Metabolismo dei composti del carbonio
|
|
|
- Lucia Campo
- 5 anni fa
- Visualizzazioni
Transcript

1 rof. Giorgio artor Metabolismo dei composti del carbonio arte 1 Degradazione ossidativa Copyright by Giorgio artor. All rights reserved. B01 - Versione Apr-18 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma di nucleotidi trifosfati, equivalenti riducenti, ecc.) e di utilizzarla (per produrre legami chimici, energia, calore, ecc.) Tutta la chimica dei viventi! B05 - v gsartor ucleotidi e coenzimi 2 1
2 Metabolismo È la somma delle reazioni EZIMATICE che avvengono in una cellula, permette di: Estrarre energia dai composti organici o dalla luce solare, Convertire i nutrienti in unità costitutive di macromolecole, Legare insieme le unità costitutive per formare macromolecole CATABLIM AABLIM B05 - v gsartor ucleotidi e coenzimi 3 Metabolismo utrienti Carboidrati Grassi roteine Macromolecole Carboidrati Grassi roteine Calore CATABLIM (demolizione) AD AD + AD + AT AD AD AABLIM (sintesi) rodotti finali semplici C 2, 2, 3 Luce rodotti Intermedi monosaccaridi, aminoacidi, composti carbonilici, amine, nucleotidi FTITEI B05 - v gsartor ucleotidi e coenzimi 4 2
 Composti organici Luce Composti organici Chemioautotrofi ( 2, Reazioni Fe,, batteri nitrificanti) C 2 redox 2, 2, 4+, 2-, Fe ++, Mn ++ Chemioeterotrofi")
3 Fonti di carbonio ed energia rganismo Fonte di carbonio Fonte di energia Donatore di elettroni Fotoautotrofi (piante, alghe verdi, cianobatteri fotosintetici) C 2 Luce 2, 2,, altri inorganici Fotoeterotrofi (rodobatteri non sulfurei) Composti organici Luce Composti organici Chemioautotrofi ( 2, Reazioni Fe,, batteri nitrificanti) C 2 redox 2, 2, 4+, 2-, Fe ++, Mn ++ Chemioeterotrofi (Animali, microrganismi, tessuti di piante non fotosintetici) Composti organici Reazioni redox 2, composti organici B05 - v gsartor ucleotidi e coenzimi 5 Fonti di carbonio ed energia D-Glucoso + 2 Luce ETERTRFI FTTRFI CaC 3 2, C 2 Ca ++ B05 - v gsartor ucleotidi e coenzimi 6 3
4 Metabolismo del carbonio.. Carbonio < +4 D-Glucoso +.. ssigeno = 0 2 Luce Cellule Cellule fotosintetiche 2, C 2.. ssigeno = -2.. Carbonio = +4 B05 - v gsartor ucleotidi e coenzimi 7 Ciclo biogeochimico del carbonio hν FTITEI Zuccheri Metabolismo degli organismi fotosintetici e autotrofi non fotosintetici C 2 Macromolecole Metabolismo organismi eterotrofi Metabolismo organismi degradatori (batteri e funghi) Macromolecole B05 - v gsartor ucleotidi e coenzimi 8 4

5 ssigeno Aerobi Usano l ossigeno come accettore finale degli elettroni Aerobi obbligati Usano L l ossigeno come accettore finale degli elettroni Anaerobi facoltativi ossono usare altri accettori di elettroni Anaerobi obbligati possono usare l ossigeno come accettore di elettroni B05 - v gsartor ucleotidi e coenzimi 9 Ciclo dell azoto L azoto in natura si trova in forma ossidata come come nitrato ( 3- ) e come gas ( 2 ), er essere utilizzato deve essere convertito in forma ridotta ( 4+ ) Ciò può avvenire in aerobiosi o anaerobiosi. B05 - v gsartor ucleotidi e coenzimi 10 5
6 Ciclo dell azoto AERBII ITRATAZIE (Batteri nitratatori) 2 - Assimilazione del nitrato ITRIFICAZIE (Batteri nitrificatori) Azoto organico 2 - DEITRIFICAZIE (Batteri denitrificanti) 2 Respirazione del nitrato AAERBII 2 FIAZIE DELL'AZT (Batteri azotofissatori) B05 - v gsartor ucleotidi e coenzimi 11 Metabolismo dell azoto iante superiori Aminoacidi Animali superiori itrati Batteri itriti Batteri fissatori dell azoto 2 atmosferico 3, Urea Batteri nitrificanti B05 - v gsartor ucleotidi e coenzimi 12 6
7 EZIMI Classificazione degli enzimi ingolo enzima Ureasi Complesso enzimatico Complesso piruvato deidrogenasi B05 - v gsartor ucleotidi e coenzimi 13 Classificazione gerarchica degli enzimi gni enzima viene classificato a secondo della reazione che catalizza. Viene classificato con un numero: ECX.Y.Z.T X = classe Y = sottoclasse Z = sotto-sottoclasse T = numero dell enzima nella sotto-sottoclasse B05 - v gsartor ucleotidi e coenzimi 14 7
8 Classificazione gerarchica degli enzimi Classi: 1. ssidoreduttasi Catalizzano una reazione redox. 2. Transferasi Catalizzano il trasferimento di un gruppo da una molecola ad un altra: X-Y + Z X-Z + Y le chinasi trasferiscono un gruppo fosfato. 3. Idrolasi Catalizzano la scissione idrolitica di legami C=, C-, C-C, --, 4. Liasi Catalizzano scissioni di legami con meccanismi diversi dalle ssidoreduttasi e dalle Idrolasi. 5. Isomerasi Catalizzano modificazioni geometriche. 6. Ligasi (intetasi) Catalizzano l unione di due molecole accoppiata al consumo di AT o di un altro nucleotide trifosfato. B05 - v gsartor ucleotidi e coenzimi 15 Classificazione gerarchica degli enzimi er esempio: Alcool + AD + Aldeide o chetone + AD ome comune: alcool deidrogenasi ome sistematico: alcool:ad + ossidoreduttasi EC umero dell enzima nella sotto-sottoclasse La sotto-sottoclasse con AD + o AD + come accettori La sottoclasse Agiscono sul gruppo di donatori C- La classe - ssidoreduttasi B05 - v gsartor ucleotidi e coenzimi 16 8
9 B05 - v gsartor ucleotidi e coenzimi 17 B05 - v gsartor ucleotidi e coenzimi 18 9
10 B05 - v gsartor ucleotidi e coenzimi 19 B05 - v gsartor ucleotidi e coenzimi 20 10
11 Isoenzimi Questa classificazione riguarda gli enzimi in quanto proteine ma in quanto CATALIZZATRI La classificazione riguarda quindi gli enziomi che catalizzano una reazione. roteine diverse (con diversa struttura primaria) che catalizzano la stessa reazione, sono IEZIMI ono le forme multiple dovute a differenze, determinate geneticamente, di struttura primaria ono anche isoenzimi quelle proteine che svolgono la stessa funzione ma sono geneticamente indipendenti, per esempio: EC malato deidrogenasi Citosolica (codificata dal DA nucleare) Mitocondriale (codificata dal DA mitocondriale) B05 - v gsartor ucleotidi e coenzimi 21 Enzimi ermettono l accoppiamento di reazioni chimiche: ello stesso posto ello stesso momento B05 - v gsartor ucleotidi e coenzimi
12 Reazioni accoppiate A B G ' = 4 kcal mole -1 Consideriamo l equilibrio [B] eq K eq = = 10 [A] eq - G ' RT = (a 25 C) e la reazione è accoppiata all idrolisi di AT attraverso un enzima: A + AT + 2 B + AD + i + + G ' = -3.3 kcal mole -1 [B] eq [AD] eq [i] eq K eq = [A] eq [AT] eq = [B] eq [A] eq [AT] eq = K eq [AD] eq [i] eq B05 - v gsartor ucleotidi e coenzimi Reazioni accoppiate ella cellula il rapporto: [AT] [AD] [i] ~ 500 [B] eq [A] eq = x 500 = ~ In presenza di idrolisi di AT la Keq di una reazione può aumentare di 10 8 volte; e l idrolisi di una mole di AT (-7.3 kcal) non bastano più moli di AT possono essere impiegate. B05 - v gsartor ucleotidi e coenzimi
13 Reazioni accoppiate A e B on sono necessariamente due composti chimici ma possono essere: differenti conformazioni di una molecola; differenti concentrazioni di uno ione ai lati di una membrana, ecc stati iniziali e finali di una TRAFRMAZIE! B05 - v gsartor ucleotidi e coenzimi Coenzimi 13
, per esempio con l adenosina, si")
14 ucleosidi Il legame si forma tra il C1 dello zucchero e con l atomo di azoto non coinvolto nella tautomeria. i formano i nucleosidi, in questo caso l adenosina. 2 5' C 2 4' 3' 1' 2' 2 C 2 2 B05 - v gsartor ucleotidi e coenzimi ucleotidi La funzione alcolica primaria rimasta libera può essere esterificata con l acido fosforico (fosfoestere), per esempio con l adenosina, si ottiene una molecola idrosolubile e carica. Adenosinmonofosfato (AM) 2 5' 4' 1' 3' 2' 2 5' 3' 1' B05 - v gsartor ucleotidi e coenzimi
15 ucleotidi iù in generale: BAE AZTATA BAE AZTATA 5' 3' 1' ' 3' 1' B05 - v gsartor ucleotidi e coenzimi Deossinucleotidi iù in generale: BAE AZTATA BAE AZTATA 5' 3' 1' ' 3' 1' B05 - v gsartor ucleotidi e coenzimi

.")
16 ucleotidi ciclici Anche la funzione alcolica in 3 può essere esterificata con una funzione acida libera dello stesso acido fosforico, si ottengono i nucleotidi ciclici: cam 2 5' 3' B05 - v gsartor ucleotidi e coenzimi ucleotidi e acidi nucleici Il legame può anche avvenire tra nucleotidi diversi attraverso esterificazioni in 3 e 5 per formare catene polinucleotidiche (acidi nucleici). ucleotide 2 5' Deossinucleotide 2 5' 2 2 3' 3' ucleotide Deossinucleotide Acido ribonucleico (RA) Acido deossiribonucleico (DA) B05 - v gsartor ucleotidi e coenzimi

 e i trifosfonucleotidi (AT) 5' 5' 2 3' 3' 1' 1'")
17 ucleotidi e acidi nucleici RA DA B05 - v gsartor ucleotidi e coenzimi ucleotidi liberi 2 I monofosfonucleotidi hanno la possibilità di essere ulteriormente fosforilati per formare i difosfonucleotidi (AD) e i trifosfonucleotidi (AT) 5' 5' 2 3' 3' 1' 1' B05 - v gsartor ucleotidi e coenzimi

18 ucleotidi liberi I monofosfonucleotidi hanno la possibilità di essere ulteriormente fosforilati per formare i difosfonucleotidi (AD) e i trifosfonucleotidi (AT) B05 - v gsartor ucleotidi e coenzimi ucleotidi liberi I legami che tengono insieme i residui di fosfato sono legami di tipo anidridico B05 - v gsartor ucleotidi e coenzimi
19 AD e AT Legami di tipo anidridico Legami di tipo fosfoestereo 2 2 AD AT B05 - v gsartor ucleotidi e coenzimi A cosa servono i nucleotidi Trasporto di energia AT (GT), AD, AM, egnali intracellulari cam Trasporto di equivalenti ridotti AD + /AD, AD + /AD FAD/FAD 2 Trasporto di acili B05 - v gsartor ucleotidi e coenzimi

20 B05 - v gsartor ucleotidi e coenzimi Energia AT 2 G 0 = -7.3 kcal mole-1 AD 2 G 0 = kcal mole-1 AM 2 GT 2 B05 - v gsartor ucleotidi e coenzimi
21 Trasporto di energia AT + 2 AD G = -7.3 kcal mole -1 (-30.6 kj mole -1 ) a p 7 G = -10 kcal mole -1 (-42 kj mole -1 ) a p B05 - v gsartor ucleotidi e coenzimi G Il valore di G dipende da: Forza ionica Concentrazione di Mg ++ Concentrazione di Ca ++ elle normali condiziono cellulari vale circa: -12 kcal mole -1 I nucleotidi sono contemporaneamente: tabili olubili Carichi Reattivi B05 - v gsartor ucleotidi e coenzimi
22 G Altre molecole fosforilate hanno G negativo G = -7.3 kcal mole -1 G = -3.3 kcal mole B05 - v gsartor ucleotidi e coenzimi erché il G dell AT vale kj mole -1? Repulsione tra cariche Maggiore in AT che in AD 2 2 B05 - v gsartor ucleotidi e coenzimi
23 erché il G dell AT vale 7.3 kcal mole -1? tabilizzazione per risonanza Del fosfato 3 -, + 3 -, + = B05 - v gsartor ucleotidi e coenzimi Trasporto di energia AT + 2 AM G 0 = -7.3 kcal mole -1 G 0 = -8 kcal mole B05 - v gsartor ucleotidi e coenzimi
24 AT e Mg ++ ella cellula l AT è spesso complessato con lo ione Mg ++ Mg++ 2 er formare un complesso ibridizzato B05 - v gsartor ucleotidi e coenzimi Altre molecole energetiche anno G 0 negativo nella reazione di idrolisi del gruppo fosfato. G = kcal mole -1 G = kcal mole -1 G = kcal mole -1 C Fosfoenolpiruvato Acetilfosfato Fosfocreatina B05 - v gsartor ucleotidi e coenzimi
25 Come si fa a ricaricare l AD per formare AT Attraverso la catalisi di un enzima (piruvato chinasi) E AD + 2 piruvato chinasi 2 + AT E iruvato B05 - v gsartor ucleotidi e coenzimi Trasporto di elettroni AD+ nicotinamide adenindinucleotide Acido nicotinico 2 icotinamide icotinamide Adenina Dinucleotide B05 - v gsartor ucleotidi e coenzimi
26 Trasporto di elettroni Come funziona il AD + 2 tato ossidato + AD e - ( - ) 2 tato ridotto AD 2 B05 - v gsartor ucleotidi e coenzimi Trasporto di elettroni Come funziona il AD + tato ossidato tato ridotto B05 - v gsartor ucleotidi e coenzimi
27 Trasporto di elettroni + R e - ( - ) R 2 AD + AD Trasporta DUE elettroni per molecola Reazione catalizzata dalle DEIDRGEAI B05 - v gsartor ucleotidi e coenzimi Gliceraldeide-3-fosfato deidrogenasi (EC ) G ' = +6.3 kj mol -1 3-fosfogliceraldeide GA C AD + AD AD+ B R AD+ R 2 B + 3 Complesso Enzima-substrato come Intermedio tioacetalico Intermedio aciltioestere AD R B C ,3-bifosfoglicerato 1,3BG GA 1 AD+ B AD+ R B + AD AD + R 1,3BG B05 - v gsartor ucleotidi e coenzimi
28 Gliceraldeide-3-fosfato deidrogenasi (EC ) AD + Cys carbossimetilata B05 - v gsartor ucleotidi e coenzimi Trasporto di elettroni AD + nicotinamide adenindinucleotide fosfato icotinamide Adenina Fosfato Dinucleotide B05 - v gsartor ucleotidi e coenzimi

29 Trasporto di elettroni FAD (flavinadenindinucleotide) (Forma ossidata) 3 C 3 C Base di hiff ridotta Ribosio in forma aperta 2 B05 - v gsartor ucleotidi e coenzimi Trasporto di elettroni FAD (flavinadenindinucleotide) (Forma ossidata) 3 C 3 C Base di hiff ridotta Ribosio in forma aperta 2 B05 - v gsartor ucleotidi e coenzimi
 C 3 C 3 Alcool Vanillico Aldeide Vanillica")
30 Trasporto di elettroni Come funziona il FAD 3 C C 3 R 3 C + R' + R C 3 R R R' FAD (ossidato) FAD z (ridotto) B05 - v gsartor ucleotidi e coenzimi Trasporto di elettroni Il FAD è legato alle proteine, (in questo caso la alcolvanillilico ossidasi) C 3 C 3 Alcool Vanillico Aldeide Vanillica (Vanillina) B05 - v gsartor ucleotidi e coenzimi
31 Coenzima A Gli acili e R gli acetili C 3 Vengono trasportati dal coenzima A () acido pantotenico AD β-mercaptoetilamina 3 C C 3 2 B05 - v gsartor ucleotidi e coenzimi Trasporto di acili Attraverso la formazione di un legame tioestere R C 3 C 3 2 Legame tioestere B05 - v gsartor ucleotidi e coenzimi

32 roteina acil- Complesso tra la proteina legante acil-, Coa e palmitoile B05 - v gsartor ucleotidi e coenzimi Vie metaboliche ono l insieme di reazioni chimiche che portano alla trasformazione di un substrato in un prodotto passando attraverso composti intermedi. Vie cataboliche: Vie di degradazione di un composto roducono energia chimica (AT e AD) e intermedi Vie anaboliche: Vie di sintesi di composti Utilizzano energia chimica (AT e AD) e intermedi. B05 - v gsartor ucleotidi e coenzimi 64 32
33 AD + i RTEIE LIACCARIDI LIIDI AT GLICLII GLUCEGEEI AD + i AT Aminoacidi Esosi; entosi Ac. Grassi; Glicerolo AD + i AT α-chetoacidi AD + i AT iruvato Acetil AD + i AT AD + i AT AT I FAE IDRLITICA AD + i II FAE IDATIVA Ciclo degli Acidi Tricarbossilici Fosforilazione ossidativa 2 AD + i AT III 3 2 C 2 B05 - v gsartor ucleotidi e coenzimi 65 Vie metaboliche ed intermedi Le vie metaboliche producono intermedi; Il destino dei composti intermedi può essere vario ; gni punto di biforcazione di una via metabolica permette una regolazione ; Le vie metaboliche più importanti sono comuni a tutti gli organismi ; rganismi diversi in ambienti diversi sono caratterizzati da vie metaboliche alternative ; Gli organismi vengono caratterizzati in funzione del modo con cui utilizzano carbonio, ossigeno ed energia. B05 - v gsartor ucleotidi e coenzimi 66 33


34 B05 - v gsartor ucleotidi e coenzimi 67 B05 - v gsartor ucleotidi e coenzimi 68 34



35 B05 - v gsartor ucleotidi e coenzimi 69 B05 - v gsartor ucleotidi e coenzimi 70 35
36 Regolazione delle vie metaboliche La velocità di una via metabolica può essere regolata sia dalla disponibilità del substrato (K m ) che dalla disponibilità dell enzima (V max ). È controllata dal passaggio più lento nella via metabolica Termodinamicamente molto favorito Catalizzato da un enzima estremamente regolato pesso in una biforcazione della via metabolica Le vie cataboliche (demolizione) ed anaboliche (sintesi) usano spesso gli stessi enzimi, ma hanno almeno un passaggio diverso. B05 - v gsartor ucleotidi e coenzimi 71 Regolazione delle vie metaboliche A B B B B B BB B C Reazione limitata dalla concentrazione di enzima. (lontana dall equilibrio) D E Reazione limitata dalla concentrazione di substrato. (all equilibrio o vicina all equilibrio) E E E E EE E E E EE F B05 - v gsartor ucleotidi e coenzimi 72 G 36
37 Glicogeno Metabolismo del Glicogeno C 2 C C Gluconeogensi Fermentazione lattica (Anaerobiosi) 2 Lattato D-Glucoso 2AD + 2i 2AD + Fruttoso-1,6-difosfato 2AT 2AD 2AD + 2 iruvato 3 C 2AD Fermentazione alcolica (Anaerobiosi) 2 C etanolo Glicolisi iruvato Acetil- Riboso-5-fosfato Ciclo dei pentosi 3-fosfogliceraldeide 2 AD AD + ssidazione aerobica AD + Ciclo di Krebs Fosforilazione ossidativa 6 C AD + i + + AT Glicogeno 3 C Glicolisi Via di Embden-Meyerhof (Warburg) Metabolismo del Glicogeno Gluconeogensi Fermentazione lattica (Anaerobiosi) 2 Lattato 2 iruvato 3 C C 2 D-Glucoso 2AD + 2i 2AD + Fruttoso-1,6-difosfato 2AT 2AD 2AD + 2AD Fermentazione alcolica (Anaerobiosi) 2 C etanolo Glicolisi iruvato Acetil- C Riboso-5-fosfato Ciclo dei pentosi 3-fosfogliceraldeide 2 AD AD + ssidazione aerobica AD + Ciclo di Krebs Fosforilazione ossidativa 6 C AD + i + + AT B05 - v gsartor ucleotidi e coenzimi 74 37

38 Glicolisi In tutte le cellule avviene la glicolisi Dieci reazioni, le stesse in tutte le cellule, ma con diversa cinetica Tre fasi: La prima fase converte il glucosio in fruttosio- 1,6-difosfato, La seconda fase scinde il fruttosio-1,6-difosfato in due triosi, La terza fase produce due molecole di piruvato. I prodotti sono: piruvato, AT e AD. Esistono tre diversi destini possibili del piruvato. B05 - v gsartor ucleotidi e coenzimi 75 Glicolisi Gli enzimi sono confinati nel citoplasma della cellula. Gli intermedi sono tutti fosforilati. on passano attraverso le membrane. Vengono riconosciuti dagli enzimi. ono convertiti in intermedi ad alto potenziale di trasferimento del fosfato che sono in grado di fosforilare l AD ad AT. B05 - v gsartor ucleotidi e coenzimi 76 38
39 Intermedi fosforilati otenziale di trasferimento del fosfato ( G ') kcal/mole Fosfoenolpiruvato 1,3-bifosfoglicerato AT Glucoso-6-fosfato 0 B05 - v gsartor ucleotidi e coenzimi 77 Fasi della glicolisi 1. Fosforilazione del substrato ed attivazione, 2. cissione dello zucchero a sei atomi di carbonio, 3. Recupero dell energia nella fosforilazione dell AD. B05 - v gsartor ucleotidi e coenzimi 78 39
40 Fosforilazione La prima fase della glicolisi consiste nella fosforilazione del glucoso per arrivare a fruttoso-1,6-difosfato attraverso il glucoso-6-fosfato che viene isomerizzato a fruttoso-6-fosfato. Vengono consumate due molecole di AT per ottenere lo zucchero difosfato. B05 - v gsartor ucleotidi e coenzimi 79 C 2 Glucoso Esochinasi Mg ++ AT AD Glucoso-6-fosfato 3 -- C 2 Fruttoso-6-fosfato 3 -- C 2 C 2 Fosfogluco isomerasi Fosfofrutto chinasi AT AD Fruttoso-1,6-bifosfato C 2 2 C C Fosfogliceratochinasi Mg ++ AT AD 3 -- Gliceraldeide- 3-fosfato deidrogenasi C 2 AD AD i Aldolasi C C 2 Fosfato triosoisomerasi fosfoglicerato 1,3-bifosfoglicerato 3-fosfogliceraldeide Diidrossiacetonfosfato Fosfogliceratomutasi Enolasi iruvato chinasi 3 -- Mg ++ Mg ++, K C 2 C 2 C 3 2 AD AT 2-fosfoglicerato Fosfoenolpiruvato iruvato B05 - v gsartor ucleotidi e coenzimi 80 40
41 Fase 1 Formazione del fruttoso- 1,6-difosfato Gli enzimi coinvolti sono due chinasi (la esochinasi e la fosfofruttochinasi) ed una isomerasi Risultato netto: C 2 Glucoso AT AD Mg ++ Glucoso-6-fosfato C Esochinasi (EC ) GLU G6 GLU + 2 AT FB + 2 AD Fosfoglucoisomerasi (EC ) C C 2 Fruttoso-6-fosfato F6 AT Fosfofruttochinasi (EC ) Mg ++ AD C C Fruttoso-1,6-bifosfato FB B05 - v gsartor ucleotidi e coenzimi 81 Esochinasi Fosforilazione del glucoso Consumo di AT Fosforilazione spontanea a causa dell idrolisi dell AT 2 2 AT AD Mg++ Glucoso Esochinasi (EC ) Glucoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi 82 41

42 Esochinasi inattiva Il legame del glucoso induce un cambiamento conformazionale B05 - v gsartor ucleotidi e coenzimi 83 Esochinasi attiva B05 - v gsartor ucleotidi e coenzimi 84 42
43 Esochinasi EC La fosforilazione confina il glucoso nella cellula La K m per il glucosio è 0.1 mm; nella cellula il glucosio è 4 mm La glucochinasi ha una K m glucoso = 10 mm, si attiva solo quando la cellula si arricchisce in glucosio La esochinasi è regolata allostericamente, viene inibita dal glucoso-6-fosfato ma non è il principale sito di regolazione della glicolisi B05 - v gsartor ucleotidi e coenzimi 85 Ruolo del Glucoso-6-fosfato Glucoso C 2 GLU Deposito di energia nel muscolo e nel fegato AT Glicogeno Mg ++ AD C C 2 Via dei pentosi fosfati Glucoso-6-fosfato G Glucoso-1-fosfato intesi di AD e di zuccheri a 3, 5, 6 e 7 atomi di carbonio C C 2 Fruttoso-6-fosfato Glucuronato Glucosamina-6-fosfato intesi di carboidrati F6 B05 - v gsartor ucleotidi e coenzimi 86 43
44 C 2 Glucoso AT GLU Mg ++ Esochinasi (EC ) AD C Glucoso-6-fosfato G6 Fosfoglucoisomerasi (EC ) C C 2 Fruttoso-6-fosfato AT F6 Mg ++ Fosfofruttochinasi (EC ) AD C C Fruttoso-1,6-bifosfato F6 B05 - v gsartor ucleotidi e coenzimi 87 C 2 Glucoso AT GLU Mg ++ Esochinasi (EC ) AD C Glucoso-6-fosfato G6 Fosfoglucoisomerasi (EC ) C C 2 Fruttoso-6-fosfato AT F6 Mg ++ Fosfofruttochinasi (EC ) AD C C Fruttoso-1,6-bifosfato FB B05 - v gsartor ucleotidi e coenzimi 88 44
45 Fosfofruttochinasi EC È la reazione che controlla la glicolisi È la seconda reazione di fosforilazione Valore di G grande e negativo, La FK è altamente regolata AT inibisce, AM elimina l inibizione Il citrato è un inibitore allosterico Il fruttoso-2,6-bifosfato è un attivatore allosterico L attività della FK aumenta quando lo stato energetico della cellula è basso. L attività della FK diminuisce quando lo stato energetico della cellula è alto. pinge la reazione verso la glicolisi e non verso il ciclo dei pentosi B05 - v gsartor ucleotidi e coenzimi 89 Glucoso C 2 GLU Deposito di energia nel muscolo e nel fegato AT Glicogeno Mg ++ Via dei pentosi fosfati AD C G6 Glucoso-6-fosfato C Glucoso-1-fosfato intesi di AD e di zuccheri a 5, 6 e 7 atomi di carbonio C C 2 Fruttoso-6-fosfato Glucuronato Glucosamina-6-fosfato F6 AT AT Citrato Fosfofruttochinasi (EC ) Mg ++ AM AD Fruttoso-2,6-bifosfato C C Fruttoso-1,6-bifosfato FB intesi di carboidrati B05 - v gsartor ucleotidi e coenzimi 90 45

46 Fosfofruttochinasi EC G ' = kj mol -1 G eritrociti = kj mol -1 Mg ++ + AT + AD Fruttoso-6-fosfato F6 Fruttoso-1,6-bifosfato FB B05 - v gsartor ucleotidi e coenzimi 91 Fosfofruttochinasi EC Dimero di un tetramero AD Mg ++ FB Mg ++ AD B05 - v gsartor ucleotidi e coenzimi 92 46

47 Fosfofruttochinasi EC Dimero di un tetramero F6 B05 - v gsartor ucleotidi e coenzimi 93 Regolazione FK B05 - v gsartor ucleotidi e coenzimi 94 47

48 Enzima tandem La regolazione della FK coinvolge un altra molecola fosforilata: il fruttoso-2,6-bifosfato, che funziona da attivatore allosterico della FK. La sintesi di questa molecola è regolata da una proteina che, su singolo polipetide, svolge due attività enzimatiche diverse a secondo se è o non è fosforilato: l enzima tandem. L enzima tandem è finemente regolato. 2 C 3 -- AT C 2 2 C cam C 2 6-Fosfofrutto-2-chinasi (EC ) + Fosforilasi Chinasi A Attiva + Enzima defosforilato Fosforilasi Chinasi A Inattiva + AD 2 C C 2 i - Enzima fosforilato Fruttoso-2,6-difosfatasi (EC ) B05 - v gsartor ucleotidi e coenzimi 95 Enzima tandem B05 - v gsartor ucleotidi e coenzimi 96 48
49 Fase 2 cissione del fruttoso- 1,6-difosfato La scissione del fruttoso-1,6-difosfato attraverso l aldolasi porta alla formazione di due triosi, diidrossiacetonfosfato e 3-fosfogligeraldeide. I due triosi sono tra loro in equilibrio attraverso la trioso fosfato isomerasi C C C C C Trioso fosfato isomerasi C (EC ) 3-fosfogliceraldeide Diidrossiacetonfosfato DA Fruttoso-1,6-bifosfato F6 Aldolasi (EC ) GA B05 - v gsartor ucleotidi e coenzimi 97 Aldolasi: meccanismo generale C C C C R C 2 R C 2 - R C 2 2 R - C 2 - R C 3 R' R' + rodotto II R' rodotto I Enzima Zn++ C Enzima Zn++ C C B05 - v gsartor ucleotidi e coenzimi 98 49

50 Aldolasi EC C C C C R - 2 C 2 R C 2 R C 2 - R C R C 3 R' R' rodotto II + R' rodotto I Enzima Zn++ C Enzima Zn++ C C B05 - v gsartor ucleotidi e coenzimi 99 Aldolasi EC FB B05 - v gsartor ucleotidi e coenzimi

51 Aldolasi EC Lys Tyr FB B05 - v gsartor ucleotidi e coenzimi 101 Trioso-fosfato isomerasi EC Converte DA in GA Il meccanismo coinvolge la formazione di enendiolo Il sito attivo contiene un Glu che agisce come base B05 - v gsartor ucleotidi e coenzimi
52 Trioso-fosfato isomerasi EC Catalizza l equilibrio: C Trioso fosfato isom erasi C (EC ) Diidrossiacetonfosfato 3-fosfogliceraldeide DA G ' = kj mol -1 L equilibrio è spostato verso sinistra ( 96% DA, 4% GA), nel procedere della glicolisi viene consumata solo GA e l equilibrio si sposta verso destra. GA B05 - v gsartor ucleotidi e coenzimi 103 Trioso-fosfato isomerasi EC Glu :B+ Glu :B Diidrossiacetonfosfato DA Glu :B+ 3-fosfogliceraldeide GA 3 -- B05 - v gsartor ucleotidi e coenzimi


53 Trioso-fosfato isomerasi EC B05 - v gsartor ucleotidi e coenzimi 105 Trioso-fosfato isomerasi EC Glu mologo di DA Lys B05 - v gsartor ucleotidi e coenzimi

54 Trioso-fosfato isomerasi EC Glu mologo di DA B05 - v gsartor ucleotidi e coenzimi 107 Fase 3 Recupero dell energia La formazione di GA permette il recupero dell energia attraverso il suo metabolismo con formazione di una serie di intermedi forsforilati: 1,3-bifosfoglicerato, 3-fosfoglicerato, 2-fosfoglicerato, fosfoenolpiruvato ed infine piruvato. Il destino del piruvato dipende dalla presenza di ossigeno e può essere diverso in cellule diverse (lievito, muscolo ) B05 - v gsartor ucleotidi e coenzimi
55 Fase 3 Recupero dell energia La 3-fosfogligeraldeide prodotta viene ossidata e fosforilata 1,3- bifosfoglicerato da una deidrogenasi, con produzione di AD il 1,3-bifosfoglicerato viene utilizzato per fosforilare l AD ad opera di una fosfogliceratochinasi e si forma 3-fosfoglicerato, che viene isomerizzato a 2- fosfoglicerato ad opera di una mutasi, il 2-fosfoglicerato perde una molecola d acqua ad opera di una enolasi e si forma il fosfoenolpiruvato che viene trasformato in piruvato ad opera della piruvato chinasi con formazione di AT. 3-fosfogliceraldeide GA 1,3-bifosfoglicerato 1,3BG -- 3 Gliceraldeide-3-fosfato deidrogenasi (EC ) C Fosfoglicerato chinasi (EC ) 3-fosfoglicerato 3G AD C Fosfoglicerato mutasi (EC ) 2-fosfoglicerato 2G Fosfoenolpiruvato E iruvato chinasi (EC ) iruvato C Enolasi (EC ) 3 -- C 2 3 C Mg ++ C AD + AD AT 2 AD AT C 2 Enolpiruvato B05 - v gsartor ucleotidi e coenzimi 109 Gliceraldeide-3-fosfato deidrogenasi EC GA è ossidata a 1,3BG L energia ottenuta dalla conversione di un aldeide ad acido carbossilico è usata per la fosforilazione a 1,3BG e per la riduzione del AD + a AD B05 - v gsartor ucleotidi e coenzimi

56 Gliceraldeide-3-fosfato deidrogenasi EC G ' = +6.3 kj mol -1 AD+ Intermedio tioacetalico 3-fosfogliceraldeide GA C AD + AD AD+ B R R 2 B + 3 Complesso Enzima-substrato come Intermedio aciltioestere AD R B C ,3-bifosfoglicerato 1,3BG GA 1 AD+ B 1,3BG B05 - v gsartor ucleotidi e coenzimi 111 R AD+ R 4 B + AD + AD Gliceraldeide-3-fosfato deidrogenasi EC B05 - v gsartor ucleotidi e coenzimi
 AD AT 3-fosfoglicerato 3G C 2 3 -- B05 - v 1.8.1 gsartor 2001-2018 ucleotidi e coenzimi 114 57")
57 Gliceraldeide-3-fosfato deidrogenasi EC AD + Cys carbossimetilata B05 - v gsartor ucleotidi e coenzimi 113 Fosfoglicerato chinasi EC ,3-bifosfoglicerato -- 3 AD 1,3BG C Fosfoglicerato chinasi (EC ) AD AT 3-fosfoglicerato 3G C B05 - v gsartor ucleotidi e coenzimi
58 Fosfoglicerato chinasi EC Il gruppo fosfato passa da C-3 a C-2 postamento di fosfato per la formazione di E i forma un intermedio fosfoistidina È stato dimostrato che del 2,3BG è richiesto per la fosforilazione di is. B05 - v gsartor ucleotidi e coenzimi 115 Fosfoglicerato chinasi EC fosfoglicerato 3G C Fosfoglicerato mutasi (EC ) G ' = +4.4 kj mol -1 2-fosfoglicerato 2G 3 -- C 2 B05 - v gsartor ucleotidi e coenzimi


59 Fosfoglicerato mutasi EC B : B + : 3-fosfoglicerato 3BG intermedio 2,3-bifosfoglicerato + 2-fosfoglicerato 2BG B05 - v gsartor ucleotidi e coenzimi fosfoglicerato 2-fosfoglicerato B05 - v gsartor ucleotidi e coenzimi


60 Fosfoglicerato mutasi EC is is Mn ++ 3G is Mn ++ B05 - v gsartor ucleotidi e coenzimi 119 Fosfoglicerato mutasi EC is is Mn ++ 3G is B05 - v gsartor ucleotidi e coenzimi
61 Enolasi EC Da 2G a E Il G globale è 1.8 kj/mol Il contenuto in energia di 2G e E è simile. L enolasi riarrangia la molecola in modo tale che possa fornire più energia nell idrolisi. B05 - v gsartor ucleotidi e coenzimi 121 Enolasi EC fosfoglicerato 2G 3 -- C 2 G ' = +1.8 kj mol -1 Mg ++ Fosfoenolpiruvato E C 2 B05 - v gsartor ucleotidi e coenzimi


62 Enolasi EC E Mg ++ B05 - v gsartor ucleotidi e coenzimi 123 Enolasi EC Glu Mg ++ B05 - v gsartor ucleotidi e coenzimi 124 E 62
63 iruvato Chinasi EC Da E a piruvato viene prodotto AT Valore di G grande e negativo. unto di regolazione Attivata allostericamente da AM, F1,6B Inibita allostericamente da AT e acetil- Tautomeria chetoenolica del piruvato. B05 - v gsartor ucleotidi e coenzimi 125 iruvato Chinasi EC G ' = kj mol -1 AD AT 2 C 3 -- Fosfoenolpiruvato iruvato chinasi (EC ) 2 C Forma enolica 3 C Forma chetonica E iruvato B05 - v gsartor ucleotidi e coenzimi
64 iruvato Chinasi EC M+ Mg++ Mg++ B 2. M+ Mg++ Mg++ 2 B. B05 - v gsartor ucleotidi e coenzimi 127 iruvato Chinasi EC K + Mg++ C 2 Mg++ Adenosina E AD AT Mg++ + C 2 Enolpiruvato C 3 iruvato B05 - v gsartor ucleotidi e coenzimi

65 iruvato Chinasi EC B05 - v gsartor ucleotidi e coenzimi 129 C 2 Glucoso Esochinasi Mg ++ AT AD Glucoso-6-fosfato 3 -- C 2 Fruttoso-6-fosfato 3 -- C 2 C 2 Fosfogluco isomerasi Fosfofrutto chinasi AT AD Fruttoso-1,6-bifosfato C 2 2 C C Fosfogliceratochinasi Mg ++ AT AD 3 -- Gliceraldeide- 3-fosfato deidrogenasi C 2 AD AD i Aldolasi C C 2 Fosfato triosoisomerasi fosfoglicerato 1,3-bifosfoglicerato 3-fosfogliceraldeide Diidrossiacetonfosfato Fosfogliceratomutasi Enolasi iruvato chinasi 3 -- Mg ++ Mg ++, K C 2 C 2 C 3 2 AD AT 2-fosfoglicerato Fosfoenolpiruvato iruvato B05 - v gsartor ucleotidi e coenzimi
66 Energia Energia Libera (kj mole -1 ) Esochinasi Fosfoglucosoisomerasi Fosfofruttochinasi Aldolasi Trioso-fosfato isomerasi GA-deidrogenasi GA Chinasi GA Mutasi Enolasi iruvato Chinasi assi della Glicolisi tato tandard ( G ) In eritrociti ( G) B05 - v gsartor ucleotidi e coenzimi 131 C 2 Glucoso 3 -- C 2 Glucoso-6-fosfato 3 -- C 2 C 2 Fruttoso-6-fosfato Mg ++ AT IT DI Esochinasi REGLAZIE AD Fosfoglucoisomerasi AT 3 -- C 2 2 C 3 -- Fruttoso-1,6-bifosfato IT DI Fosfofruttochinasi REGLAZIE AD 2 C 3 -- Diidrossiacetonfosfato Aldolasi Gliceraldeide- 1,3-bifosfoglicerato 3-fosfato 3 -- C 2 deidrogenasi 3 -- C 2 AD + + i fosfogliceraldeide AD AD Fosfogliceratochinasi Mg ++ C 3 iruvato IT DI REGLAZIE AT iruvato chinasi Mg ++, K + AD C Fosfoenolpiruvato AT 3 -- C 2 Fosfogliceratomutasi C 2 Enolasi 3 -- Mg ++ 3-fosfoglicerato 2-fosfoglicerato B05 - v gsartor ucleotidi e coenzimi

67 ssigeno Aerobiosi e anaerobiosi Destino del piruvato In assenza di ossigeno Riduzione a lattato rima riduzione a lattato poi decarbossilazione ad acetato rima decarbossilazione ad aldeide poi riduzione ad etanolo In presenza di ossigeno Decarbossilazione, Ciclo di Krebs, Respirazione cellulare Ripristino di AD + per continuare la glicolisi B05 - v gsartor ucleotidi e coenzimi
68 Destino del piruvato In assenza di ossigeno el muscolo Ridotto a Lattato el lievito Decarbossilato ad aldeide ridotta quindi ad etanolo Ripristino di AD + per continuare la glicolisi B05 - v gsartor ucleotidi e coenzimi 135 chema generale del metabolismo dei glucidi Glicogeno Metabolismo del Glicogeno C 2 C C Gluconeogensi Fermentazione lattica (Anaerobiosi) 2 Lattato D-Glucoso 2AD + 2i 2AD + Fruttoso-1,6-difosfato 2AT 2AD 2AD + 2 iruvato 3 C 2AD Fermentazione alcolica (Anaerobiosi) 2 C etanolo Glicolisi iruvato Acetil- Riboso-5-fosfato Ciclo dei pentosi 3-fosfogliceraldeide 2 AD AD + ssidazione aerobica AD + Ciclo di Krebs Fosforilazione ossidativa 6 C AD + i + + AT B05 - v gsartor ucleotidi e coenzimi
69 Anaerobiosi 3-fosfogliceraldeide Diidrossiacetonfosfato 3 -- C 2 2 C 3 -- Gliceraldeide- 3-fosfato deidrogenasi i 1,3-bifosfoglicerato 3 -- C 2 el muscolo AD AD + Lattato C 3 Fosfogliceratochinasi 3 -- AD Mg ++ 3-fosfoglicerato 3 -- C 2 AT Lattico deidrogenasi Fosfogliceratomutasi iruvato C 3 iruvato chinasi Mg ++, K + Fosfoenolpiruvato 2-fosfoglicerato C 2 Enolasi 3 -- Mg ++ C AT AD B05 - v gsartor ucleotidi e coenzimi 137 Lattico deidrogenasi EC R 2 AD 2 3 C is195 2 R + Arg171 AD C B05 - v gsartor ucleotidi e coenzimi

70 Lattico deidrogenasi EC mologo del piruvato Arg 171 is 195 AD + B05 - v gsartor ucleotidi e coenzimi 139 Anaerobiosi 3-fosfogliceraldeide Diidrossiacetonfosfato 3 -- C 2 2 C 3 -- Gliceraldeide- 3-fosfato deidrogenasi i 1,3-bifosfoglicerato 3 -- C 2 el lievito Etanolo C 3 C 2 AD + AD 3 -- AD Alcol deidrogenasi Fosfogliceratochinasi Mg ++ Acetaldeide C 3 3-fosfoglicerato 3 -- C 2 AT C 2 Fosfogliceratomutasi iruvato decarbossilasi iruvato C 3 iruvato chinasi Mg ++, K + Fosfoenolpiruvato 2-fosfoglicerato C 2 Enolasi 3 -- Mg ++ C AT AD B05 - v gsartor ucleotidi e coenzimi
71 iruvato decarbossilasi EC Catalizza la decarbossilazione del piruvato. Usa la tiamina pirofosfato come coenzima 2 3 C ACID + C 3 ACID B05 - v gsartor ucleotidi e coenzimi 141 iruvato decarbossilasi EC Acetaldeide C iruvato R + C 3 C C 3 R' + R + C 3 δ- R' T R + C 3 C 3 R' + C 2 C 3 R + C 3 R' C 3 R C 3 R' C 3 C R + C 3 R' B05 - v gsartor ucleotidi e coenzimi

72 iruvato decarbossilasi EC Mg ++ T B05 - v gsartor ucleotidi e coenzimi 143 iruvato decarbossilasi EC Catalizza la reazione di ossidoriduzione: C 3 + AD C 3 + AD+ Acetaldeide R Alcol etilico 2 + C 3 Zn++ C 3 AD+ B05 - v gsartor ucleotidi e coenzimi


73 iruvato decarbossilasi EC AD + Zn ++ B05 - v gsartor ucleotidi e coenzimi 145 Alcool deidrogenasi EC AD + B05 - v gsartor ucleotidi e coenzimi



74 Alcool deidrogenasi EC AD + AD + Cys Cys Trifluoroetanolo Zn ++ is Trifluoroetanolo Zn ++ is B05 - v gsartor ucleotidi e coenzimi 147 Alcool deidrogenasi EC AD + mologo di AD + Cys Cys Trifluoroetanolo Zn ++ is Etanolo Zn ++ is B05 - v gsartor ucleotidi e coenzimi
75 Energetica della Glicolisi I valori di G sono variabili (positivi e negativi) G nelle cellule ha valori vicini a zero olo tre reazioni su dieci hanno valori di G negativo e grande. Le reazioni i cui valori di G sono grandi e negativi sono punti di regolazione. B05 - v gsartor ucleotidi e coenzimi 149 Energia Energia Libera (kj mole -1 ) Esochinasi Fosfoglucosoisomerasi Fosfofruttochinasi Aldolasi Trioso-fosfato isomerasi GA-deidrogenasi GA Chinasi GA Mutasi Enolasi iruvato Chinasi assi della Glicolisi tato tandard ( G ) In eritrociti ( G) B05 - v gsartor ucleotidi e coenzimi
76 Energia Energia Libera (kj mole -1 ) tato tandard ( G ) In eritrociti ( G) assi della Glicolisi B05 - v gsartor ucleotidi e coenzimi 151 Energia dalla glicolisi G = kj mol -1 C C G = kj mol -1 La scissione di glucoso a piruvato utilizza solo il 1.7% del contenuto energetico del glucoso. B05 - v gsartor ucleotidi e coenzimi
77 rganizzazione Gli enzimi glicolitici possono formare complessi multiezimatici; ella purificazione di proteine da cellule le interazioni non covalenti tra proteine viene persa; È stato suggerito che gli enzimi glicolitici si assemblino in un complesso multienzimatico dove i substrati sono canalizzati da un enzima all altro senza passare in soluzione. B05 - v gsartor ucleotidi e coenzimi 153 Gluconeogenesi intesi di glucoso da piruvato La gluconeogenesi avviene principalmente nel fegato. La sintesi di glucoso da piruvato utilizza alcuni enzimi della glicolisi. Tre reazioni glicolitiche hanno un valore di G talmente negativo e grande che le reazioni sono irreversibili: Esochinasi Fosfofruttochinasi iruvato chinasi Questi passaggi sono by-passati nella gluconeogenesi. B05 - v gsartor ucleotidi e coenzimi
78 Bypass della piruvato chinasi La piruvato chinasi della glicolisi catalizza la reazione: 2 C 3 -- M g ++, K + 3 C AD AT Fosfoenolpiruvato iruvato L idrolisi del E ha un valore di G (negativo) maggiore dell AT. Il G ottenibile dall idrolisi di un legame fosfato è insufficiente per sintetizzare il E. È richiesta l idrolisi di due legami fosfoanidridici (da due T diversi, AT e GT o AT o i). B05 - v gsartor ucleotidi e coenzimi 155 Bypass della piruvato chinasi er bypassare la piruvato chinasi occorrono due reazioni: Carbossilazione del piruvato, catalizzata da piruvato carbossilasi (EC ): Fosforilazione e decarbossilazione (spontanea) dell ossalacetato a E catalizzata dalla E carbossichinasi (EC ): 3 C iruvato C 3 - AT AD ssalacetato GT GD C ssalacetato C 2 Fosfoenolpiruvato B05 - v gsartor ucleotidi e coenzimi
79 iruvato carbossilasi EC È un enzima a biotina 3 C C 3 La biotina si lega ad una lisina nel sito attivo dell enzima formando un lungo braccio flessibile ad Lisina una estremità del quale vi è il sito di carbossilazione. * ito di carbossilazione 3 C C 3 B05 - v gsartor ucleotidi e coenzimi 157 Legame ε-peptidico * iruvato carbossilasi EC Il lungo braccio flessibile permette il movimento della biotina tra il sito di carbossilazione e il sito di decarbossilazione e formazione del ossalacetato. La carbossilazione avviene ad opera di carbossifosfato che si forma nel sito di carbossilazione per reazione di AT e bicarbonato. AT AD C 3 - Carbossifosfato Biotina 3 C C 3 Carbossibiotina 3 C C 3 C 3 3 C B05 - v gsartor ucleotidi e coenzimi
80 iruvato carbossilasi EC La decarbossilazione della biotina e la formazione di ossalacetato avviene nel secondo sito della piruvato carbossilasi dove si lega il piruvato per formare ossalacetato. 3 C C 3 3 C 3 C 3 C C 3 C 3 C 3 3 C B05 - v gsartor ucleotidi e coenzimi 159 iruvato carbossilasi EC Regola il destino del piruvato Glucoso-6- Glucoso Glicolisi iruvato Aminoacidi Gluconeogenesi + Acetil- Aminoacidi ssalacetato Ciclo di Krebs Citrato B05 - v gsartor ucleotidi e coenzimi

 EC4.1.1.38 (i) EC4.1.1.49 (AT) nei batteri C 2 C 2 GT GD 2 C 3 -- ssalacetato Enolpiruvato Fosfoenolpiruvato Mg ++ Mn ++ Mn ++ Mg ++ B05 - v 1.")
81 iruvato carbossilasi EC Quando la gluconeogenesi è attiva nel fegato l ossalacetato va a formare glucoso. La diminuzione di ossalacetato causa la riduzione di Acetil che entra nel ciclo di Krebs. L aumento di Acetil attiva, allostericamente, la piruvato carbossilasi per formare ossalacetato. La concentrazione di ossalacetato limita il ciclo di Krebs. B05 - v gsartor ucleotidi e coenzimi 161 iruvato carbossichinasi EC (GT) EC (i) EC (AT) nei batteri C 2 C 2 GT GD 2 C 3 -- ssalacetato Enolpiruvato Fosfoenolpiruvato Mg ++ Mn ++ Mn ++ Mg ++ B05 - v gsartor ucleotidi e coenzimi
82 Globalmente Bypass della piruvato chinasi C 3 - AT AD GT C 2 GD 3 C iruvato ssalacetato C Fosfoenolpiruvato C 3 - AT AD GT GD C 2 3 C iruvato C Fosfoenolpiruvato B05 - v gsartor ucleotidi e coenzimi 163 Gluconeogenesi C 3 - AT AD ssalacetato GT GD C 2 Fosfoenolpiruvato 2-fosfoglicerato 3 C iruvato iruvato carbossilasi EC Diidrossiacetonfosfato 2 C fosfogliceraldeide 3 -- Trioso fosfato isomerasi E carbossichinasi EC (GT) EC (i) EC (AT) Aldolasi C 2 C AD + + i AD Gliceraldeide- 3-fosfato deidrogenasi 3 -- Enolasi Fosfogliceratomutasi 3-fosfoglicerato AD Fosfogliceratochinasi AT C 2 C ,3-bifosfoglicerato 3 -- C Fruttoso-1,6-bifosfato C 2 C B05 - v gsartor ucleotidi e coenzimi

83 Fruttoso-1,6-bifosfatasi EC Catalizza la reazione inversa della fosfofruttochinasi: i 3 -- C 2 2 C 3 -- C 2 2 C 3 -- Fruttoso-1,6-bifosfato Fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi 165 Fruttoso-1,6-bifosfatasi EC Zn ++ (Mg ++ ) i F6 F6 B05 - v gsartor ucleotidi e coenzimi

84 Glucoso-6-fosfatasi EC Catalizza la reazione inversa della esochinasi attraverso una fosfoistidina. R R R + + R Extracellulare È un enzima della membrane del reticolo endoplasmatico con funzione di traslocasi per la secrezione extracellulare del glucoso. È ancorato alla membrana da nove eliche transmembrana e secerne nel lume del reticolo. B05 - v gsartor ucleotidi e coenzimi 167 Glucoso-6-fosfatasi EC is i B05 - v gsartor ucleotidi e coenzimi
85 Glicolisi e Gluconeogenesi C 2 Glucoso AT IT DI Mg ++ Esochinasi 3 -- C 2 REGLAZIE AD Glucoso-6-fosfato Fosfoglucoisomerasi 3 -- C 2 C 2 Fruttoso-6-fosfato AT IT DI 3 -- C Fosfofruttochinasi 2 2 C 3 -- REGLAZIE Fruttoso-1,6-bifosfato AD C 3 iruvato AT 2 C 3 -- Diidrossiacetonfosfato IT DI REGLAZIE iruvato chinasi Mg ++, K + AD C Fosfoenolpiruvato Aldolasi C 2 3-fosfogliceraldeide Enolasi Mg ++ AD + + i C fosfoglicerato AD 3 -- Fosfogliceratochinasi Fosfogliceratomutasi C Mg ++ 3-fosfoglicerato AD AT Fruttoso-6-fosfato Glucoso-6-fosfato Glucoso 3 -- Fruttoso-1,6-bifosfato C 2 C 2 2 C C 2 2 C C 3 -- Fruttoso-1,6-bifosfatasi EC i i Fosfoglucoisomerasi Glucoso-6-fosfatasi EC B05 - v gsartor ucleotidi e coenzimi 169 C 2 Glicolisi e Gluconeogenesi 3 C iruvato C 3 - AT 2 C 3 -- AD iruvato carbossilasi EC Diidrossiacetonfosfato Fruttoso-1,6-bifosfato Fruttoso-6-fosfato 3 -- C 2 C 2 ssalacetato GT C 3 -- GD C 2 C 2 E carbossichinasi EC (GT) EC (i) EC (AT) 3-fosfogliceraldeide Aldolasi 2 C 3 -- C Gliceraldeide- 1,3-bifosfoglicerato 3-fosfato deidrogenasi 3 -- C 2 C 3 - ssalacetato C 2 Fosfoenolpiruvato 2-fosfoglicerato AD GD AT GT iruvato C E carbossichinasi Enolasi carbossilasi EC (GT) EC C 2 C 2 iruvato EC (i) EC (AT) Fosfogliceratomutasi 3-fosfoglicerato AD Fosfogliceratochinasi AT C Diidrossiacetonfosfato 3-fosfogliceraldeide AD + + i AD C 3 -- Gliceraldeide C 2 3-fosfato deidrogenasi 3 -- C 2 Aldolasi 1,3-bifosfoglicerato Glucoso AT Mg ++ Esochinasi 3 -- C 2 AD Glucoso-6-fosfato Fosfoglucoisomerasi 3 -- C 2 C 2 Fruttoso-6-fosfato AT 3 -- C Fosfofruttochinasi 2 2 C 3 -- Fruttoso-1,6-bifosfato AD Aldolasi Gliceraldeide- 1,3-bifosfoglicerato 3-fosfato 2 C C 2 deidrogenasi 3 -- C 2 Diidrossiacetonfosfato AD + + i fosfogliceraldeide AD AD Gliceraldeide- 3-fosfato 3 -- deidrogenasi i Fosfoenolpiruvato AD + + i AD Enolasi 3-fosfoglicerato C 2 C 2 Fosfogliceratomutasi AD Fosfogliceratochinasi AT Fruttoso-1,6-bifosfatasi EC fosfoglicerato ,3-bifosfoglicerato 3 -- C Fosfogliceratochinasi Mg ++ 2 C 3 -- Fosfoglucoisomerasi C 3 iruvato AT 3 -- C 2 iruvato Fosfogliceratomutasi chinasi C Mg ++, K + 2 C 2 Enolasi Mg ++ 3-fosfoglicerato AT AD Fosfoenolpiruvato 2-fosfoglicerato Glucoso-6-fosfato Glucoso C 2 i Glucoso-6-fosfatasi EC B05 - v gsartor ucleotidi e coenzimi
86 C 2 Glicolisi e Gluconeogenesi 3 C iruvato C 3 - AT 2 C 3 -- AD iruvato carbossilasi EC Diidrossiacetonfosfato Fruttoso-1,6-bifosfato Fruttoso-6-fosfato 3 -- C 2 C 2 ssalacetato Aldolasi GT C 3 -- GD C 2 C 2 E carbossichinasi EC (GT) EC (i) EC (AT) 3-fosfogliceraldeide 2 C 3 -- C Glucoso AT Mg ++ Esochinasi 3 -- C 2 AD Glucoso-6-fosfato Fosfoglucoisomerasi 3 -- C 2 C 2 Fruttoso-6-fosfato AT 3 -- C Fosfofruttochinasi 2 2 C 3 -- Fruttoso-1,6-bifosfato AD Aldolasi Gliceraldeide- 1,3-bifosfoglicerato 3-fosfato 2 C C 2 deidrogenasi 3 -- C 2 Diidrossiacetonfosfato AD + + i fosfogliceraldeide AD AD Gliceraldeide- 3-fosfato 3 -- deidrogenasi i Fosfoenolpiruvato AD + + i AD Enolasi C 2 C 2 Fosfogliceratomutasi 3-fosfoglicerato AD Fosfogliceratochinasi AT Fruttoso-1,6-bifosfatasi EC fosfoglicerato ,3-bifosfoglicerato 3 -- C Fosfogliceratochinasi Mg ++ 2 C 3 -- Fosfoglucoisomerasi C 3 iruvato chinasi Mg ++, K + AT AD AT 3 -- C 2 Fosfogliceratomutasi C 2 C 2 Enolasi Mg ++ 3-fosfoglicerato Glucoso-6-fosfato Glucoso C 2 i Glucoso-6-fosfatasi EC iruvato Fosfoenolpiruvato 2-fosfoglicerato B05 - v gsartor ucleotidi e coenzimi 171 Glicolisi e gluconeogenesi La glicolisi e la gluconeogenesi sono vie metaboliche spontanee. e fossero attive simultaneamente nella cellula si sarebbe in presenza di un ciclo futile con consumo di energia. Glicolisi (da glucoso a piruvato): C AD + + 2AD + 2 i Gluconeogenesi (da piruvato a glucoso): 2C AD AT + 2GT Glicolisi + Gluconeogenesi: 2C AD AT C AD + + 4AD + 2GD + 6 i 2 AT + 2 GT 2 AD + 2 GD + 4 i B05 - v gsartor ucleotidi e coenzimi
87 Controllo Locale er prevenire la perdita di energia nel ciclo futile la Glicolisi e la Gluoconeogenesi sono reciprocamente regolate. Controllo locale: Reciproco controllo allosterico ad opera dei nucleotidi adenilici: La fosfofruttochinasi (Glicolisi) è inibita da AT e stimolata da AM. La fruttoso-1,6-bifosfatasi (Gluconeogenesi) è inibita da AM. Quando la concentrazione di AT è alta (concentrazione di AM bassa) il glucoso è degradato per produrre AT. In queste condizioni la cellula accumula glicogeno. Quando la concentrazione di AT è bassa (concentrazione di AM alta) la cellula spende energia per sintetizzare glucoso. B05 - v gsartor ucleotidi e coenzimi 173 Controllo Globale Controllo globale: egli epatociti vi è l effetto reciproco sulle due vie dell AM ciclico, la cui cascata è attivata dall ormone GLUCAGE quando il glucoso ematico è basso. La rotein Chinasi A (rotein Chinasi cam Dipendente) provoca la fosforilazione di enzimi e proteine regolatrici il cui risultato è: inibizione della glicolisi stimolazione della gluconeogenesi, Ciò porta alla disponibilità di glucosio nel sangue. B05 - v gsartor ucleotidi e coenzimi
88 Controllo Globale Controllo globale: Gli enzimi che sono FFRILATI dalla roteina Chinasi A sono: iruvato Chinasi: enzima glicolitico che è inibito quando fosforilato. CREB (cam response element binding protein): che attiva attraverso sistemi di trascrizione il gene della E Carbossichinasi, con conseguente aumento della gluconeogenesi. L enzima tandem: che regola la formazione e la degradazione del regolatore allosterico fruttoso- 2,6-bifosfato. B05 - v gsartor ucleotidi e coenzimi 175 Controllo Globale Controllo globale: L enzima tandem: che regola la formazione e la degradazione del regolatore allosterico fruttoso- 2,6-bifosfato. Il fruttoso-2,6-bifosfato attiva la Fosfofruttochinasi anche in presenza di alto AT (che la inibisce). L attività in presenza di fruttoso-2,6-bifosfato è simile all attività con AT basso. Il controllo attraverso fruttoso-2,6-bifosfato (la cui concentrazione viene controllata da segnali esterni: ormoni) è gerarchicamente più importante del controllo locale da AT. Il fruttoso-2,6-bifosfato inibisce l enzima della gluconeogenesi fruttoso-1,6-bifosfatatasi. B05 - v gsartor ucleotidi e coenzimi
 che catalizza: Fruttoso-6-fosfato + AT fruttoso-2,6-bifosfato + AD Fruttoso-bifosfatasi (FBasi2) (EC3.1.3.46)che catalizza: Fruttoso-2,6-bifosfato + 2 fruttoso-6-fosfato + i F6 B05 - v 1.8.")
89 modimero Due domini catalitici: Enzima tandem Dominio di Fosforilazione (regolatorio) Fosfofruttochinasi (FK2) (EC ) che catalizza: Fruttoso-6-fosfato + AT fruttoso-2,6-bifosfato + AD Fruttoso-bifosfatasi (FBasi2) (EC )che catalizza: Fruttoso-2,6-bifosfato + 2 fruttoso-6-fosfato + i F6 B05 - v gsartor ucleotidi e coenzimi 177 Enzima tandem L enzima tandem è regolato dalla cascata del cam che a sua volta è controllato da ormoni AT 6-Fosfofrutto-2-chinasi (EC ) Enzima defosforilato + AD C C 2 Fosforilasi Chinasi A Attiva + cam Fosforilasi Chinasi A Inattiva C 3 -- C 2 i - Enzima fosforilato Fruttoso-2,6-difosfatasi (EC ) B05 - v gsartor ucleotidi e coenzimi

90 Metabolismo del Glicogeno Glicogenolisi e glicogenosintesi Glicogeno (Amido) Il glicogeno è un polimero del glucoso, I monomeri sono legati con legami glicosidici 1α 4 nelle catene principali e 1α 6 nelle ramificazioni. È un sistema di accumulazione del glucoso sottoforma di granuli in genere nel fegato. C 2 C 2 C 2 n 1α 1µm C 2 C 2 C 2 6C 2 C 2 C 2 n 1α 4 B05 - v gsartor ucleotidi e coenzimi
91 Glicogeno (Amido) Il glicogeno è un polimero del glucoso, I monomeri sono legati con legami glicosidici 1α 4 nelle catene principali e 1α 6 nelle ramificazioni. È un sistema di accumulazione del glucoso sottoforma di granuli in genere nel fegato. C 2 C 2 C 2 1α n C 2 C 2 C 2 6C 2 C 2 C 2 1α 4 n B05 - v gsartor ucleotidi e coenzimi 181 Glicogeno (Amido) La struttura dei granuli di glicogeno permette una rapida mobilizzazione (scissione) delle catene polisaccaridiche poiché vi sono molte estremità diverse attaccabili. C 2 C 2 C 2 1α n C 2 C 2 C 2 6C 2 C 2 C 2 1α 4 n B05 - v gsartor ucleotidi e coenzimi

92 Catabolismo del glicogeno La catena lineare polisaccaridica viene scissa nei monomeri (come glucoso-1-fosfato) ad opera dalla glicogenofosforilasi (EC ): C 2 C 2 n C 2 R Glicogeno fosforilasi EC C C 2 n C 2 R Fosforolisi R R' R + + R' Idrolisi R R' R + + R' Date le dimensioni del sito l enzima riesce a tagliare il legame 1α 4 fino a quattro residui dal legame 1α 6. B05 - v gsartor ucleotidi e coenzimi 183 Glicogeno fosforilasi EC B05 - v gsartor ucleotidi e coenzimi


93 Glicogeno (n-1) Metabolismo dei composti del Carbonio /04/ :30 Glicogeno fosforilasi EC B05 - v gsartor ucleotidi e coenzimi 185 Glicogeno fosforilasi EC Base di hiff Lisina Glicogeno (n) + iridossalfosfato R + C 3 n C 2 C 2 C 2 Lisina + R C 2 n C C 3 iridossale Glucoso-1-fosfato C 2 B05 - v gsartor ucleotidi e coenzimi
 + iridossalfosfato R + C 3 n C 2 C 2 C 2 Lisina Glicogeno (n-1) + R C 2 n C 2 + + C 3 iridossale Glucoso-1-fosfato C 2 B05 - v 1.8.")

94 Glicogeno fosforilasi EC Base di hiff Lisina Glicogeno (n) + iridossalfosfato R + C 3 n C 2 C 2 C 2 Lisina Glicogeno (n-1) + R C 2 n C C 3 iridossale Glucoso-1-fosfato C 2 B05 - v gsartor ucleotidi e coenzimi 187 Enzima deramificante EC oiché la glicogeno fosforilasi non riesce a scindere tutti i legami 1α 4 e 1α 6 interviene l enzima deramificante con il suo dominio transferasico, scinde una catena di tre monomeri dal monomero dove vi è la ramificazione e lega i tre monomeri alla catena lineare adiacente, Il residuo legato in 1α 6 viene idrolizzato dalla funzione deramificante con produzione di glucosio. B05 - v gsartor ucleotidi e coenzimi
95 Glicogeno fosforilasi EC Glicogeno fosforilasi C 2 C 2 C 2 C 2 C 2 C 2 1α C 2 6 C 2 1α 4 4 C 2 C 2 Terminale non riducente (legame glicosidico) B05 - v gsartor ucleotidi e coenzimi 189 Glicogeno fosforilasi EC Glicogeno fosforilasi C 2 C 2 C 2 C 2 C 2 C 2 1α C 2 1α 6 C C 2 C 2 B05 - v gsartor ucleotidi e coenzimi

96 Glicogeno fosforilasi EC Ramificazione limite Glicogeno fosforilasi C 2 C C 2 C 2 C 2 C 2 1α C 2 6C 2 1α 4 4 C 2 C 2 Glucoso-1-fosfato B05 - v gsartor ucleotidi e coenzimi 191 Glicogeno fosforilasi EC Ramificazione limite Glicogeno fosforilasi C 2 C 2 C 2 C 2 C 2 1α C 2 6 C 2 1α 4 4 C 2 C 2 B05 - v gsartor ucleotidi e coenzimi


97 Enzima deramificante EC Enzima deramificante Transferasi C 2 ito di glicosidazione C 2 C 2 C 2 C 2 1α C 2 6C 2 1α 4 4 C 2 C 2 B05 - v gsartor ucleotidi e coenzimi 193 Enzima deramificante EC Enzima deramificante Transferasi C 2 C 2 C 2 C 2 C 2 C 2 1α 1α 6 C C 2 C 2 ito di glicosidazione B05 - v gsartor ucleotidi e coenzimi


98 Enzima deramificante EC Transferasi Enzima deramificante C 2 C 2 C 2 C 2 C 2 1α C 2 6 C 2 1α 4 4 ito di glicosidazione 2 C 2 C 2 B05 - v gsartor ucleotidi e coenzimi 195 Enzima deramificante EC Enzima deramificante Transferasi Transferasi C 2 Glucoso C 2 C 2 C 2 1α C 2 ito di glicosidazione C 2 1α 6 C C 2 C 2 B05 - v gsartor ucleotidi e coenzimi


99 Glicogeno fosforilasi EC Glicogeno fosforilasi C 2 C 2 C 2 C 2 C 2 1α 6 C C 2 C 2 B05 - v gsartor ucleotidi e coenzimi 197 Glicogeno fosforilasi EC Glicogeno fosforilasi C 2 C 2 C 2 C 2 C 2 1α 6 C C 2 C B05 - v gsartor ucleotidi e coenzimi
 che catalizza la conversione attraverso la formazione di un intermedio glucoso-1,6-bifosfato.")
100 Glucoso-1-fosfato Il prodotto della glicogenolisi è principalmente glucoso-1- fosfato il quale entra nella glicolisi dopo essere stato convertito in glucoso-6-fosfato da una fosfoglucomutasi (EC ) che catalizza la conversione attraverso la formazione di un intermedio glucoso-1,6-bifosfato. Il meccanismo è simile a quello della fosfoglicerato mutasi il quale usa, invece, un residuo di istidina. Fosfoserina C 2 Glucoso-1-fosfato Fosfoserina erina C 2 Glucoso-1,6-bifosfato (non rilasciato) C 2 Glucoso-6-fosfato GLICLII B05 - v gsartor ucleotidi e coenzimi 199 Fosfoglucomutasi EC B05 - v gsartor ucleotidi e coenzimi
 è il precursore per la sintesi di glicogeno.")
101 Fosfoglucomutasi EC Glucoso-1,6-bifosfato er B05 - v gsartor ucleotidi e coenzimi 201 Glicogenosintesi Il glucoso uridin-difosfato (UDG) è il precursore per la sintesi di glicogeno. Un residuo di glucoso è addizionato al glicogeno e viene rilasciato un UD. Gli zuccheri nucleotidi difosfati sono i precursori della sintesi di carboidrati complessi, glicoproteine ecc. Viene sintetizzato da glucoso-1-fosfato e UT ad opera della UD- Glucoso pirofosforilasi (EC ). + UD-Glucoso pirofosforilasi EC B05 - v gsartor ucleotidi e coenzimi
102 Amido er produrre l amido viene sintetizzato amilosio attraverso la formazione di AD-glucoso Viene sintetizzato da glucoso-1-fosfato e AT ad opera della AD- Glucoso pirofosforilasi (EC ) con meccanismo analogo al glicogeno. 2 + AD-Glucoso pirofosforilasi EC B05 - v gsartor ucleotidi e coenzimi 203 Glicogenosintesi Il glicogeno si forma a partire da una proteina primer, la glicogenina, alla quale si lega il primo residuo di glucoso attraverso un residuo di tirosina. L enzima che si occupa della catalisi è la stessa glicogenina (EC ) (autoglicosilazione). + R R + R B05 - v gsartor ucleotidi e coenzimi R

103 Glicogenosintesi Il processo viene ripetuto fino a che si forma una corta catena di glucoso (fino a cinque residui). + R x4 Glicogenina EC R + 4 R R B05 - v gsartor ucleotidi e coenzimi 205 Glicogenina EC B05 - v gsartor ucleotidi e coenzimi

104 Glicogenina EC UD-glucoso Tyr B05 - v gsartor ucleotidi e coenzimi 207 Glicogenosintesi uccessivamente interviene la Glicogeno sintasi (EC ) per l allungamento della catena. È un complesso di una subunità catalitica e della proteina glicogenina. + R 4 R + Glicogeno sintasi EC R R 4 B05 - v gsartor ucleotidi e coenzimi


105 Glicogenosintesi La ramificazione 1α 6 viene catalizzata da un enzima ramificante (EC ). Lo stesso enzima è responsabile della conversione di amilosio in amilopectina e dell ulteriore ramificazione dell amilopectina C 2 C 2 C 2 C 2 1α C 2 C 2 C 2 1α 4 6 C 2 C 2 Enzima ramificante EC C 2 C 2 C 2 C 2 1α C 2 C 2 C 2 6 1α 4 C 2 C 2 B05 - v gsartor ucleotidi e coenzimi 209 Enzima ramificante EC B05 - v gsartor ucleotidi e coenzimi
106 Controllo Il metabolismo dei carboidrati ha ruoli diversi nel muscolo e nel fegato. el muscolo: per generate AT el fegato: mantenere il livello ematico di glucoso (produce ed esporta glucoso o importa ed immagazzina glucoso in risposta alla glicemia). La sintesi e degradazione del glucosio e del glicogeno sono quindi sottoposte al controllo ormonale attraverso il sistema del cam. B05 - v gsartor ucleotidi e coenzimi 211 Controllo del metabolismo del glicogeno C UD-Glucoso Glucoso-1-fosfato C 2 n C 2 R C 2 C (Glicogeno) n (Glicogeno) n+1 Glicogeno fosforilasi n UD Glicogeno + sintasi + i C 2 R ia la sintesi che la scissione del glicogeno sono processi termodinamicamente spontanei, se le due reazioni fossero attive simultaneamente si avrebbe la perdita netta di un legame ad alta energia per ciclo (si forma UD-Glucoso). er prevenire questa eventualità la glicogeno sintasi e la glicogeno fosforilasi sono regolate reciprocamente da effettori allosterici e dalla fosforilazione. B05 - v gsartor ucleotidi e coenzimi
107 C 2 C 2 C 2 C 2 C 2 C 2 C 2 C 2 Metabolismo dei composti del Carbonio /04/ :30 Regolazione allosterica La glicogeno fosforilasi nel muscolo è regolata da AM, AT e glucoso-6-fosfato. (un isoenzima diverso nel fegato è meno sensibile a questi controlli allosterici). AM (presente quando l AT manca) attiva la fosforilasi promuovendone la conformazione R. AT e glucoso-6-fosfato, che spiazzano l AM dalla fosforilasi, la inibiscono promuovendo la conformazione T. Quindi la rottura del glicogeno è inibita quando sono presenti elevate concentrazioni di AT e glucoso-6-fosfato. La glicogeno sintasi è attivata dal glucoso-6-fosfato (effetto opposto nella glicogeno fosforilasi). Quindi la glicogeno fosforilasi è attiva quando un alto livello ematico di glucoso porta ad un elevato livello cellulare di glucoso-6-fosfato. B05 - v gsartor ucleotidi e coenzimi 213 Regolazione allosterica Il glucoso-6-fosfato può entrare nella glicolisi o (nel fegato) essere defosforilato ad opera della glucoso-6- fosfatasi e rilasciato nel sangue. In quasi tutti gli altri tessuti manca questo enzima. Glicogeno C 2 Glicogeno sintetasi C 2 Glucosio Esochinasi o glucochinasi Glucoso-6-fosfatasi Glicogeno fosforilasi C Glucosio-1- C Glucosio-6- C 2 Glucosio AD AM AT Extracellulare GLICLII C 3 iruvato B05 - v gsartor ucleotidi e coenzimi
108 Regolazione ormonale (covalente) FFRILAZIE Gli ormoni Glucagone e adrenalina attivano i recettori di membrana accoppiati alla proteina G (GCR) i quali innescano la cascata del cam che porta alla forsforilazione di proteine bersaglio. Entrambi gli ormoni sono prodotti in risposta a bassi livelli ematici di glucoso. Il glucagone è sintetizzato dalle cellule α delpancreas e attiva la formazione di cam nel fegato. L adrenalina attiva la formazione di cam nel muscolo. B05 - v gsartor ucleotidi e coenzimi 215 Regolazione ormonale (covalente) FFRILAZIE La cascata del cam porta alla fosforilazione di una er nella glicogeno fosforilasi promuovendone la forma R attiva. L enzima fosforilato è meno sensibile agli inibitori allosterici. Quindi, anche se AT e glucoso-6-fosfato sono a valori elevati la fosforilasi è ancora attiva. Il glucoso-1-fosfato prodotto dal glicogeno nel fegato può essere convertito a glucoso ematico. La regolazione ormonale permette alle necessità dell organismo di prevalere sulle necessità della cellula. B05 - v gsartor ucleotidi e coenzimi
109 Regolazione ormonale (covalente) FFRILAZIE La cascata del cam ha effetti opposti nella glicogeno sintesi. La glicogeno sintasi è anch essa fosforilata dalla cascata, ma viene promossa la conformazione b, meno attiva. Quindi la cascata del cam inibisce la sintesi di glicogeno. Invece di essere convertito in glicogeno il glucoso-1-fosfato può esser defosforilato e rilasciato nel sangue. B05 - v gsartor ucleotidi e coenzimi 217 rmone (Adrenalina o glucagone) Cascata di segnali G α -GT Adenilato ciclasi (inattiva) Adenilato ciclasi (attiva) AT roteina chinasi A (inattiva) Forma a attiva ed indipendente da effettori allosterici Forma b meno attiva e dipendente da fattori allosterici Fosforilasi chinasi (b-inattiva) i AM cam + i Fosfodiesterasi roteina chinasi A (attiva) AT AD Fosforilasi chinasi (a-attiva) Fosfatasi AT Glicogeno AD Glicogeno Fosforilasi Fosforilasi (b-allosterica) Fosfatasi (a-attiva) B05 - v gsartor ucleotidi e coenzimi 218 i Glucoso Glicogeno 109
110 rmone (Adrenalina o glucagone) Cascata di segnali G α -GT Adenilato ciclasi (inattiva) Glicogeno Adenilato ciclasi (attiva) AT Glucoso roteina chinasi A (inattiva) Fosforilasi chinasi (b-inattiva) i AM cam + i Fosfodiesterasi roteina chinasi A (attiva) AT i AD Fosforilasi chinasi (a-attiva) Fosfatasi AT Glicogeno sintasi (b-attiva) AD Fosfatasi Glicogeno sintasi (a-inattiva) B05 - v gsartor ucleotidi e coenzimi 219 Insulina L insulina è un ormone prodotto dalle cellule β del pancreas in risposta ad alti livelli ematici di glucoso. Attiva una cascata di segnali separata che porta alla attivazione delle fosfoproteina fosfatasi. Queste fosfatasi catalizzano la rimozione del fosfato sia dalla fosforilasi chinasi che dalla glicogeno fosforilasi che dalla glicogeno sintasi. Quindi l insulina antagonizza gli effetti della cascata del cam indotta da glucagone e adrenalina. B05 - v gsartor ucleotidi e coenzimi
 i Fosfodiesterasi roteina chinasi A (attiva) Fosfatasi AD Fosforilasi chinasi (a-attiva) B05 - v 1.8.")

111 rmone (Adrenalina o glucagone) Cascata di segnali Insulina istema Recettore, roteina G (G α -GT) Adenilato ciclasi (attiva) AM Adenilato ciclasi (inattiva) AT roteina chinasi A (inattiva) cam + i AT Fosforilasi chinasi (b-inattiva) i Fosfodiesterasi roteina chinasi A (attiva) Fosfatasi AD Fosforilasi chinasi (a-attiva) B05 - v gsartor ucleotidi e coenzimi 221 AT Glicogeno Fosforilasi (b-allosterica) i + + Fosfatasi AD Glicogeno Fosforilasi (a-attiva) Ca ++ Anche lo ione Ca ++ gioca un ruolo nel metabolismo del glicogeno nel muscolo. Al momento della contrazione muscolare lo ione Ca ++ viene rilasciato dal reticolo sarcoplasmatico della cellula muscolare attraverso l attivazione di un canale specifico. Il Ca ++ rilasciato nel citoplasma attiva l interazione actina/miosina el muscolo la fosforilasi chinasi ha un dominio calmodulinico nella subunità δ che lega il Ca++ e attiva parzialmente (modula) la fosforilasi chinasi. La fosforilazione indotta dalla cascata del cam innescata dall adrenalina porta ad una ulteriore attivazione. Questo processo porta al rilascio di glucoso dal glicogeno che, attraverso la glicolisi, porta alla produzione di AT. B05 - v gsartor ucleotidi e coenzimi
112 rmone (Adrenalina o glucagone) Cascata di segnali istema Recettore, roteina G (G α -GT) Adenilato ciclasi (attiva) AM Adenilato ciclasi (inattiva) Fosforilasi chinasi (b-inattiva) AT roteina chinasi A (inattiva) cam + i Fosfodiesterasi roteina chinasi A (attiva) B05 - v gsartor ucleotidi e coenzimi 223 AT AD Fosforilasi chinasi (a-attiva) Fosfatasi i AT AD Glicogeno Fosforilasi (b-allosterica) Ca ++ Fosforilasi chinasi Ca ++ (parz. attiva) Ca ++ i Fosfatasi Glicogeno Fosforilasi (a-attiva) Via dei pentosi fosfati Altri nomi: Via del fosfogluconato hunt dell esoso monofosfato La parte lineare della via porta alla ossidazione e decarbossilazione di glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C). Il resto della via converte ribuloso-5-fosfato a riboso-5-fosfato (5C) oppure a gliceraldeide-3-fosfato (3C) e fruttoso-6-fosfato (6C) Con produzione di AD B05 - v gsartor ucleotidi e coenzimi
113 Da glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C) Glucoso-6-fosfato isomerasi (EC ) AD AD + Glucoso-6-fosfato deidrogenasi (EC ) α-d-glucoso-6-fosfato β-d-glucoso-6-fosfato 6-fosfo-D-glucono-1,5-lattone 6-fosfogluconolattonasi (EC ) D-ribuloso-5-fosfato C 2 AD AD + Fosfogluconato deidrogenasi (EC ) 6-fosfo-D-gluconato B05 - v gsartor ucleotidi e coenzimi 225 Da glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C) Glucoso-6-fosfato isomerasi (EC ) AD AD + Glucoso-6-fosfato deidrogenasi (EC ) α-d-glucoso-6-fosfato β-d-glucoso-6-fosfato 6-fosfo-D-glucono-1,5-lattone Il glucoso-6-fosfato viene convertito dalla forma α alla forma β attraverso la glucoso-6-fosfato isomerasi La glucoso-6-fosfato deidrogenasi catalizza l ossidazione del gruppo aldeidico (semiacetale) del glucoso-6-fosfato (in C1) ad acido carbossilico. i forma un legame estereo (lattone). Il AD + serve come accettore di elettroni, viene prodotto AD. B05 - v gsartor ucleotidi e coenzimi


114 Glucoso-6-fosfato isomerasi EC QY4 B05 - v gsartor ucleotidi e coenzimi 227 Glucoso-6-fosfato isomerasi EC Ca ++ G6 B05 - v gsartor ucleotidi e coenzimi


115 Glucoso-6-fosfato deidrogenasi EC Asp177 is B05 - v gsartor ucleotidi e coenzimi 229 Glucoso-6-fosfato deidrogenasi EC AD + B05 - v gsartor ucleotidi e coenzimi

116 Glucoso-6-fosfato deidrogenasi EC Ca ++ G6 mologo del AD + B05 - v gsartor ucleotidi e coenzimi 231 Regolazione della glucoso-6-fosfato deidrogenasi La reazione catalizzata dalla glucoso-6-fosfato deidrogenasi è un passo obbligatorio della via dei pentosi fosfati e la sua attività è regolata dalla presenza di AD +. Il AD è utilizzato nelle vie biosintetiche e viene convertito in AD + Il AD + stimola la via dei pentosi fosfati che porta alla formazione di AD. B05 - v gsartor ucleotidi e coenzimi

117 Da glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C) La 6-fosfogluconolattonasi catalizza l idrolisi del legame estereo con apertura dell anello. Il prodotto è 6- fosfogluconato. L apertura dell anello avverrebbe anche in assenza di enzima. La lattonasi aumenta la velocità della reazione favorendo la scomparsa del 6-fosfogluconolattone che è altamente reattivo e potenzialmente tossico. 6-fosfo-D-glucono-1,5-lattone 6-fosfogluconolattonasi (EC ) 6-fosfo-D-gluconato B05 - v gsartor ucleotidi e coenzimi fosfogluconolattonasi EC Citrato B05 - v gsartor ucleotidi e coenzimi
 è ossidato a chetone. Ciò promuove la perdita del carbossile dal C1 come C 2. Il AD + serve da ossidante.")
118 Da glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C) La fosfogluconato deidrogenasi (EC ) catalizza la decarbossilazione ossidativa del 6-fosfogluconato. i forma il chetone a 5-C ribuloso-5-fosfato. Il gruppo al C3 (C2 del prodotto) è ossidato a chetone. Ciò promuove la perdita del carbossile dal C1 come C 2. Il AD + serve da ossidante. D-ribuloso-5-fosfato C 2 AD AD + Fosfogluconato deidrogenasi (EC ) 6-fosfo-D-gluconato B05 - v gsartor ucleotidi e coenzimi 235 Fosfogluconato deidrogenasi EC fosfogluconato B05 - v gsartor ucleotidi e coenzimi
119 Fosfogluconato deidrogenasi EC mologo del AD + B05 - v gsartor ucleotidi e coenzimi 237 Da glucoso-6-fosfato (6C) a ribuloso-5-fosfato (5C) Glucoso-6-fosfato isomerasi (EC ) AD AD + Glucoso-6-fosfato deidrogenasi (EC ) α-d-glucoso-6-fosfato β-d-glucoso-6-fosfato 6-fosfo-D-glucono-1,5-lattone D-ribuloso-5-fosfato AD 6-fosfogluconolattonasi (EC ) C 2 AD + Fosfogluconato deidrogenasi (EC ) 6-fosfo-D-gluconato B05 - v gsartor ucleotidi e coenzimi
120 AD + e AD La riduzione del AD + (così come del AD + ) coinvolge il trasferimento di 2 e e 1 + alla porzione nicotinamidica della molecola (come - ). Il AD, prodotto dalla via dei pentosi fosfati è la molecola riducente nelle vie sintetiche (anaboliche) della cellula. Il AD + serve come accettore di elettroni nelle vie cataboliche (demolizione) dove i metaboliti sono ossidati. Il AD che si forma è riossidato nella catena respiratoria per la produzione di AT e B05 - v gsartor ucleotidi e coenzimi 239 AD + e AD AD + AD + AD + e AD + differiscono solo per la presenza del fosfato legato al riboso nel AD +. er quanto riguarda l attività redox non vi è sostanziale differenza. La presenza del fosfato serve per il riconoscimento di uno o dell altro dagli enzimi. Ciò permette la separazione delle vie cataboliche e anaboliche nella cellula. B05 - v gsartor ucleotidi e coenzimi
121 Il resto della via converte il ribuloso-5-fosfato a riboso-5-fosfato (5C) e xiluloso-5-fosfato attraverso la catalisi effettuata da epimerasi e isomerasi e quindi a gliceraldeide-3-fosfato (3C), sedoeptuloso- 7-fosfato (7C), eritroso-4-fosfato (4C) e fruttoso-6-fosfato (6C) attraverso transaldolasi e transchetolasi B05 - v gsartor ucleotidi e coenzimi 241 D-xiluloso-5-fosfato Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato Riboso-5-fosfato isomerasi (EC ) D-riboso-5-fosfato Transchetolasi (EC ) + D-gliceraldeide-3-fosfato D-sedoeptuloso-7-fosfato Transaldolasi (EC ) D-gliceraldeide- 3-fosfato Transchetolasi (EC ) D-eritroso-4-fosfato + D-fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi
 5C Riboso-5-fosfato isomerasi (EC 5.3.1.6) 7C D-riboso-5-fosfato 5C D-sedoeptuloso-7-fosfato Transchetolasi (EC 2.2.1.1) 3C 4C + 6C D-fruttoso-6-fosfato D-gliceraldeide- 3-fosfato D-eritroso-4-fosfato GLICLII - GLUCEGEI intesi di UCLETIDI FFATI B05 - v 1.")
 D-riboso-5-fosfato truttura ad α β barrel B05 - v 1.8.1 gsartor 2001-2018 ucleotidi e coenzimi 244 122")
122 GLICLII - GLUCEGEI D-xiluloso-5-fosfato 5C Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato Transchetolasi (EC ) 3C + D-gliceraldeide-3-fosfato Transaldolasi (EC ) 5C Riboso-5-fosfato isomerasi (EC ) 7C D-riboso-5-fosfato 5C D-sedoeptuloso-7-fosfato Transchetolasi (EC ) 3C 4C + 6C D-fruttoso-6-fosfato D-gliceraldeide- 3-fosfato D-eritroso-4-fosfato GLICLII - GLUCEGEI intesi di UCLETIDI FFATI B05 - v gsartor ucleotidi e coenzimi 243 Epimerasi (EC ) D-xiluloso-5-fosfato Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato Riboso-5-fosfato isomerasi (EC ) D-riboso-5-fosfato truttura ad α β barrel B05 - v gsartor ucleotidi e coenzimi
 D-ribuloso-5-fosfato Riboso-5-fosfato isomerasi (EC 5.3.1.6) D-riboso-5-fosfato B05 - v 1.")
 D-ribuloso-5-fosfato D-riboso-5-fosfato + Riboso-5-fosfato isomerasi (EC 5.")

123 Isomerasi (EC ) D-xiluloso-5-fosfato Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato Riboso-5-fosfato isomerasi (EC ) D-riboso-5-fosfato B05 - v gsartor ucleotidi e coenzimi 245 Isomerasi (EC ) D-xiluloso-5-fosfato Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato D-riboso-5-fosfato + Riboso-5-fosfato isomerasi (EC ) + Intermedio enediolato B05 - v gsartor ucleotidi e coenzimi
 sia al riboso-5- fosfato che all eritroso-4- fosfato (aldosi). D-xiluloso-5-fosfato B05 - v 1.8.")
124 Transchetolasi EC Le transchetolasi e le transaldolasi catalizzano, rispettivamente, il trasferimento di frammenti di due o tre atomi di carbonio da un chetoso donatore ad un aldoso accettore. La transchetolasi trasferisce un frammento 2-C dal xiluloso-5-fosfato (chetoso) sia al riboso-5- fosfato che all eritroso-4- fosfato (aldosi). D-xiluloso-5-fosfato B05 - v gsartor ucleotidi e coenzimi 247 Transchetolasi (EC ) + D-gliceraldeide-3-fosfato D-riboso-5-fosfato D-sedoeptuloso-7-fosfato Transchetolasi EC D-xiluloso-5-fosfato D-riboso-5-fosfato Ca ++ T Eritroso-4-fosfato Transchetolasi (EC ) + D-gliceraldeide-3-fosfato D-sedoeptuloso-7-fosfato B05 - v gsartor ucleotidi e coenzimi
 derivato dalla vitamina B 1.")
.")
125 Transchetolasi EC D-xiluloso-5-fosfato D-eritroso-4-fosfato Ca ++ T Eritroso-4-fosfato Transchetolasi (EC ) + D-gliceraldeide-3-fosfato D-sedoeptuloso-7-fosfato B05 - v gsartor ucleotidi e coenzimi 249 Tiaminapirofosfato La transchetolasi utilizza come gruppo prostetico il T (tiaminapirofosfato) derivato dalla vitamina B 1. Il T si lega nel sito attivo piegato a V. Il protone tra gli atomi di azoto e zolfo nell anello tiazolico è acido e dissocia. T Il gruppo aminico dell anello aminopiridino è vicino al protone dissociabile e serve come accettore (base). Il trasferimento del protone è favorito dalla presenza di un residuo di Glu adiacente all anello pirimidinico. 3 C Ca ++ C Eritroso-4-fosfato B05 - v gsartor ucleotidi e coenzimi
126 Tiaminapirofosfato Il carbanione attacca il carbonile del xiluloso-5- per formare un intermedio di addizione. Il + nell anello tiazolico agisce come un accettore di elettroni favorendo la rottura del legame C-C. 3 C 3 C C 3 + C C C 2 C 2 - Intermedio D-xiluloso-5-fosfato - B05 - v gsartor ucleotidi e coenzimi 251 Tiaminapirofosfato i forma l aldoso a tre atomi di carbonio gliceraldeide-3- che viene rilasciato, il frammento a 2- C rimane legato al T. Il frammento 2-C condensa con un aldoso (eritroso-4- o riboso-5-) per formare un chetoso-. Il trasferimento del frammento 2-C sul riboso-5- forma il sedoeptuloso-7-fosfato. 3 C 3 C C 2 D-gliceraldeide-3-fosfato - D-riboso-5-fosfato D-sedoeptuloso-7-fosfato B05 - v gsartor ucleotidi e coenzimi
127 Tiaminapirofosfato i forma l aldoso a tre atomi di carbonio gliceraldeide-3- che viene rilasciato, il frammento a 2- C rimane legato al T. Il frammento 2-C condensa con un aldoso (eritroso-4- o riboso-5-) per formare un chetoso-. Il trasferimento del frammento 2-C sul eritroso-4- forma il fruttoso-6-7-fosfato. 3 C 3 C + C D-gliceraldeide-3-fosfato - D-eritroso-4-fosfato D-fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi 253 Transaldolasi EC La transaldolasi trasferisce la porzione 3-C (diidrossiacetone) dal sedoeptuloso-7-fosfato alla gliceraldeide-3-fosfato. Il gruppo ε-aminico della transaldolasi reagisce con il carbonile del sedoeptuloso-7- fosfato. Lys 2 + D-sedoeptuloso-7-fosfato Intermedio base di hiff i forma la base di hiff protonata. i libera l eritroso-4-fosfato. La base di hiff stabilizza il carbanione in C3. La reazione prosegue con l attacco del carbanione al carbonile della gliceraldeide-3-fosfato per formare fruttoso-6-fosfato. + C D-gliceraldeide-3-fosfato + D-eritroso-4-fosfato D-fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi

128 Transaldolasi EC La transaldolasi ha una struttura α,β barrel. Lys Lys base di hiff ridotta B05 - v gsartor ucleotidi e coenzimi 255 Transaldolasi EC La transaldolasi ha una struttura α,β barrel. B05 - v gsartor ucleotidi e coenzimi
129 Bilancio ella via dei pentosi fosfati entrano tre pentosi (15 atomi di carbonio) che vengono convertiti in due esosi ed un trioso. C 5 + C 5 C 3 + C 7 (Transchetolasi) C 3 + C 7 C 6 + C 4 (Transaldolasi) C 5 + C 4 C 6 + C 3 (Transchetolasi) 3 C 5 2 C 6 + C 3 Il glucoso-6-fosfato può esser rigenerato sia da gliceraldeide- 3-fosfato che dal fruttoso-6-fosfato attraverso la Gluconeogenesi. B05 - v gsartor ucleotidi e coenzimi 257 GLICLII - GLUCEGEI D-xiluloso-5-fosfato 5C 3 1 Ribuloso-fosfato 3-epimerasi (EC ) D-ribuloso-5-fosfato 1 2 Transchetolasi (EC ) 3C + D-gliceraldeide-3-fosfato Transaldolasi (EC ) 5C Riboso-5-fosfato isomerasi (EC ) 7C D-riboso-5-fosfato 5C D-sedoeptuloso-7-fosfato Transchetolasi (EC ) 3C 4C + 6C D-fruttoso-6-fosfato D-gliceraldeide- 3-fosfato D-eritroso-4-fosfato 2 3 GLICLII - GLUCEGEI intesi di UCLETIDI FFATI B05 - v gsartor ucleotidi e coenzimi
130 trategia A secondo dei bisogni della cellula per riboso-5-fosfato, AD, e AT, la via dei entosi fosfati opera in vari modi per massimizzare la concentrazione dei diversi prodotti. B05 - v gsartor ucleotidi e coenzimi 259 intesi di riboso-5- e AD Duplicazione cellulare. Il ribuloso-5- fosfato viene convertito in riboso-5-fosfato, necessario per la sintesi di nucleotidi e acidi nucleici. Viene anche prodotto del AD. α-d-glucoso-6-fosfato D-xiluloso-5-fosfato D-gliceraldeide- 3-fosfato Ribuloso-fosfato 3-epimerasi (EC ) Transchetolasi (EC ) Glucoso-6-fosfato isomerasi (EC ) Transchetolasi (EC ) + D-gliceraldeide-3-fosfato Transaldolasi (EC ) D-eritroso-4-fosfato β-d-glucoso-6-fosfato Fosfogluconato deidrogenasi (EC ) D-ribuloso-5-fosfato + AD AD + Glucoso-6-fosfato deidrogenasi (EC ) AD Riboso-5-fosfato isomerasi (EC ) AD + C 2 6-fosfo-D-glucono-1,5-lattone D-riboso-5-fosfato D-sedoeptuloso-7-fosfato 6-fosfogluconolattonasi (EC ) 6-fosfo-D-gluconato D-fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi
131 intesi di AD massimizzata Attività sintetica della cellula. ia la gliceraldeide-3- fosfato che il fruttoso-6-fosfato possono essere convertiti in glucoso-6-fosfato per massimizzare la sintesi di AD. a-d-glucoso-6-fosfato D-xiluloso-5-fosfato Transchetolasi (EC ) + D-gliceraldeide- 3-fosfato D-eritroso-4-fosfato Glucoso-6-fosfato isomerasi (EC ) Ribuloso-fosfato 3-epimerasi (EC ) Transchetolasi (EC ) + D-gliceraldeide-3-fosfato Transaldolasi (EC ) β-d-glucoso-6-fosfato Fosfogluconato deidrogenasi (EC ) D-ribuloso-5-fosfato AD AD + Glucoso-6-fosfato deidrogenasi (EC ) AD Riboso-5-fosfato isomerasi (EC ) AD + C 2 6-fosfo-D-glucono-1,5-lattone D-riboso-5-fosfato D-sedoeptuloso-7-fosfato 6-fosfogluconolattonasi (EC ) 6-fosfo-D-gluconato D-fruttoso-6-fosfato B05 - v gsartor ucleotidi e coenzimi 261 intesi di AD e AT Energia La gliceraldeide-3- fosfato e il fruttoso-6- fosfato possono entrare nella glicolisi per la produzione di AT. Viene anche prodotto del AD. GLICLII AT α-d-glucoso-6-fosfato D-xiluloso-5-fosfato D-gliceraldeide- 3-fosfato Ribuloso-fosfato 3-epimerasi (EC ) Transchetolasi (EC ) Glucoso-6-fosfato isomerasi (EC ) Transchetolasi (EC ) + D-gliceraldeide-3-fosfato Transaldolasi (EC ) D-eritroso-4-fosfato β-d-glucoso-6-fosfato Fosfogluconato deidrogenasi (EC ) D-ribuloso-5-fosfato + AD AD + Glucoso-6-fosfato deidrogenasi (EC ) Riboso-5-fosfato isomerasi (EC ) AD + C 2 AD 6-fosfo-D-glucono-1,5-lattone D-riboso-5-fosfato D-sedoeptuloso-7-fosfato D-fruttoso-6-fosfato 6-fosfogluconolattonasi (EC ) 6-fosfo-D-gluconato B05 - v gsartor ucleotidi e coenzimi
 2 C 2 + 2 etanolo Glicolisi iruvato Acetil- Riboso-5-fosfato Ciclo dei pentosi 3-fosfogliceraldeide 2 AD AD")
132 ssigeno Aerobiosi e anaerobiosi Aerobiosi Glicogeno Metabolismo del Glicogeno C 2 C C Gluconeogensi Fermentazione lattica (Anaerobiosi) 2 Lattato D-Glucoso 2AD + 2i 2AD + Fruttoso-1,6-difosfato 2AT 2AD 2AD + 2 iruvato 3 C 2AD Fermentazione alcolica (Anaerobiosi) 2 C etanolo Glicolisi iruvato Acetil- Riboso-5-fosfato Ciclo dei pentosi 3-fosfogliceraldeide 2 AD AD + ssidazione aerobica AD + Ciclo di Krebs Fosforilazione ossidativa 6 C AD + i + + AT 132
 ssidazione dell acetil- a C 2 (Ciclo di Krebs) Trasferimento di elettroni e fosforilazione ossidativa (produzione di 2 e")
133 Aerobiosi In condizioni aerobiche il piruvato prodotto dalla glicolisi e dalla degradazione degli aminoacidi è ossidato a 2 e C 2 nella respirazione cellulare. Ciò avviene in tre stadi roduzione di acetil- (decarbossilazione del piruvato) ssidazione dell acetil- a C 2 (Ciclo di Krebs) Trasferimento di elettroni e fosforilazione ossidativa (produzione di 2 e AT con consumo di 2 ). B05 - v gsartor ucleotidi e coenzimi 265 B05 - v gsartor ucleotidi e coenzimi
134 iruvato deidrogenasi roduzione di acetil- (decarbossilazione del piruvato) Trasporto del piruvato Il piruvato è trasportato all interno della matrice mitocondriale dove viene ossidato ad acetil dal complesso enzimatico piruvato deidrogenasi. Il piruvato viene trasportato attraverso la membrana mitocondriale attraverso un trasportatore specifico che lo scambia con ioni -. La membrana esterna mitocondriale permette il passaggio a ioni e piccole molecole e contiene canali anionici voltaggio dipendenti (VDAC: voltage dependent anion channels). B05 - v gsartor ucleotidi e coenzimi

135 Mitocondrio B05 - v gsartor ucleotidi e coenzimi 269 Acetil- C 3C 3-2 Acetile β-mercaptoetanolamina Acido antotenico 3 C C 3C 3 Adenosina-3'-difosfato 2 Acetil- B05 - v gsartor ucleotidi e coenzimi
136 Acetil- - Acetile β-mercaptoetanolamina Acido pantotenico Adenosina-3 -difosfato Acetil- B05 - v gsartor ucleotidi e coenzimi 271 Complesso piruvato deidrogenasi Il complesso piruvato deidrogenasi è un gruppo di enzimi associati non covalentemente che catalizzano la decarbossilazione del piruvato e formazione di Acetil-. La razione forma contemporaneamente AD trasferendo uno ione - al AD +. Il AD passa gli elettroni alla catena respiratoria La reazione ha un G = kj/mol (essenzialmente irreversibile). AD+ AD C 3 T, Lipoato, FAD C 3 C 2 + Complesso piruvato deidrogenasi (E 1 + E 2 + E 3 ) B05 - v gsartor ucleotidi e coenzimi
137 Complesso piruvato deidrogenasi Il complesso piruvato deidrogenasi catalizza cinque reazionisequenziali, richiede tre enzimie cinque coenzimi. I cinque coenzimi sono: Il FAD e il AD + sono trasportatori di elettroni. La T trasferisce il gruppo acetile al lipoato. Il lipoato è trasportatore di elettroni e di acili. Il è il trasportatore di acili, lega in modo covalente il gruppo acilico attraverso un legame tioestere ad alta energia. B05 - v gsartor ucleotidi e coenzimi 273 I reagenti e i prodotti FAD AD+ C 3 C 2 Idrossietil-T C 3 T C T iruvato deidrogenasi E 1 Lipoamide R Diidrolipoil transacetilasi E 2 3 C Diidrolipoil deidrogenasi E 3 FAD R C 3 Diidrolipoamide AD R Acetil-diidrolipoamide B05 - v gsartor ucleotidi e coenzimi
138 Le cinque reazioni nel complesso FAD AD+ C 3 C 2 Idrossietil-T C 3 C T T 1 2 iruvato deidrogenasi E 1 Lipoamide 3 C R Diidrolipoil transacetilasi E 2 4 Diidrolipoil deidrogenasi E 3 FAD 3 R C 3 Diidrolipoamide 5 AD R Acetil-diidrolipoamide B05 - v gsartor ucleotidi e coenzimi 275 I tre enzimi del complesso FAD AD+ C 3 C 2 Idrossietil-T C 3 T C T iruvato deidrogenasi E 1 Lipoamide R Diidrolipoil transacetilasi E 2 3 C Diidrolipoil deidrogenasi E 3 FAD R C 3 Diidrolipoamide AD R Acetil-diidrolipoamide B05 - v gsartor ucleotidi e coenzimi
139 I cinque coenzimi FAD AD+ C 3 C 2 Idrossietil-T C 3 C T T iruvato deidrogenasi E 1 Lipoamide 3 C R Diidrolipoil transacetilasi E 2 Diidrolipoil deidrogenasi E 3 FAD R C 3 Diidrolipoamide AD R Acetil-diidrolipoamide B05 - v gsartor ucleotidi e coenzimi 277 I cinque coenzimi 3 C C C C 3 C Base di hiff ridotta Ribosio in forma aperta FAD (ossidato) C 3 R FAD + C 3 R FAD z (ridotto) Coenzima A rotone acido 2 + AD + 2 AD 2 R 2 3 C + 2 C 3 Tiamina pirofosfato (T) Lipoamide e - Diidrolipoamide B05 - v gsartor ucleotidi e coenzimi
140 I tre enzimi Il complesso piruvato deidrogenasi consiste in tre enzimi: piruvato deidrogenasi EC (E 1, arancio) (B), diidrolipoil transacetilasi EC (E 2, verde) (A), diidrolipoil deidrogenasi EC (E 3, violetto) (B). In E. coli il complesso consiste in 24 coppie di E 1, 24 coppie di E 2 e di 12 coppie di E 3. E 2 funziona come core del complesso (C). B05 - v gsartor ucleotidi e coenzimi 279 I tre enzimi gni coppia di E 2 contiene tre molecole di lipoato legate covalentemente. Il lipoato ha un braccio flessibile che trasporta le molecole di acetile da un sito attivo ad un altro. E 1 ha come coenzima il molecola di T ed E 3 ha come coenzima il FAD. B05 - v gsartor ucleotidi e coenzimi
. R' C 3 + R C C 3 R' C 3 R C 3 B05 - v 1.8.")
141 iruvato deidrogenasi (E 1 ) EC R' C 3 + R R' C 3 + R C + + R' C 3 + R C 3 T C 3 C 2 Gli intermedi rimangono legati al complesso. Il piruvato reagisce con il T legato a E 1 e viene decarbossilato al derivato idrossietil-t (carbanione reattivo stabilizzato per risonanza). R' C 3 + R C C 3 R' C 3 R C 3 B05 - v gsartor ucleotidi e coenzimi 281 iruvato deidrogenasi (E 1 ) EC Mg ++ T 1L8A B05 - v gsartor ucleotidi e coenzimi


142 iruvato deidrogenasi (E 1 ) EC Cavità Carbanione T 1L8A B05 - v gsartor ucleotidi e coenzimi 283 iruvato deidrogenasi (E 1 ) EC T 1I4 B05 - v gsartor ucleotidi e coenzimi


143 iruvato deidrogenasi (E 1 ) EC T 1I4 B05 - v gsartor ucleotidi e coenzimi 285 iruvato deidrogenasi (E 1 ) EC Cavità Carbanione 1I4 T B05 - v gsartor ucleotidi e coenzimi
144 Diidrolipoil transacetilasi (E 2 ) EC R' C 3 C + R' 3 + R' C 3 + R + R + C R C C 3 C 3 C 3 Lipoamide Diidroacetilipoamide Il gruppo idrossietile derivato da idrossietil-t è trasferito al lipoato (legato ad una is di E 2 ) come acetile, attraverso l attacco nucleofilo del carbanione sull atomo di zolfo della lipoamide. B05 - v gsartor ucleotidi e coenzimi 287 Diidrolipoil transacetilasi (E 2 ) EC C R' 3 + R C C 3 Diidroacetilipoamide Il braccio del lipoato si muove sul sito di legame del acetil- e ne transesterifica il gruppo formando Acetil. 3 C C 3 B05 - v gsartor ucleotidi e coenzimi


145 Diidrolipoil transacetilasi (E 2 ) EC Dominio catalitico B05 - v gsartor ucleotidi e coenzimi 289 Diidrolipoil transacetilasi (E 2 ) EC Dominio catalitico Acetil Lipoamide B05 - v gsartor ucleotidi e coenzimi
 EC1.8.1.4 AD + FAD 1LV B05 - v 1.")
146 Diidrolipoil deidrogenasi (E 3 ) EC FAD FAD FAD 2 AD + AD + + Il lipoato ridotto viene riossidato dall E 3 utilizzando il FAD che si riduce a FAD 2. Il FAD 2 viene riossidato dal AD + che si riduce a AD e + i rigenera la piruvato deidrogenasi B05 - v gsartor ucleotidi e coenzimi 291 Diidrolipoil deidrogenasi (E 3 ) EC AD + FAD 1LV B05 - v gsartor ucleotidi e coenzimi
 e li trasferisce al diidrolipoil deidrogenasi (E 3 ).")
147 Diidrolipoil deidrogenasi (E 3 ) EC AD + FAD B05 - v gsartor ucleotidi e coenzimi 293 La diidrolipoil transacetilasi (E 2 ) è centrale in questo meccanismo. Il braccio flessibile del lipoato: lega il gruppo acetile e lo trasferisce al e accetta due elettroni dalla piruvato deidrogenasi (E 1 ) e li trasferisce al diidrolipoil deidrogenasi (E 3 ). B05 - v gsartor ucleotidi e coenzimi
148 Legame del piruvato E 1 R' 3 C + R C C 3 R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi 295 Decarbossilazione del piruvato E 1 R' 3 C + R C C 3 C 2 R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi
149 Formazione di diidroacetillipoamide E 1 R' 3 C + R C 3 C R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi 297 Legame del Coenzima A E 1 R' 3 C + R C 3 C R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi
150 Formazione di Acetil- E 1 R' 3 C + R C 3 C R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi 299 ssidazione della diidrolipoamide E 1 R' 3 C + R C Cys R C 3 C 3 Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi
151 Riduzione del FAD E 1 R' 3 C + R C R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi 301 Legame del AD + E 1 R' 3 C + R C R + 2 R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi
152 Riduzione del AD + a AD + + E 1 R' 3 C + R C R R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi 303 Ritorno al punto di partenza E 1 R' 3 C + R C C 3 R C 3 C 3 Cys Cys E 3 E 2 B05 - v gsartor ucleotidi e coenzimi
153 3 C T E 1 E 2 E 3 FAD C 3 C 2 T E 1 E 2 E 3 FAD AD C AD + FAD T E 1 E 2 E 3 T E 1 E 2 E 3 FAD 2 FAD T E 1 E 2 E 3 C 3 B05 - v gsartor ucleotidi e coenzimi 305 Controllo della piruvato deidrogenasi Inibizione competitiva da prodotti AD compete con AD + in E 3 Acetil- compete con - in E 2 La concentrazione dei due coenzimi regola anche la direzione della catalisi di E 2 e E 3. egli eucarioti E 1 può essere fosforilato da una chinasi attivata dalla forma acetilata di E 2 La forma fosforilata di E 1 è inattiva mentre la forma defosforilata è attiva. B05 - v gsartor ucleotidi e coenzimi
154 Controllo della piruvato deidrogenasi T * C 3 3 C * E 1 E 2 E 3 FAD 3 C * 3 C T * E 1 E 2 E 3 FAD AD Ca ++ (alto Mg ++ ) iruvato, AD, K + iruvato deidrogenasi chinasi Mg ++Ca++ 2 iruvato deidrogenasi fosfatasi Acetil- AD C 2 AT T C * 3 C 3 * E 1 E 2 E 3 FAD C B05 - v gsartor ucleotidi e coenzimi 307 Controllo della piruvato deidrogenasi 3 C T T 3 C C C C 3 3 FAD C 3 2 CE 31 E 2 E 3 T C C 3 3 C E 3 1 E 2 E 3 FAD AD + + AD + 3 C 3 C FAD 2 T CE 31 E 2 E 3 3 C 3 C 3 C FAD T CE 31 E 2 E 3 C 3 C T C E 31 E 2 3 E 3 FAD C 3 B05 - v gsartor ucleotidi e coenzimi


155 Ciclo di Krebs Ciclo degli acidi tricarbossilici (TCA) Ciclo dell acido citrico ssidazione dell acetil- a C 2 Krebs e Lipmann remio obel per la Medicina 1953 B05 - v gsartor ucleotidi e coenzimi
156 Ciclo di Krebs Il ciclo di Krebs è al centro del metabolismo. Le vie degradative (catabolismo) lo alimentano, le vie sintetiche (anabolismo) ne usano i componenti. È una via AFIBLICA, opera infatti sia nel catabolismo che nell anabolismo cellulare. B05 - v gsartor ucleotidi e coenzimi 311 Ciclo di Krebs Tutte le reazioni avvengono nella matrice mitocondriale. ei mitocondri vi sono anche gli enzimi della fosforilazione ossidativa e quelli della ossidazione degli acidi grassi e degli aminoacidi. Acetil + 3 AD + + FAD + GD + i C AD + FAD 2 + GT B05 - v gsartor ucleotidi e coenzimi
157 Krebs and Johnson (1937) The role of citric acid in the intermediate metabolism in animal tissues. Enzymologica 4: B05 - v gsartor ucleotidi e coenzimi 313 Ciclo di Krebs AD + AD + + Acetil C 2 2 C 3 3 C iruvato ssalacetato Malato Citrato Fumarato 2 FAD 2 FAD uccinato cis-aconitato uccinil GT GD + i α-chetoglutarato C 2 AD + + AD + ssalosuccinato Isocitrato AD + AD C 2 B05 - v gsartor ucleotidi e coenzimi
158 Ciclo di Krebs AD + AD + + Acetil C 2 2 Malato Fumarato ssalacetato C 3 Citrato 3 C iruvato 2 FAD 2 FAD uccinato GT uccinil GD + i α-chetoglutarato C 2 AD + + AD + ssalosuccinato cis-aconitato Isocitrato AD + AD C 2 B05 - v gsartor ucleotidi e coenzimi 315 B05 - v gsartor ucleotidi e coenzimi
159 Ciclo di Krebs AD + + AD + Acetil C 2 2 C 3 3 C ssalacetato ucleotidi trifosfati FAD 2 FAD Equivalenti riducenti 2 2 GT GD + i 2C 2 C 2 AD + + AD + AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 317 Ciclo di Krebs AD + AD + + C 2 2 C 3 3 C Malato deidrogenasi EC Fumarasi EC Citrato sintasi EC FAD 2 FAD uccinato deidrogenasi EC uccinil sintetasi EC Aconitasi EC Aconitasi EC GT GD + i α-chetoglutarato deidrogenasi EC Isocitrato deidrogenasi EC AD + + C 2 AD + AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi
160 Ciclo di Krebs AD + AD + + C 2 2 C 3 3 C Malato deidrogenasi EC Fumarasi EC Citrato sintasi EC FAD 2 FAD uccinato deidrogenasi EC uccinil sintetasi EC Deidrogenasi Aconitasi EC Aconitasi EC GT GD + i α-chetoglutarato deidrogenasi EC Isocitrato deidrogenasi EC AD + + C 2 AD + AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 319 Ciclo di Krebs AD + AD + + C 2 FAD 2 FAD GT GD + i 2 uccinil sintetasi EC AD + + Malato deidrogenasi EC Fumarasi EC uccinato deidrogenasi EC C 2 AD α-chetoglutarato deidrogenasi EC C 3 Citrato sintasi EC Isocitrato deidrogenasi EC Aconitasi EC AD + AD C Aconitasi EC C 2 B05 - v gsartor ucleotidi e coenzimi




161 Ciclo di Krebs AD + AD + + C 2 2 C 3 3 C FAD 2 FAD 6 2 GT GD + i 4 AD + + C 2 AD + AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 321 Ciclo di Krebs ALIMETAZIE RIRITI del CICL ECD A di RDUZIE di EERGIA5 1 REARAZIE 2 del CICL 3 4 RIM A di RDUZIE di EERGIA B05 - v gsartor ucleotidi e coenzimi
162 Ciclo di Krebs 2 C 4 C 4 AD + + AD + C 4 C 2 C 3 C 2 C 6 3 C 2 FAD 2 FAD C 4 C 6 2 GT GD + i C 4 C C 2 1 AD + + AD + C 5 C 6 C 6 AD + AD + + C C 2 1 B05 - v gsartor ucleotidi e coenzimi 323 Citrato sintasi EC FAD 2 FAD 2 Fumarasi EC uccinato deidrogenasi EC uccinil sintetasi EC AD + + AD + Malato deidrogenasi EC ssalacetato Acetil C 2 C 3 Citrato sintasi EC Citrato Aconitasi EC C Aconitasi EC GT GD + i AD + + α-chetoglutarato deidrogenasi EC C 2 AD + Isocitrato deidrogenasi EC AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi


163 Citrato sintasi EC FAD 2 FAD 2 Fumarasi EC uccinato deidrogenasi EC uccinil sintetasi EC AD + + AD + Malato deidrogenasi EC ssalacetato Acetil C 2 C 3 Citrato sintasi EC Citrato Aconitasi EC C Aconitasi EC GT α-chetoglutarato deidrogenasi GD + i Isocitrato Quando lega ossalacetato EC cambia conformazione, deidrogenasi EC i crea il sito per acetil-, C 2 L ossalacetato non è più accessibile all acqua. AD + AD + + AD + + AD + C 2 B05 - v gsartor ucleotidi e coenzimi 325 Citrato sintasi EC Quando lega ossalacetato cambia conformazione, Ci crea il sito per acetil-, L ossalacetato non è più accessibile all acqua. B05 - v gsartor ucleotidi e coenzimi
 che coinvolge is-320.")
164 Citrato sintasi EC ssalacetato Acetil- B05 - v gsartor ucleotidi e coenzimi 327 Citrato sintasi EC La condensazione del ossalacetato con acetil- porta alla formazione di citril- È una catalisi acido base che coinvolge is274 e Asp375. L idrolisi del legame tioestere nel citril- porta alla formazione di citrato e -. La reazione è spontanea a causa dell idrolisi del legame tioestere (-31.5 kj/mol) che coinvolge is-320. is 320 is 274 ssalacetato Asp Citrato is 320 is- 320 is C Asp 375 is 274 Asp 375 B05 - v gsartor ucleotidi e coenzimi
 che coinvolge is-320. B05 - v 1.8.")
165 Citrato sintasi EC La condensazione del ossalacetato con acetil- porta alla formazione di citril- È una catalisi acido base che coinvolge is274 e Asp375. L idrolisi del legame tioestere nel citril- porta alla formazione di citrato e -. La reazione è spontanea a causa dell idrolisi del legame tioestere (-31.5 kj/mol) che coinvolge is-320. B05 - v gsartor ucleotidi e coenzimi 329 Citrato sintasi EC Acetil C 2 C 3 2 Aconitasi FAD 2 uccinato EC La regolazione della citrato sintasi è data dalla FAD deidrogenasi EC disponibilità di substrati, iruvato carbossilasi La concentrazione di ossalacetato è limitante, uccinil sintetasi Aconitasi EC EC AD + + AD + Malato Citrato sintasi deidrogenasi EC EC Fumarasi ssalacetato EC Citrato L ossalacetato è anche substrato della gluconeogenesi, GT α-chetoglutarato deidrogenasi GD + i Isocitrato In mancanza di ossalacetato EC si deidrogenasi accumula Acetil- EC , C 2 La presenza di Acetil- stimola la piruvato AD + AD + + AD + carbossilasi a produrre + AD ossalacetato. + C 3 2 C 2 B05 - v gsartor ucleotidi e coenzimi
166 Aconitasi EC AD + AD + + C 2 FAD 2 FAD 2 uccinato deidrogenasi EC Malato deidrogenasi EC Fumarasi EC uccinil sintetasi EC C 3 Citrato sintasi EC Citrato Aconitasi EC C cis-aconitato Aconitasi EC GT GD + i AD + + α-chetoglutarato deidrogenasi EC C 2 AD + Isocitrato deidrogenasi EC Isocitrato AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 331 Aconitasi EC Catalizza la conversione stereospecifica del 2 citrato in rilasciato dal sito attivo, FAD 2 uccinato el sito FAD attivo deidrogenasi vi è un cluster Fe EC C 3 3 C cis-aconitato GT α-chetoglutarato deidrogenasi Isocitrato GD + i Isocitrato EC deidrogenasi prodotto viene rimosso, il G si EC C 2 avvicina a zero a concentrazioni AD + AD + reali. + AD + + AD + AD + AD + + isocitrato attraverso una Malato deidrogenasi deidratazione e una reidratazione, Fumarasi EC EC L intermedio cis-aconitato non è (Fe 4 4 ) Tre degli ioni Fe sono complessati uccinil da un di tre Cys, sintetasi il quarto è il EC sito che lega il substrato. Il G della reazione è positivo, il Citrato sintasi EC C 2 Citrato Aconitasi EC Aconitasi EC C 2 B05 - v gsartor ucleotidi e coenzimi
167 Aconitasi EC Cys Fe Cys Fe Fe Cys Fe B05 - v gsartor ucleotidi e coenzimi 333 Aconitasi EC Fe Fe Asp- 100 is+ 101 Fe Fe Arg er 642 Citrato Fe Fe Asp- 100 is+ 101 Fe Arg+ 580 Fe 2 3 er 642 (2R,3)-Isocitrato Arg+ 580 Arg+ 580 er 642 er 642 Asp- 100 Asp Rovesciamento di is 101 is 101 Fe Fe Fe Fe Fe Fe cis-aconitato intermedio Fe Fe B05 - v gsartor ucleotidi e coenzimi


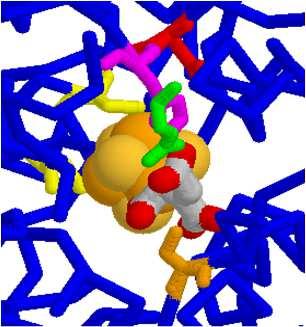
168 Aconitasi EC B05 - v gsartor ucleotidi e coenzimi 335 Aconitasi EC Cys Fe Cys Fe Fe is101 Cys Fe Asp100 Cys Arg580 er642 B05 - v gsartor ucleotidi e coenzimi



169 Aconitasi EC trans-aconitato B05 - v gsartor ucleotidi e coenzimi 337 Aconitasi EC B05 - v gsartor ucleotidi e coenzimi

170 Isocitrato deidrogenasi EC AD + AD + + C 2 2 C 3 3 C Malato deidrogenasi EC Fumarasi EC Citrato sintasi EC FAD 2 FAD uccinato deidrogenasi EC Aconitasi EC uccinil sintetasi EC Aconitasi EC GT GD + i α-chetoglutarato deidrogenasi Isocitrato Isocitrato EC deidrogenasi α-chetoglutarato EC ssalosuccinato C 2 AD + AD + + AD + + AD + C 2 Mg ++ B05 - v gsartor ucleotidi e coenzimi 339 Isocitrato deidrogenasi EC C 2 AD + Catalizza la decarbossilazione + AD ossidativa del isocitrato, + C 3 3 C La porzione nicotinamidica 2 del AD + (o del AD + in un isoenzima) ossida il gruppo Malato a Citrato carbonile, sintasi deidrogenasi EC EC i forma l intermedio Fumarasi ossalosuccinato nel quale vi è EC l interazione con lo ione Mg ++ (Mn ++ ), 2 Il gruppo C - FAD 2 non interessato nella formazione Aconitasi del uccinato EC FAD complesso esce deidrogenasi come C EC , i forma α chetoglutarato. uccinil sintetasi EC Aconitasi EC GT GD + i α-chetoglutarato deidrogenasi Isocitrato Isocitrato EC deidrogenasi α-chetoglutarato EC ssalosuccinato C 2 AD + AD + + AD + + AD + C 2 Mg ++ B05 - v gsartor ucleotidi e coenzimi
 2 Attivata FAD 2 da Ca ++ e AD + (AD antagonista)")

171 Isocitrato deidrogenasi EC C 2 AD + + AD + C 3 C decarbossilazione che quello 3 di ossidazione sono 2 spontanei ( G = kj/mol). Malato Citrato sintasi deidrogenasi EC EC Fumarasi EC La reazione è altamente spontanea, sia il processo di L isocitrato deidrogenasi è regolata: Attivata allostericamente da AD (AT antagonista) 2 Attivata FAD 2 da Ca ++ e AD + (AD antagonista) Aconitasi uccinato EC FAD deidrogenasi Quando il rapporto EC AT/AD è basso il ciclo di Krebs è attivo. uccinil sintetasi EC Aconitasi EC GT GD + i α-chetoglutarato deidrogenasi Isocitrato Isocitrato EC deidrogenasi α-chetoglutarato EC ssalosuccinato C 2 AD + AD + + AD + + AD + C 2 Mg ++ B05 - v gsartor ucleotidi e coenzimi 341 Isocitrato deidrogenasi EC Isocitrato Mg ++ Esce come C 2 AD + C- ossidato a C= Isocitrato B05 - v gsartor ucleotidi e coenzimi
172 α-chetoglutarato deidrogenasi EC AD + AD + + C 2 2 C 3 3 C Malato deidrogenasi EC Fumarasi EC Citrato sintasi EC FAD 2 FAD GT GD + i uccinil sintetasi EC uccinil α-chetoglutarato deidrogenasi EC uccinato deidrogenasi EC AD + + α-chetoglutarato C 2 AD + Isocitrato deidrogenasi EC Aconitasi EC AD + AD + + Aconitasi EC C 2 B05 - v gsartor ucleotidi e coenzimi 343 α-chetoglutarato deidrogenasi EC C 2 AD + È un complesso enzimatico + AD + che converte l α-chetoglutarato a succinil- e C 2. C 3 3 C 2 La reazione produce AD Malatoe conserva Citrato sintasi l energia nel legame deidrogenasi EC tioestere EC Fumarasi EC La reazione è spontanea ( G = kj/mol). 2 Aconitasi FAD 2 uccinato EC FAD deidrogenasi EC E 2 ed E 3 sono conservati nei due complessi enzimatici, Meccanismo quasi identico a quello del complesso piruvato deidrogenasi (E 1, E 2, E 3 ) E 1 ha diversa specificità di substrato. GT GD + i uccinil sintetasi EC uccinil α-chetoglutarato deidrogenasi EC α-chetoglutarato C 2 AD + + AD + C 2 Isocitrato deidrogenasi EC Aconitasi EC AD + AD B05 - v gsartor ucleotidi e coenzimi
173 uccinil sintetasi EC AD + AD + + C 2 2 C 3 3 C Malato deidrogenasi EC Fumarasi EC Citrato sintasi EC FAD 2 FAD GT GD + i uccinato deidrogenasi EC uccinato AD + + C 2 uccinil sintetasi EC uccinil α-chetoglutarato deidrogenasi EC AD + Isocitrato deidrogenasi EC Aconitasi EC AD + AD + + Aconitasi EC C 2 B05 - v gsartor ucleotidi e coenzimi 345 uccinil sintetasi EC AD + AD + + C 2 FAD 2 FAD GT GD + i 2 AD + + C 2 AD + Fumarasi EC uccinato deidrogenasi EC uccinato uccinil sintetasi EC uccinil C 3 3 C Converte il succinil- in Malato succinato Citrato sintasi e produce GT (AT) deidrogenasi EC EC α-chetoglutarato deidrogenasi EC da GD (AD) e i. L enzima è fosforilato in una is. 2 Aconitasi È un eterodimero: EC La subunità α contiene il sito di fosforilazione (is) e lega Aconitasi EC La subunità β dà la specificità Isocitrato per AT o GT. deidrogenasi EC ( G =-2.9 kj/mol). AD +. La reazione è vicina all equilibrio AD + + C 2 B05 - v gsartor ucleotidi e coenzimi
 ubunità α B05 - v 1.")
174 Meccanismo uccinil + + uccinato : + Enzima Enzima GT 2 GD 2 B05 - v gsartor ucleotidi e coenzimi 347 uccinil sintetasi EC ubunità β ito di Fosforilazione (fosfoistidina) ubunità α B05 - v gsartor ucleotidi e coenzimi
175 uccinato deidrogenasi EC AD + AD + + C 2 2 Malato deidrogenasi EC Fumarasi EC C 3 Citrato sintasi EC C FAD 2 FAD Fumarato uccinato deidrogenasi EC uccinato uccinil sintetasi EC Aconitasi EC Aconitasi EC GT GD + i AD + + α-chetoglutarato deidrogenasi EC C 2 AD + Isocitrato deidrogenasi EC AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 349 uccinato deidrogenasi EC er completare il ciclo il succinato deve venire convertito in ossalacetato. Il succinato è deidrogenato a fumarato stereospecificatamente dalla flavoproteina succinato deidrogenasi con produzione di FAD 2. La succinato deidrogenasi è il solo enzima di membrana del ciclo di Krebs. Gli elettroni passano dal succinato al FAD, che è legato covalentemente alla proteina attraverso un residuo di is. Il FAD 2 è riossidato a FAD dal Coenzima Q nella catena di trasporto degli elettroni. Il malonato, strutturalmente analogo al succinato, è un forte inibitore competitivo e blocca il ciclo. uccinato Malonato B05 - v gsartor ucleotidi e coenzimi
176 Fumarasi EC AD + AD + + C 2 2 Fumarasi EC Malato Malato deidrogenasi EC C 3 Citrato sintasi EC C FAD 2 FAD uccinato deidrogenasi EC Fumarato uccinil sintetasi EC Aconitasi EC Aconitasi EC GT GD + i AD + + α-chetoglutarato deidrogenasi EC C 2 AD + Isocitrato deidrogenasi EC AD + AD + + C 2 B05 - v gsartor ucleotidi e coenzimi 351 Fumarasi EC AD + AD + + C 2 2 Fumarasi EC Malato Malato deidrogenasi EC C 3 Citrato sintasi EC C Fumarato 2 Aconitasi FAD 2 uccinato EC FAD deidrogenasi EC La fumarasi catalizza uccinil l idratazione reversibile del sintetasi Aconitasi EC EC fumarato, si forma ()-malato (L-malato). GT α-chetoglutarato È una reazione deidrogenasi stereospecifica, viene idratato il GD + i Isocitrato EC deidrogenasi doppio legame trans ma EC non il cis (maleato). C 2 ella reazione inversa solo il L-malato è AD + AD + + AD + substrato dell enzima + AD + non l isomero D. C 2 B05 - v gsartor ucleotidi e coenzimi
177 Meccanismo - Fumarato + Carbanione C - C + + -Malato Carbocatione B05 - v gsartor ucleotidi e coenzimi 353 Malato deidrogenasi EC AD + + AD + C 3 3 C 2 ssalacetato Citrato sintasi EC Malato Malato deidrogenasi Fumarasi EC EC Aconitasi FAD 2 uccinato EC FAD deidrogenasi EC ossalacetato rigenerando il composto di partenza del La L-malato deidrogenasi ossida il malato a ciclo e producendo AD. uccinil sintetasi Aconitasi EC La reazione è sfavorita ( G = 29.7 kj/mol), 2 ma la EC GT concentrazione di α-chetoglutarato ossalacetato è bassa (<10-6 M), il deidrogenasi GD + i Isocitrato EC che spinge la reazione in avanti. deidrogenasi EC C 2 AD + AD + + AD + + AD + sottrae ulteriormente l ossalacetato dal mezzo. Inoltre la reazione successiva catalizzata dalla citrato sintasi è altamente favorita ( G = kj/mole) e C 2 C 2 B05 - v gsartor ucleotidi e coenzimi
178 40 Energia G ' (kj/mol) Citrato sintasi Aconitasi Isocitrato deidrogenasi α-chetoglutarato deidrogenasi uccinil- sintasi uccinato deidrogenasi Fumarasi Malato deidrogenasi R eazione B05 - v gsartor ucleotidi e coenzimi 355 Energia ITIV G (kj/mol) 0 Citrato sintasi Aconitasi Isocitrato deidrogenasi α-chetoglutarato deidrogenasi uccinil- sintasi uccinato deidrogenasi Fumarasi Malato deidrogenasi EGATIV R eazione B05 - v gsartor ucleotidi e coenzimi
 lo alimentano, le vie sintetiche (anabolismo) ne usano i componenti. È una via AFIBLICA, opera infatti sia nel catabolismo che nell anabolismo cellulare.")
179 Energia B05 - v gsartor ucleotidi e coenzimi 357 Ciclo di Krebs Il ciclo di Krebs è al centro del metabolismo. Le vie degradative (catabolismo) lo alimentano, le vie sintetiche (anabolismo) ne usano i componenti. È una via AFIBLICA, opera infatti sia nel catabolismo che nell anabolismo cellulare. Tutte le reazioni avvengono nella matrice mitocondriale. ei mitocondri vi sono anche gli enzimi della fosforilazione ossidativa e quelli della ossidazione degli acidi grassi e degli aminoacidi. B05 - v gsartor ucleotidi e coenzimi
Metabolismo dei carboidrati I
 Prof. Giorgio Sartor Metabolismo dei carboidrati I Copyright 2001-2017 by Giorgio Sartor. All rights reserved. B09-I Versione 3.0 Feb-17 Indice Glicogeno Metabolismo del Glicogeno C 2 C 2 P 3 -- 3 C Gluconeogensi
Prof. Giorgio Sartor Metabolismo dei carboidrati I Copyright 2001-2017 by Giorgio Sartor. All rights reserved. B09-I Versione 3.0 Feb-17 Indice Glicogeno Metabolismo del Glicogeno C 2 C 2 P 3 -- 3 C Gluconeogensi
Metabolismo dei carboidrati
 rof. Giorgio artor Metabolismo dei carboidrati Copyright 2001-2015 by Giorgio artor. All rights reserved. Versione 1.9.3 apr 2015 Indice Glicogeno Metabolismo del Glicogeno C 2 C 2 3 -- 3 C Gluconeogensi
rof. Giorgio artor Metabolismo dei carboidrati Copyright 2001-2015 by Giorgio artor. All rights reserved. Versione 1.9.3 apr 2015 Indice Glicogeno Metabolismo del Glicogeno C 2 C 2 3 -- 3 C Gluconeogensi
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Nucleotidi e coenzimi
 rof. Giorgio Sartor ucleotidi e coenzimi Copyright 2001-2012 by Giorgio Sartor. All rights reserved. B05 -Versione 1.7 mar 2012 Basi azotate Le basi azotate coinvolte nella formazione di nucleotidi e coenzimi
rof. Giorgio Sartor ucleotidi e coenzimi Copyright 2001-2012 by Giorgio Sartor. All rights reserved. B05 -Versione 1.7 mar 2012 Basi azotate Le basi azotate coinvolte nella formazione di nucleotidi e coenzimi
Carboidrati! Oligosaccaridi Polisaccaridi. B) Carboidrati complessi
 Carboidrati complessi") Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
Utilizzo del glucosio: la glicolisi
 Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Reazione prossima all equilibrio Molto rapida. Trioso-fosfato isomerasi
 5 a reazione Delle 2 molecole prodotte dalla scissione del fruttosio 1,6-bisfosfato solo la gliceraldeide 3-P è il substrato della successiva reazione della glicolisi, perciò il diidrossiacetonefosfato
5 a reazione Delle 2 molecole prodotte dalla scissione del fruttosio 1,6-bisfosfato solo la gliceraldeide 3-P è il substrato della successiva reazione della glicolisi, perciò il diidrossiacetonefosfato
Cap.16 GLICOLISI. Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP H 2 O 2 NAD + 2 NADH + 2H +
 Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato
 2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
AGISCONO NEL SISTEMA NAVETTA 2 ISOFORME DELL ENZIMA: UNA FORMA MITOCONDRIALE ( MEMBRANA MIT. INTERNA) FAD-dipendente
 FAD-dipendente") METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
GLICOLISI ADP ATP. Condizioni aerobiche. Condizioni anaerobiche ATP
 GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
Biosintesi dei carboidrati
 Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
metano alcool metilico formaldeide acido formico CO 2
 -4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
-4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
Catabolismo dei carboidrati Glicolisi
 Catabolismo dei carboidrati Glicolisi Principali vie di utilizzo del Glucosio L estrazione dell energia dai composti organici può essere divisa in tre stadi Metabolismo del glucosio C 6 H 12 O 6 + 6O 2
Catabolismo dei carboidrati Glicolisi Principali vie di utilizzo del Glucosio L estrazione dell energia dai composti organici può essere divisa in tre stadi Metabolismo del glucosio C 6 H 12 O 6 + 6O 2
1. L'enzima che catalizza la trasformazione della 3-fosfogliceraldeide in acido 1,3 difosfoglicerico è una:
 1. L'enzima che catalizza la trasformazione della 3-fosfogliceraldeide in acido 1,3 difosfoglicerico è una: a) idrolasi b) isomerasi c) aldolasi d) fosfatasi e) deidrogenasi 2. Quale dei sottoelencati
1. L'enzima che catalizza la trasformazione della 3-fosfogliceraldeide in acido 1,3 difosfoglicerico è una: a) idrolasi b) isomerasi c) aldolasi d) fosfatasi e) deidrogenasi 2. Quale dei sottoelencati
La quantità di glu presente nei liquidi extracellulari è di circa 20g, 80kcal.
 I principali nutrienti metabolici sono il glucosio e gli acidi grassi. In circostanze normali il glucosio è l unico nutriente che possa essere utilizzato dal cervello. E anche utilizzato preferenzialmente
I principali nutrienti metabolici sono il glucosio e gli acidi grassi. In circostanze normali il glucosio è l unico nutriente che possa essere utilizzato dal cervello. E anche utilizzato preferenzialmente
Lattato. Glicolisi. Piruvato
 Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
GLUCOSIO. Vena porta DIGESTIONE DEI DISACCARIDI. Enzimi parete intestinale
 DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
Lezione 5. Il metabolismo dei carboidrati
 Lezione 5 Il metabolismo dei carboidrati 1 La produzione di energia: catabolismo dei nutrienti La degradazione enzimatica delle sostanze nutritive principali (carboidrati, lipidi e proteine) procede attraverso
Lezione 5 Il metabolismo dei carboidrati 1 La produzione di energia: catabolismo dei nutrienti La degradazione enzimatica delle sostanze nutritive principali (carboidrati, lipidi e proteine) procede attraverso
Metabolismo dei carboidrati II
 rof. Giorgio Sartor Metabolismo dei carboidrati II Gluconeogenesi Glicogeno Via dei pentosi fosfati 1 Copyright 2001-2019 by Giorgio Sartor. All rights reserved. Versione 3.3.1 Mar-19 Indice Glicogeno
rof. Giorgio Sartor Metabolismo dei carboidrati II Gluconeogenesi Glicogeno Via dei pentosi fosfati 1 Copyright 2001-2019 by Giorgio Sartor. All rights reserved. Versione 3.3.1 Mar-19 Indice Glicogeno
Aerobiosi C A T A B O L I S M O. Lez 4A. Schema generale del metabolismo dei glucidi
 Schema generale del metabolismo dei glucidi -1- Aerobiosi In condizioni aerobiche il piruvato prodotto dalla glicolisi e dalla degradazione di alcuni aminoacidi è ossidato a H 2 O e CO 2 nella respirazione
Schema generale del metabolismo dei glucidi -1- Aerobiosi In condizioni aerobiche il piruvato prodotto dalla glicolisi e dalla degradazione di alcuni aminoacidi è ossidato a H 2 O e CO 2 nella respirazione
Modulo 14 Il ciclo di Krebs
 Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
Modulo 13 La glicolisi
 1 Modulo 13 La glicolisi La glicolisi: via metabolica centrale del catabolismo del glucosio Via metabolica antica avviene interamente nel citoplasma delle cellule. onverte il glucosio in piruvato, producendo
1 Modulo 13 La glicolisi La glicolisi: via metabolica centrale del catabolismo del glucosio Via metabolica antica avviene interamente nel citoplasma delle cellule. onverte il glucosio in piruvato, producendo
Metabolismo del glucosio
 Metabolismo del glucosio Metabolismo del glucosio La glicolisi avviene in tutte le cellule umane Percorsi metabolici del glucosio in varie cellule: è metabolizzato in modo diverso nelle varie cellule TABELLA
Metabolismo del glucosio Metabolismo del glucosio La glicolisi avviene in tutte le cellule umane Percorsi metabolici del glucosio in varie cellule: è metabolizzato in modo diverso nelle varie cellule TABELLA
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
Prof. Giorgio Sartor. Invertebrati marini. I nutrienti. Fonti di carbonio ed energia. Fonte di carbonio. Fonte di energia. Composti organici.
 Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
Via finale comune per l ossidazione di tutti i combustibili metabolici che entrano nella via come molecola di acetil CoA
 Via finale comune per l ossidazione di tutti i combustibili metabolici che entrano nella via come molecola di acetil CoA C 6 H 12 O 6 + 6O 2 6CO 2 + 6H 2 O Processi molecolari in cui è coinvolto il consumo
Via finale comune per l ossidazione di tutti i combustibili metabolici che entrano nella via come molecola di acetil CoA C 6 H 12 O 6 + 6O 2 6CO 2 + 6H 2 O Processi molecolari in cui è coinvolto il consumo
Gluconeogenesi. Sintesi de-novo di glucosio
 Gluconeogenesi Sintesi de-novo di glucosio La gluconeogenesi garantisce il mantenimento dei livelli di glucosio nel sangue anche per molto tempo dopo l assorbimento e la completa ossidazione del glucosio
Gluconeogenesi Sintesi de-novo di glucosio La gluconeogenesi garantisce il mantenimento dei livelli di glucosio nel sangue anche per molto tempo dopo l assorbimento e la completa ossidazione del glucosio
Il metabolismo cellulare
 Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Variazioni di energia libera nella glicolisi
 Variazioni di energia libera nella glicolisi 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio
Variazioni di energia libera nella glicolisi 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio
Fosforilazione a livello del substrato
 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
Fosforilazione a livello del substrato
 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
La via glicolitica. Caratteristiche generali Le reazioni della via glicolitica. Francesca Anna Scaramuzzo, PhD
 Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche La via glicolitica Caratteristiche generali Le reazioni della via glicolitica Francesca
Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche La via glicolitica Caratteristiche generali Le reazioni della via glicolitica Francesca
catabolismo anabolismo
 Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
Capitolo B2 Il metabolismo energetico: dal glucosio all ATP
 Capitolo B2 Il metabolismo energetico: dal glucosio all ATP Quesiti e problemi 1 Le funzioni del metabolismo cellulare sono: ricavare energia utile per la cellula attraverso la degradazione di sostanze
Capitolo B2 Il metabolismo energetico: dal glucosio all ATP Quesiti e problemi 1 Le funzioni del metabolismo cellulare sono: ricavare energia utile per la cellula attraverso la degradazione di sostanze
Cap.19. Ciclo di Krebs. o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico
 Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Una panoramica del ciclo dell acido ciclico
 Ciclo di Krebs Una panoramica del ciclo dell acido ciclico Concetti chiave Il ciclo dell acido citrico, o ciclo di Krebs, o ciclo degli acidi tricarbossilici (TCA), è un processo catalitico a 8 tappe che
Ciclo di Krebs Una panoramica del ciclo dell acido ciclico Concetti chiave Il ciclo dell acido citrico, o ciclo di Krebs, o ciclo degli acidi tricarbossilici (TCA), è un processo catalitico a 8 tappe che
a-amilasi salivare e pancreatica destrine lattasi maltasi oligosaccaridasi saccarasi superficie luminale degli enterociti
 Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
INTRODUZIONE AL METABOLISMO. dal gr. metabolè = trasformazione
 INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
Modulo 12 : Il metabolismo: concetti di base
 Modulo 12 : Il metabolismo: concetti di base Il metabolismo Il metabolismo è l insieme di tutte le trasformazioni chimiche che avvengono nelle cellule (ed in ultima analisi negli organismi) per adempiere
Modulo 12 : Il metabolismo: concetti di base Il metabolismo Il metabolismo è l insieme di tutte le trasformazioni chimiche che avvengono nelle cellule (ed in ultima analisi negli organismi) per adempiere
Introduzione allo studio del metabolismo Parte I
 Introduzione allo studio del metabolismo Parte I FOTOTROFI Le piante ed alcuni tipi di batteri ricavano l energia libera dal Sole mediante la fotosintesi, un processo in cui l energia luminosa è convertita
Introduzione allo studio del metabolismo Parte I FOTOTROFI Le piante ed alcuni tipi di batteri ricavano l energia libera dal Sole mediante la fotosintesi, un processo in cui l energia luminosa è convertita
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
METABOLISMO OSSIDATIVO
 METABOLISMO OSSIDATIVO IDROLISI DEI LIPIDI LIPASI Fosfolipidi Fosfolipasi β ossidazione degli acidi grassi NEL CITOPLASMA: attivazione dell acido grasso Consumo di 2 ATP ΔG idrolisi ATP = -35,9 kj mol
METABOLISMO OSSIDATIVO IDROLISI DEI LIPIDI LIPASI Fosfolipidi Fosfolipasi β ossidazione degli acidi grassi NEL CITOPLASMA: attivazione dell acido grasso Consumo di 2 ATP ΔG idrolisi ATP = -35,9 kj mol
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
2 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA
 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI
 METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
GLICOLISI ADP ATP. Condizioni aerobiche. Condizioni anaerobiche ATP
 GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
6 H 2. con G=-686 kcal/mole di H 12 O 6 O + 6 CO O 2. glucosio La respirazione avviene in tre stadi principali; ognuno di questi
 La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
Indice generale 1. Introduzione alla biochimica 2. Acqua, ph, acidi, basi, sali, tamponi
 Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
Indice generale 1. Introduzione alla biochimica 1 Unità di misura 1 Energia e calore 3 Struttura dell atomo 3 Tavola periodica degli elementi 4 Legami chimici 8 Legami covalenti 8 Legami non covalenti
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia.
 La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
Energia e metabolismi energetici
 Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
Il metabolismo ossidativo
 Il metabolismo ossidativo 1 13.3.2010 Ore 14:00 2 Riposare stanca! Respirare (muscoli, cuore) Pensare/sognare (cervello, cuore) Digerire (apparato gastroenterico, cuore, reni) Controllo termico (cuore,
Il metabolismo ossidativo 1 13.3.2010 Ore 14:00 2 Riposare stanca! Respirare (muscoli, cuore) Pensare/sognare (cervello, cuore) Digerire (apparato gastroenterico, cuore, reni) Controllo termico (cuore,
OSSIDATIVA ASSOCIATA ALLA CATENA DI TRASPORTO ELETTRONICO
 LA RESPIRAZIONE Il processo respiratorio consiste essenzialmente in una serie di reazioni ossidative finalizzate a demolire le molecole organiche (zuccheri, ma anche acidi grassi) per recuperare parte
LA RESPIRAZIONE Il processo respiratorio consiste essenzialmente in una serie di reazioni ossidative finalizzate a demolire le molecole organiche (zuccheri, ma anche acidi grassi) per recuperare parte
INTRODUZIONE AL METABOLISMO
 INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA
 Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
BIOCHIMICA e BIOTECNOLOGIE degli ALIMENTI
 Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2016-17 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2016-17 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
ENERGIA LIBERA DI GIBBS (G)
") METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
NADH e NADPH. ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente)
 della cellula NADH, NADPH = 2 moneta (potere riducente)") NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
IL GLUCOSIO. Il glucosio può provenire dalla dieta, in quanto tale, o derivare dalla demolizione di carboidrati complessi.
 IL GLUCSI Il glucosio può provenire dalla dieta, in quanto tale, o derivare dalla demolizione di carboidrati complessi. Viene metabolizzato per via aerobica o anaerobica, fornendo energia all organismo.
IL GLUCSI Il glucosio può provenire dalla dieta, in quanto tale, o derivare dalla demolizione di carboidrati complessi. Viene metabolizzato per via aerobica o anaerobica, fornendo energia all organismo.
Il metabolismo: concetti di base
 Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche Il metabolismo: concetti di base Caratteristiche generali Strategie Trofiche Il metabolismo:
Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche Il metabolismo: concetti di base Caratteristiche generali Strategie Trofiche Il metabolismo:
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO)
") RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
IL METABOLISMO Somma delle reazioni che convertono sostanze nutrienti in fonti di ENERGIA o PRODOTTI COMPLESSI.
 IL METABOLISMO Somma delle reazioni che convertono sostanze nutrienti in fonti di ENERGIA o PRODOTTI COMPLESSI. reazioni enzimatiche organizzate in vie discrete H recettore procedono attraverso diversi
IL METABOLISMO Somma delle reazioni che convertono sostanze nutrienti in fonti di ENERGIA o PRODOTTI COMPLESSI. reazioni enzimatiche organizzate in vie discrete H recettore procedono attraverso diversi
Laboratorio Fitness & Wellness
 Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE
 BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA
 I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
Capitolo 6 La respirazione cellulare
 Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
glucosio piruvato Ac-CoA
 glucosio piruvato Ac-CoA Biochimica FA040 NADH CTF 2011 Gluconeogenesi e metabolismo del glicogeno fegato & muscolo GLICGEN TRIGLICERIDI Ac-Coa Gluconeogenesi Via metabolica per la sintesi di glucosio
glucosio piruvato Ac-CoA Biochimica FA040 NADH CTF 2011 Gluconeogenesi e metabolismo del glicogeno fegato & muscolo GLICGEN TRIGLICERIDI Ac-Coa Gluconeogenesi Via metabolica per la sintesi di glucosio
Le reazioni esoergoniche e quelle endoergoniche del metabolismo cellulare sono legate dalla molecola di ATP.
 1 METABOLISMO CELLULARE Definizione Metabolismo cellulare insieme delle reazioni che producono e necessitano energia nel nostro corpo. Nel metabolismo si distinguono due fasi: Fase catabolica Insieme delle
1 METABOLISMO CELLULARE Definizione Metabolismo cellulare insieme delle reazioni che producono e necessitano energia nel nostro corpo. Nel metabolismo si distinguono due fasi: Fase catabolica Insieme delle
Disponibilità di trasportatori del glucosio ESOCHINASI
 ESOCHINASI Disponibilità di trasportatori del glucosio Le esochinasi sono attivate dall elevato flusso di glucosio in direzione intracellulare (la loro attività è aumentata dalla concentrazione del substrato
ESOCHINASI Disponibilità di trasportatori del glucosio Le esochinasi sono attivate dall elevato flusso di glucosio in direzione intracellulare (la loro attività è aumentata dalla concentrazione del substrato
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico
 Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
IL METABOLISMO ENERGETICO
 IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI
 GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
Esercitazione Esempi di domande del 2 test
 Esercitazione Esempi di domande del 2 test Esempio 1. Gli enzimi opposti glicogeno fosforilasi e glicogeno sintasi sono regolati in maniera opposta da una serie di effettori. Quali? Come avviene la regolazione
Esercitazione Esempi di domande del 2 test Esempio 1. Gli enzimi opposti glicogeno fosforilasi e glicogeno sintasi sono regolati in maniera opposta da una serie di effettori. Quali? Come avviene la regolazione
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
prodotto finale della glicolisi
 Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
IL METABOLISMO. Dal carbonio agli OGM Capitolo 2
 IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
Biologia. La cellula al lavoro
 Biologia La cellula al lavoro Capitolo 5 La cellula al lavoro 1. Il metabolismo cellulare: come le cellule ricavano energia 2. La glicolisi è la prima fase della demolizione del glucosio 3. La respirazione
Biologia La cellula al lavoro Capitolo 5 La cellula al lavoro 1. Il metabolismo cellulare: come le cellule ricavano energia 2. La glicolisi è la prima fase della demolizione del glucosio 3. La respirazione
Metabolismo del Glucosio
 Metabolismo del Glucosio Schema generale del metabolismo del glucosio Una panoramica della glicolisi Conce2 chiave La glicolisi consiste nella degradazione del glucosio a piruvato e utilizza, al contempo,
Metabolismo del Glucosio Schema generale del metabolismo del glucosio Una panoramica della glicolisi Conce2 chiave La glicolisi consiste nella degradazione del glucosio a piruvato e utilizza, al contempo,
La fotosintesi e la respirazione cellulare
 La fotosintesi e la respirazione cellulare 1 La fotosintesi immagazzina energia e rilascia ossigeno L ATP è una molecola antica e universale, sintetizzata da tutti gli organismi viventi. L ATP viene utilizzata
La fotosintesi e la respirazione cellulare 1 La fotosintesi immagazzina energia e rilascia ossigeno L ATP è una molecola antica e universale, sintetizzata da tutti gli organismi viventi. L ATP viene utilizzata
Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre
 Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre molecole, anche chimicamente simili al substrato. Saturabilità:
Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre molecole, anche chimicamente simili al substrato. Saturabilità:
Come le cellule traggono energia dal cibo: produzione di ATP
 Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Metabolismo glucidico
 Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
Metabolismo glucidico Digestione degli zuccheri a-amilasi salivare e pancreatica maltotrioso destrina maltosio glucosio lattosio amido saccarosio maltosio maltotrioso destrine lattasi maltasi oligosaccaridasi
scaricato da www.sunhope.it
 CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO