidazione richiede oltre all enoil enoil-coa isomerasi,, un secondo enzima ausiliario la 2,4- dienol-coa reduttasi NADPH- dipendente.
41 Ossidazione di un acido grasso poliinsaturo (b) L ossidazione richiede oltre all enoil enoil-coa isomerasi,un secondo enzima ausiliario: la 2,4-dienol dienol-coa reduttasi NADPH-dipendente dipendente. L azione combinata di questi due enzimi converte l intermedio trans-δ 2,cis-Δ - dienoil-coa in trans-δ 2 - enoil-coa, che può proseguire nella β ossidazione.
42 Per l ossidazione l di qualsiasi acido grasso polinsaturo sono sufficienti 2 enzimi addizzionali I doppi legami in posizione dispari sono convertiti dall isomerasi isomerasi; I doppi legami in posizione pari dalla riduttasi e dall isomerasi isomerasi.




43 Regolazione allosterica dell ossidazione degli acidi grassi Il malonil-coa CoA,, il primo intermedio della biosintesi citosolica degli acidi grassi a catena lunga a partire da acetil- CoA,, inibisce la carnitina aciltransferasi I, impedendo l accesso degli acil CoA alla matrice mitocondriale nei momenti di abbondanza. Regolando l acetill CoA carbossilasi,, che sintetizza malonil CoA, è possibile regolare in modo coordinato la sintesi e la degradazione degli acidi grassi Il malonil-coa aumenta quando l animale è ben rifornito di carboidrati. Regolazione ormonale: Adrenalina e glucagone mobilitazione e rilascio di acidi grassi (+ lipasi) L insulina si oppone a questo effetto
44 Nelle piante la β ossidazione non è una fonte rilevante di energia metabolica I mitocondri delle cellule delle piante sono privi degli enzimi della β-ossidazione. Durante la germinazione, i triacilgliceroli di riserva sono convertiti in glucosio e in una varietà di metaboliti essenziali. Gli acidi grassi sono ossidati a acetil-coa nei gliossisomi. L acetil-coa viene convertito attraverso il ciclo del gliossilato in precursori a quattro atomi di carbonio che entrano nella gluconeogenesi.

45 Confronto tra la β ossidazione mitocondriale e quella perossisomiale e gliossisomiale
46 Funzione della β ossidazione nella conversione dei triacilgliceroli in glucosio nei semi in germinazione.
47 Acetil - CoA + ossalacetato - ossalacetato (digiuno, diabete: l ossalacetato viene utilizzato per sintetizzare glucosio) Ciclo dell acido citrico Formazione di corpi chetonici Il destino dell acetil acetil-coa dipende dalla disponibilità di ossalacetato per formare citrato (prima reazione del ciclo dell acido citrico).
48 Corpi chetonici Acetoacetato D-β-idrossibutirrato
49 Può avvenire anche non enzimaticamente Formazione dei corpi chetonici dall acetil acetil-coa
50
51
52 Il β-idrossibutirrato come fonte di energia L acetoacetato e il D-β-idrossibutirrato sono trasportati dal sangue ai tessuti extraepatici dove sono ossidati nel ciclo dell acido citrico. L acetoacetato e il 3-idrossibutirrato 3 sono combustibili normali della respirazione e sono quantitativamente importanti come fonte energetica Il muscolo cardiaco e la corteccia renale impiegano di preferenza l acetoacetatol anziché il glucosio. Il glucosio, invece, è il principale combustibile del cervello e degli eritrociti negli individui ben nutriti che assumono una dieta bilanciata. Il cervello si adatta all utilizzo di acetoacetato durante il digiuno e nello stato diabetico
53
54
55 L acetoacetato può essere considerato una forma solubile e trasportabile di unità acetile. Gli acidi grassi vengono rilasciati dal tessuto adiposo e convertiti in unità acetile dal fegato che poi li esporta sotto forma di acetoacetato.
 esaltano la conversione dell acetil acetil-coa in acetoacetato (es.")
56 Formazione ed esportazione dei corpi chetonici. Le condizioni che determinano un aumento della gluconeogenesi (con sottrazione di ossalacetato dal ciclo dell acido citrico) esaltano la conversione dell acetil acetil-coa in acetoacetato (es.: diabete, digiuno prolungato) Elevate concentrazioni di acetoacetato nel sangue significano un abbondanza di unità acetile e determinano diminuzione della velocità della lipolisi nel tessuto adiposo.
57
58 Biosintesi degli acidi grassi
59
60 Gli acidi grassi sono sintetizzati e degradati attraverso vie differenti La sintesi degli acidi grassi avviene nel citosol; ; la degradazione nella matrice mitocondriale. Gli intermedi della sintesi sono legati covalentemente ai gruppi sulfidrilici di una proteina trasportatrice di acili (ACP); gli intermedi della degradazione sono legati al coenzima A. La molecola attivata che cede unità bicarboniose nel processo di allungamento è il malonil-acp ACP.. La reazione di allungamento è favorita dal rilascio di CO 2. L agente riducente nella sintesi degli acidi grassi è il NADPH. Negli animali superiori gli enzimi della sintesi degli acidi grassi sono uniti in una unica catena polipeptidica detta acido grasso sintasi. L allungamento della catena ad opera della acidido grasso sintasi si ferma alla formazione di palmitato (C 16 ). Ulteriori allungamenti e l inserzione l di doppi legami sono effettuati da altri sistemi enzimatici.





61 Il citrato trasporta dai mitocondri al citosol i gruppi acetile che vengono usati per la sintesi degli acidi grassi. Per ogni acetil CoA trasferito dai mitocondri al citosol viene prodotta 1 molecola di NADPH.
62 La formazione di malonil CoA è la tappa di comando nella sintesi degli acidi grassi La biosintesi degli acidi grassi inizia nel citosol con la formazione di malonil CoA catalizzata dalla acetil CoA carbossilasi,, che contiene biotina. Come nel caso degli altri enzimi che contengono biotina (es. piruvato carbossilasi) ) si forma un intermedio carbossibiotina a spese di ATP Il gruppo CO 2 attivato viene poi trasferito per formare malonil CoA biotina-enzima + ATP + HCO 3- CO 2 -biotina-enzima + ADP + P i CO 2 -biotina-enzima + acetil CoA malonil CoA + biotina-enzima
63 Gli intermedi della sintesi degli acidi grassi sono legati ad una a proteina trasportatrice di acili (ACP); il legame avviene al terminale sulfidrilico di una fosfopanteteina,, lo stesso gruppo presente nel coenzima A.
64 La proteina che trasporta gli acili
65 Le tappe della degradazione e della sintesi degli acidi grassi

66 La sequenza a quattro tappe che allunga la catena acilica nascente di due atomi di carbonio.
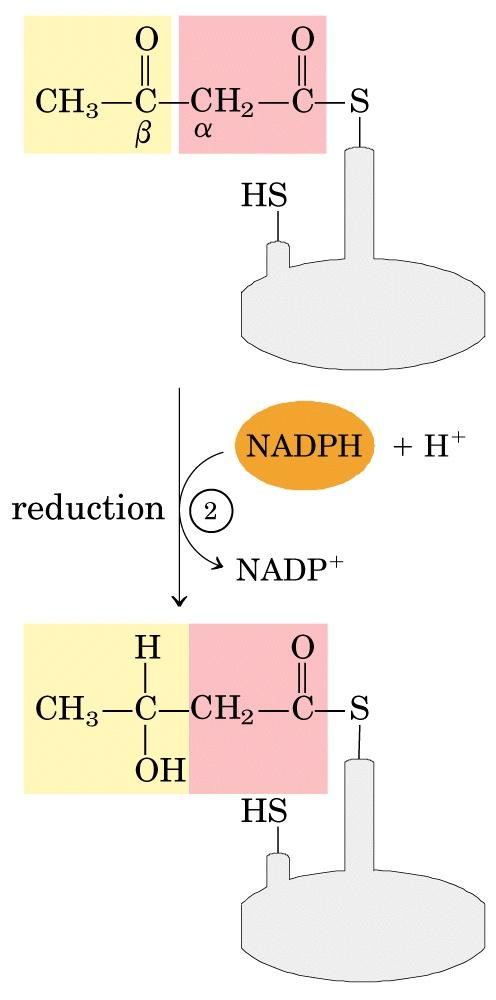
67

68
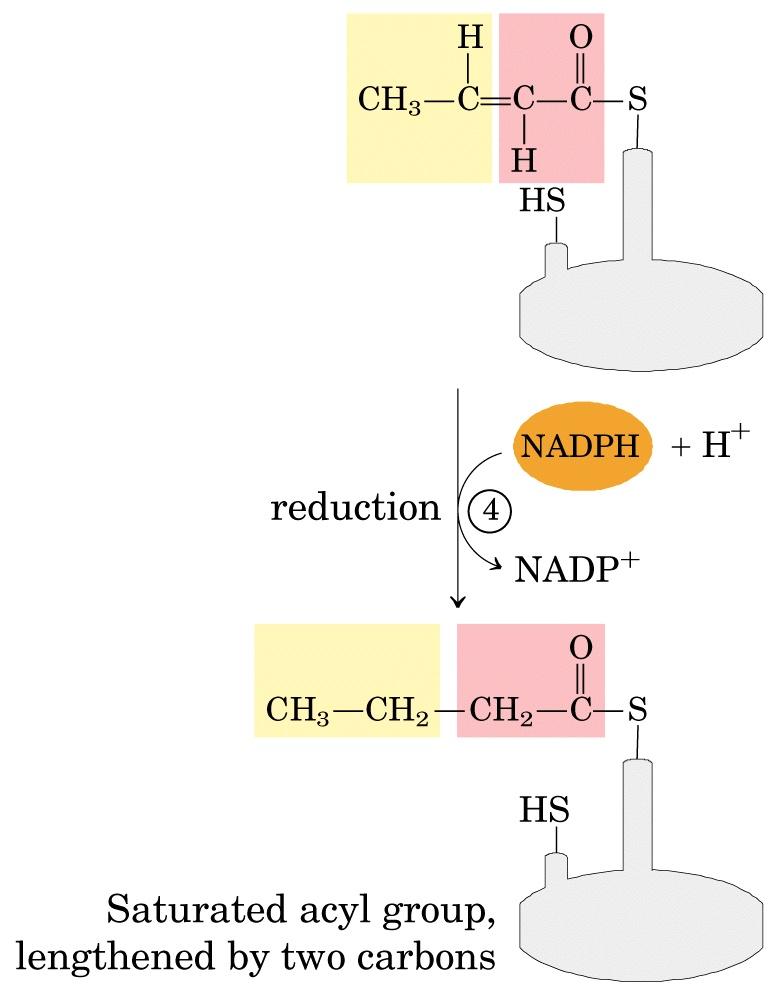
69

70 La sintesi del palmitato I cicli di allungamento si susseguono fino alla formazione di C 16 -acil-acp, che non è più substrato dell enzima enzima condensante, ma viene idrolizzato a palmitato e ACP.
71
72 Sequenza delle reazioni che avvengono durante la sintesi di un acido a grasso Il complesso dell acido grasso sintasi è mostrato in modo schematico sotto forma di un disco. Ogni segmento del disco rappresenta una delle sei attività enzimatiche del complesso. Al centro del disco si trova la proteina che trasporta gli acili (ACP), con il braccio fosfopanteteinico.
 Enzima: acetil-coa CoA-ACPACP transacetilasi (AT)")
73 Prima tappa Il gruppo acetilico dell acetil CoA viene trasferito al gruppo -SH di un residuo di cisteina della β-chetoacil sintasi (KS) Enzima: acetil-coa CoA-ACPACP transacetilasi (AT)

74 Seconda tappa Il gruppo malonile viene trasferito dal malonil CoA al gruppo -SH dell ACP Enzima: malonil-coa CoA-ACPACP transferasi (MT)
, o")
75 Condensazione Enzima: β-chetoacil-acp sitasi (KS), o enzima condensante


76 Riduzione Enzima: β-chetoacil-acp reduttasi (KR)

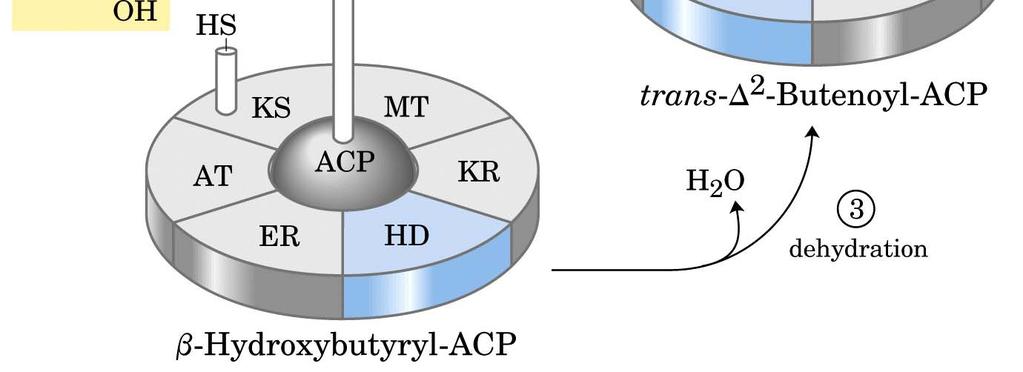
77 Deidratazione Enzima: β-idrossiacil-acp (HD) deidratasi

78 Riduzione del doppio legame Enzima: enoil-acp reduttasi (ER)
 Enzima:")
 che può trasferire gruppi acile diversi dal")
79 Traslocazione del gruppo butirrilico sulla Cys della β- chetoacil-acp sintasi (KS) Enzima: Nuovamente l l acetil-coa CoA-ACPACP transacetilasi (AT) che può trasferire gruppi acile diversi dal gruppo acetile

80 Secondo ciclo delle reazioni del complesso dell acido grasso sintasi.

81 Struttura delle varie acido grasso sintasi L acido grasso sintasi dei batteri e delle piante è un complesso formato da 7 polipeptidi diversi. Nel lievito le 7 attività sono riunite in 2 polipeptidi. Nei vertebrati tutte le attività sono su di una sola catena polipeptidica
 contiene la acetil transferasi (AT), la malonil transferasi (MT) e l enzima condensante (CE).")
82 L acido grasso sintasi animale La proteina funzionale è un dimero.. Ciascuna delle catene identiche nel dimero contiene tre domini. Il dominio 1 (blu) contiene la acetil transferasi (AT), la malonil transferasi (MT) e l enzima condensante (CE). Il dominio 2 (giallo)) contiene la proteina trasportatrice di acili (ACP), la β-chetoacil riduttasi (KR), la deidratasi (DH) e la enoil reduttasi (ER). Il dominio 3 (rosso)) contiene la tioesterasi (TE). Il gruppo flessibile di fosfopanteteina (verde)) trasporta la catena di acido grasso da un sito catalitico su una catena all altro, altro, nonché tra le catene nel dimero.
83 La stechiometria della sintesi del palmitato è: Acetil CoA + 7 malonil CoA + 14 NADPH + 20 H + palmitato + 7 CO NADP CoA + 6 H 2 O L equazione per la sintesi del malonil CoA è: 7 Acetil CoA + 7 CO ATP 7 malonil CoA + 7 ADP + 7 P i + 14 H + La stechiometria complessiva per la sintesi di palmitato è: 8 Acetil CoA + 7 ATP + 14 NADPH + 6 H + palmitato + 14 NADP CoA + 6 H 2 O + 7 ADP + 7 P i


84 Il sistema di trasporto dei gruppi acetilici dai mitocondri al citosol,, per la sintesi degli acidi grassi L acetato viene trasportato fuori dai mitocondri sotto forma di citrato
85 Il NADPH è generato dalla via del pentoso fosfato e dall enzima malico
86 La acetil CoA carbossilasi svolge un ruolo essenziale nella regolazione del metabolismo degli acidi grassi. Acetil CoA carbossilasi Acetil CoA + citrato - palmitoil CoA + insulina - glucagone Malonil CoA Acidi grassi Elevati livelli di citrato indicano che unità a due atomi di carbonio e ATP sono disponibili per la sintesi degli acidi grassi. ATP inibisce isocitrato deidrogenasi = citrato
87 Regolazione della sintesi degli acidi grassi
88

89
90
91
92
ANABOLISMO DEI LIPIDI
 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
Cap.21 ANABOLISMO DEI LIPIDI
 Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Degradazione degli acidi grassi
 Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico 2014-15
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2014-15 Lezione 16 Degradazione dei lipidi
Idratazione (introduciamo un gruppo OH sul Cβ) per mezzo della enoil-coa idratasi
 per mezzo della enoil-coa idratasi") β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
β-ossidazione di un Ac. Grasso Saturo con n PARI di atomi di carbonio Deidrogenazione FAD-dipendente del legame Cα- Cβ per mezzo della Acil-CoA deidrogenasi che è legata alla membrana mitocondriale interna)
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) L acetil CoA carbossilasi è un enzima regolato allostericamente e covalentemente. Esso determina la velocità di biosintesi degli acidi grassi nei tessuti animali. Figura 16.7
da: Nelson & Cox (IV Ed.) L acetil CoA carbossilasi è un enzima regolato allostericamente e covalentemente. Esso determina la velocità di biosintesi degli acidi grassi nei tessuti animali. Figura 16.7
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
I LIPIDI. Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi
 Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
Metabolismo Lipidi I LIPIDI Circa il 95% dei lipidi introdotti con la dieta è rappresentato da trigliceridi In media il 40% o anche più dell energia richiesta giornalmente dall uomo nei paesi industrializzati
BIOCHIMICA e BIOTECNOLOGIE degli ALIMENTI
 Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2015-16 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI
 METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
METABOLISMO DEGLI ACIDI GRASSI Lipidi introdotti con gli alimenti Mobilizzazione dei trigliceridi di riserva IDROLIZZATI DA LIPASI IN GLICEROLO E ACIDI GRASSI MOBILIZZAZIONE DEI LIPIDI DI RISERVA Condizioni
Biosintesi degli acidi grassi SCARICATO DA
 Biosintesi degli acidi grassi 1 2 3 La sintesi degli acidi grassi richiede 7 reazioni enzimatiche catalizzate dal complesso multienzimatico acido grasso sintasi e parte da acetil CoA + malonil CoA Il complesso
Biosintesi degli acidi grassi 1 2 3 La sintesi degli acidi grassi richiede 7 reazioni enzimatiche catalizzate dal complesso multienzimatico acido grasso sintasi e parte da acetil CoA + malonil CoA Il complesso
Biosintesi dei carboidrati
 Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
METABOLISMO DEI LIPIDI
 METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
METABOLISMO DEI LIPIDI STRUTTURA DEI TRIACILGLICEROLI -Molecole non polari (grassi neutri) -insolubili in H 2 O O= R-C-O - + R -OH O= R-C-O-R + H 2 O Acidi grassi: acidi carbossilici con catena idrocarburica
Il catabolismo degli acidi grassi
 Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Il catabolismo degli acidi grassi I trigliceridi sono la principale fonte di energia Il processo a 4 tappe di ossidazione degli acidi grassi a catena lunga ad acetil-coa, noto come β-ossidazione, è una
Lattato. Glicolisi. Piruvato
 Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
GLUCOSIO. Vena porta DIGESTIONE DEI DISACCARIDI. Enzimi parete intestinale
 DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
Biosintesi degli acidi grassi
 Biosintesi degli acidi grassi La capacità degli organismi di biosintetizzare diverse molecole lipidiche è essenziale Le vie metaboliche di biosintesi dei lipidi sono endoergoniche e riduttive, necessitano
Biosintesi degli acidi grassi La capacità degli organismi di biosintetizzare diverse molecole lipidiche è essenziale Le vie metaboliche di biosintesi dei lipidi sono endoergoniche e riduttive, necessitano
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale
 Stechiometria della reazione finale") Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
Cap.19. Ciclo di Krebs. o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico
 Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia.
 La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di
 IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
AcetilCoA carbossilasi
 AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
AcetilCoA carbossilasi
 AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
AcetilCoA carbossilasi Acido grasso sintasi (FAS) (FAS II) (FAS I) Proteina trasportatrice di acili (ACP) 7 Acetil-CoA + 7 CO 2 + 7ATP 7 malonil-coa + 7ADP + 7 Pi Acetil-CoA + 7 malonil-coa + 14
Modulo 14 Il ciclo di Krebs
 Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
METABOLISMO LIPIDICO
 METABOLISMO LIPIDICO I LIPIDI FUNZIONI GENERALI DEI LIPIDI Ø Funzione energetica: forniscono 9 Kcal/gr Ø Funzione ormonale (ormoni steroidei) e di messaggeri intracellulari Ø Funzione termica: mantenimento
METABOLISMO LIPIDICO I LIPIDI FUNZIONI GENERALI DEI LIPIDI Ø Funzione energetica: forniscono 9 Kcal/gr Ø Funzione ormonale (ormoni steroidei) e di messaggeri intracellulari Ø Funzione termica: mantenimento
INTRODUZIONE AL METABOLISMO. dal gr. metabolè = trasformazione
 INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
LEZIONE DEL 15/05/2017 PRIMA DELLA BETA OSSIDAZIONE L'INTERVENTO DEL TRASPORTATORE
 LEZIONE DEL 15/05/2017 PRIMA DELLA BETA OSSIDAZIONE L'INTERVENTO DEL TRASPORTATORE Dato che un acido grasso non può attraversare direttamente la prima e la seconda membrana usufruirà di una CARNITINA ACIL
LEZIONE DEL 15/05/2017 PRIMA DELLA BETA OSSIDAZIONE L'INTERVENTO DEL TRASPORTATORE Dato che un acido grasso non può attraversare direttamente la prima e la seconda membrana usufruirà di una CARNITINA ACIL
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
BETA OSSIDAZIONE DEGLI ACIDI GRASSI
 Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
Formazione dei chilomicroni
 Formazione dei chilomicroni I triacilgliceroli forniscono il combustibile per i lunghi voli migratori del Piviere minore. I triacilgliceroli sono riserve di energia molto concentrate essendo ridotti ed
Formazione dei chilomicroni I triacilgliceroli forniscono il combustibile per i lunghi voli migratori del Piviere minore. I triacilgliceroli sono riserve di energia molto concentrate essendo ridotti ed
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico
 Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 31 Lipidi I lipidi si dividono in due gruppi principali: I gruppo: comprende composti a catena aperta con una testa polare
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 31 Lipidi I lipidi si dividono in due gruppi principali: I gruppo: comprende composti a catena aperta con una testa polare
METABOLISMO DEGLI ACIDI GRASSI
 METABLISM DEGLI ACIDI GRASSI glucosio piruvato Ac-CoA NAD H ACIDI GRASSI: composti che contengono una catena idrocarburica (idrofobica) ed un gruppo terminale carbossilico (polare). Sono componenti o precursori
METABLISM DEGLI ACIDI GRASSI glucosio piruvato Ac-CoA NAD H ACIDI GRASSI: composti che contengono una catena idrocarburica (idrofobica) ed un gruppo terminale carbossilico (polare). Sono componenti o precursori
Regolazione del Ciclo di Krebs 8 reazioni
 Regolazione del Ciclo di Krebs 8 reazioni Stechiometria del Ciclo Acetil-CoA + 3NAD + + FAD + GDP + Pi + 2H 2 O 2 CO 2 + HS-CoA + 3NADH + 2H + + FADH2 + GTP (GTP ATP) 1 NADH 2,5 ATP 1 FADH 2 1,5 ATP Per
Regolazione del Ciclo di Krebs 8 reazioni Stechiometria del Ciclo Acetil-CoA + 3NAD + + FAD + GDP + Pi + 2H 2 O 2 CO 2 + HS-CoA + 3NADH + 2H + + FADH2 + GTP (GTP ATP) 1 NADH 2,5 ATP 1 FADH 2 1,5 ATP Per
Il catabolismo dei lipidi e l ossidazione degli acidi grassi
 Il catabolismo dei lipidi e l ossidazione degli acidi grassi Nella maggior parte degli organismi i lipidi si trovano principalmente nella forma di TRIACILGLICEROLI (trigliceridi) In molte piante una grossa
Il catabolismo dei lipidi e l ossidazione degli acidi grassi Nella maggior parte degli organismi i lipidi si trovano principalmente nella forma di TRIACILGLICEROLI (trigliceridi) In molte piante una grossa
Carboidrati! Oligosaccaridi Polisaccaridi. B) Carboidrati complessi
 Carboidrati complessi") Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI
 METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
LIPIDI COMPLESSI E LIPOPROTEINE
 LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
LIPIDI COMPLESSI E LIPOPROTEINE Principali lipidi assunti con la dieta Fosfolipidi e colesterolo (membrane) Triacilgliceroli (olii e grassi) Le cellule importano ACIDI GRASSI e GLICEROLO SATURI MONOINSATURI
Catabolismo dei lipidi
 Catabolismo dei lipidi Il 90% dei lipidi è costituito da trigliceridi o triacilgliceroli o grassi neutri! 5 25 % peso corporeo dei mammiferi! E la fonte primaria di riserva di energia degli animali! Nel
Catabolismo dei lipidi Il 90% dei lipidi è costituito da trigliceridi o triacilgliceroli o grassi neutri! 5 25 % peso corporeo dei mammiferi! E la fonte primaria di riserva di energia degli animali! Nel
Regolazione ormonale del metabolismo energetico. Lezione del 11 marzo 2014
 Regolazione ormonale del metabolismo energetico Lezione del 11 marzo 2014 Effetti fisiologici e metabolici dell adrenalina: preparazione all azione EFFETTO IMMEDIATO EFFETTO COMPLESSIVO Effetto fisiologico
Regolazione ormonale del metabolismo energetico Lezione del 11 marzo 2014 Effetti fisiologici e metabolici dell adrenalina: preparazione all azione EFFETTO IMMEDIATO EFFETTO COMPLESSIVO Effetto fisiologico
Gluconeogenesi sintesi di glucosio da precursori non glucidici
 GLUCONEOGENESI Gluconeogenesi sintesi di glucosio da precursori non glucidici Precursori: Lattato, ossalacetato, glicerolo Amminoacidi (scheletro carbonioso) Usa le reazioni glicolitiche in direzione
GLUCONEOGENESI Gluconeogenesi sintesi di glucosio da precursori non glucidici Precursori: Lattato, ossalacetato, glicerolo Amminoacidi (scheletro carbonioso) Usa le reazioni glicolitiche in direzione
Cap.16 GLICOLISI. Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP H 2 O 2 NAD + 2 NADH + 2H +
 Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
INSULINA RECERE REC INS T U T L O INA RE OR
 INSULINA RECERE REC INSULINA TTORE OR Lipidi Sono un gruppo eterogeneo di composti con una caratteristica comune: l isolubilità in acqua Lipidi di riserva (triacilgliceroli o trigliceridi e cere) Lipidi
INSULINA RECERE REC INSULINA TTORE OR Lipidi Sono un gruppo eterogeneo di composti con una caratteristica comune: l isolubilità in acqua Lipidi di riserva (triacilgliceroli o trigliceridi e cere) Lipidi
Ruolo del fegato nella nutrizione
 Ruolo del fegato nella nutrizione Centro di distribuzione dei nutrienti per tutti gli altri organi Annulla le fluttuazioni del metabolismo determinate dall assunzione intermittente del cibo Funge da deposito
Ruolo del fegato nella nutrizione Centro di distribuzione dei nutrienti per tutti gli altri organi Annulla le fluttuazioni del metabolismo determinate dall assunzione intermittente del cibo Funge da deposito
IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI
 GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la
 Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
NADH e NADPH. ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente)
 della cellula NADH, NADPH = 2 moneta (potere riducente)") NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
Capitolo 6 La respirazione cellulare
 Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
GLICOLISI ADP ATP. Condizioni aerobiche. Condizioni anaerobiche ATP
 GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
 1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
1. Cosa si intende per steatorrea? 2. Indica la sede di formazione delle seguenti lipoproteine: a) HDL (fegato-intestino) b) LDL (dal metabolismo intravasale delle VLDL) c) VLDL (fegato) d) Chilomicroni
INTRODUZIONE AL METABOLISMO
 INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
Metabolismo. Il metabolismo è l insieme delle migliaia di reazioni chimiche che avvengono in tutte le cellule.
 Metabolismo Il metabolismo è l insieme delle migliaia di reazioni chimiche che avvengono in tutte le cellule. Le reazioni chimiche che compongono il metabolismo sono organizzate nelle : VIE METABOLICHE
Metabolismo Il metabolismo è l insieme delle migliaia di reazioni chimiche che avvengono in tutte le cellule. Le reazioni chimiche che compongono il metabolismo sono organizzate nelle : VIE METABOLICHE
Fosforilazione a livello del substrato
 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
prodotto finale della glicolisi
 Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
6 H 2. con G=-686 kcal/mole di H 12 O 6 O + 6 CO O 2. glucosio La respirazione avviene in tre stadi principali; ognuno di questi
 La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore)
") Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
scaricato da www.sunhope.it
 CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
IL METABOLISMO. Dal carbonio agli OGM Capitolo 2
 IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
Attenzione : lunedì 29 aprile NON ci sarà lezione
 Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
catabolismo anabolismo
 Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
La fotosintesi e la respirazione cellulare
 La fotosintesi e la respirazione cellulare 1 La fotosintesi immagazzina energia e rilascia ossigeno L ATP è una molecola antica e universale, sintetizzata da tutti gli organismi viventi. L ATP viene utilizzata
La fotosintesi e la respirazione cellulare 1 La fotosintesi immagazzina energia e rilascia ossigeno L ATP è una molecola antica e universale, sintetizzata da tutti gli organismi viventi. L ATP viene utilizzata
Regolazione dell anabolismo lipidico (Acidi grassi, trigliceridi e colesterolo)
") Regolazione dell anabolismo lipidico (Acidi grassi, trigliceridi e colesterolo) Biosintesi degli acidi grassi Ac CoA MAT ACP AG sintasi KS condensazione KS MAT: malonyl/acetyl transferase MAT Malonil CoA
Regolazione dell anabolismo lipidico (Acidi grassi, trigliceridi e colesterolo) Biosintesi degli acidi grassi Ac CoA MAT ACP AG sintasi KS condensazione KS MAT: malonyl/acetyl transferase MAT Malonil CoA
Seminario. Ciclo di Krebs 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A
 Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Catabolismo dei carboidrati Glicolisi
 Catabolismo dei carboidrati Glicolisi Principali vie di utilizzo del Glucosio L estrazione dell energia dai composti organici può essere divisa in tre stadi Metabolismo del glucosio C 6 H 12 O 6 + 6O 2
Catabolismo dei carboidrati Glicolisi Principali vie di utilizzo del Glucosio L estrazione dell energia dai composti organici può essere divisa in tre stadi Metabolismo del glucosio C 6 H 12 O 6 + 6O 2
Utilizzazione metabolica dei nutrienti
 Utilizzazione metabolica dei nutrienti Il rifornimento di substrati è discontinuo Fase postprandiale utilizzazione dei nutrienti esogeni e messa in riserva Fase postassorbitiva (digiuno) mobilizzazione
Utilizzazione metabolica dei nutrienti Il rifornimento di substrati è discontinuo Fase postprandiale utilizzazione dei nutrienti esogeni e messa in riserva Fase postassorbitiva (digiuno) mobilizzazione
Il FEGATO Cardarera, cap.3
 Il FEGATO Cardarera, cap.3 2.5% della massa corporea 1,6-1.8 Kg nel maschio 1.3-1.4 Kg nella femmina Tramite la vena porta, è il primo organo a ricevere il sangue arricchito di nutrienti assorbiti dall
Il FEGATO Cardarera, cap.3 2.5% della massa corporea 1,6-1.8 Kg nel maschio 1.3-1.4 Kg nella femmina Tramite la vena porta, è il primo organo a ricevere il sangue arricchito di nutrienti assorbiti dall
Utilizzo del glucosio: la glicolisi
 Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
AGISCONO NEL SISTEMA NAVETTA 2 ISOFORME DELL ENZIMA: UNA FORMA MITOCONDRIALE ( MEMBRANA MIT. INTERNA) FAD-dipendente
 FAD-dipendente") METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
L ossidazione completa del glucosio da parte dell O 2. può essere suddivisa in due semi-reazioni
 L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
Connessioni. Le cellule cardiache sono unite tra loro da parti più ispessite del sarcolemma dette: Gap - junction
 Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA
 I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
Come le cellule traggono energia dal cibo: produzione di ATP
 Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Scheletrico Striato. Cardiaco Striato. Liscio
 IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
Indice generale 1. Introduzione alla biochimica 2. Acqua, ph, acidi, basi, sali, tamponi
 Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
2 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA
 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
scaricato da 1
 scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
METABOLISMO LIPIDICO. Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva. Lipidi introdotti con gli alimenti.
 METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
METABOLISMO LIPIDICO cistifellea Lipidi introdotti con gli alimenti Gli ac. grassi sono ossidati o incorporati in trigliceridi di riserva inestino I lipidi sono emulsionati dai sali biliari e resi accessibili
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
Scheletrico Striato. Cardiaco Striato. Liscio
 IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
dotto di Wirsung che si riversa nella papilla maggiore del duodeno;
 Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Metabolismo dei lipidi Abbiamo già visto che il metabolismo energetico con produzione di ATP, avviene in seguito alla demolizione di zuccheri (glicolisi), lipidi, e in piccolissima percentuale delle proteine.
Scheletrico Striato. Cardiaco Striato. Liscio
 IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
IL METABOLISMO DEL TESSUTO MUSCOLARE Scheletrico Striato La maggior parte della massa muscolare corporea Volontario Cardiaco Striato Cuore Involontario Liscio Non striato Visceri e vasi sanguigni Involontario
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI
 LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
I processi metabolici cellulari
 I processi metabolici cellulari 1 Metabolismo E l insieme delle reazioni chimiche che avvengono in una cellula o, più in generale, in qualsiasi organismo. Le sostanze coinvolte in tali reazioni sono dette
I processi metabolici cellulari 1 Metabolismo E l insieme delle reazioni chimiche che avvengono in una cellula o, più in generale, in qualsiasi organismo. Le sostanze coinvolte in tali reazioni sono dette
Prof. Maria Nicola GADALETA DISPENSA N. 23
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Metabolismo glucidico
 Metabolismo glucidico Glicogeno glicogenolisi Riboso-5P (NADPH) Colesterolo glicogenosintesi shunt pentoso fosfati Glucoso Glucoso-6P glicolisi Piruvato gluconeogenesi CO 2 -ossidazione Trigliceridi Acidi
Metabolismo glucidico Glicogeno glicogenolisi Riboso-5P (NADPH) Colesterolo glicogenosintesi shunt pentoso fosfati Glucoso Glucoso-6P glicolisi Piruvato gluconeogenesi CO 2 -ossidazione Trigliceridi Acidi
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
Anatomia biochimica di un mitocondrio
 Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più