I CANALI IONICI, I RECETTORI IONOTROPI ed i RECETTORI METABOTROPI LEZIONE NR PSICOBIOLOGIA
|
|
|
- Miranda Moretti
- 6 anni fa
- Visualizzazioni
Transcript

1 I CANALI IONICI, I RECETTORI IONOTROPI ed i RECETTORI METABOTROPI LEZIONE NR PSICOBIOLOGIA
.")
2 I CANALI IONICI I canali ionici sono sistemi proteici che attraversano l intero spessore della membrana plasmatica e mediano il trasporto PASSIVO di ioni liberi mediante diffusione facilitata. Possono essere costituiti da una singola proteina o da un complesso multi-proteico che comprende diverse sub-unità (ognuna indicata con una lettera greca). Le varie sub-unità formano un complesso proteico che interagisce direttamente con i lipidi di membrana ed ha al suo interno un poro acquoso che aprendosi su entrambi i versanti della cellula costituisce un passaggio o gate.

 sono coinvolti nella generazione di potenziali d azione (attraverso il passaggio di centinaia di milioni di ioni al sec.")
3 I CANALI IONICI Trattandosi di trasporto passivo, i canali non possono influenzare la direzione del movimento che avverrà sempre secondo gradiente di concentrazione, ma ne possono regolare il flusso, a seconda che il canale sia aperto o chiuso. Tramite queste regolazioni i canali ionici possono modificare la differenza di cariche elettriche intra-ed extracellulari, che di norma costituiscono il potenziale di membrana a riposo. Nelle cellule nervose (o in generale in cellule eccitabili) sono coinvolti nella generazione di potenziali d azione (attraverso il passaggio di centinaia di milioni di ioni al sec.) L importanza dei canali ionici è infine dimostrata dalle patologie che derivano dal loro malfunzionamento (canalopatie) che può avvenire per cause genetiche (fibrosi cistica, epilessia idiopatica, atassia spinocerebellare etc) o per cause tossiche (veleni o farmaci)


4 I CANALI IONICI I canali ionici come molti altri composti proteici possono esistere con una grande varietà di isoforme, cioè di catene polipeptidiche alternative, che vengono originate da modificazioni posttraduzionali o attraverso l editing degli mrna. Questo conferisce caratteristiche strutturali e funzionali altamente specifiche ad un canale e/o alle sue isoforme, consentendo alla cellula di ottenere funzioni specializzate per i vari composti molecolari.

5 I CANALI IONICI Un importante caratteristica dei canali ionici è la loro selettività per un dato ione o un dato tipo di ione (anioni, cationi). Tale proprietà dipende dalle carattiristiche strutturali ed elettrostatiche del poro acquoso. Infatti l imboccatura del canale è di norma dotata di un filtro (o anello) di selettività, composto da amminoacidi con carica elettrica positiva o negativa. Altri fattori di selettività dipendono dalla dimensione e dalla struttura del poro acquoso, nonché nel meccanismo molecolare di trasferimento dello ione attraverso il poro (Ioni spogliati del guscio di idratazione)
, i canali sono di norma in modalità chiusa e si aprono solo in presenza di uno stimolo")
6 I CANALI IONICI Un altra caratteristica molto importante dei canali ionici riguarda la regolazione della loro apertura. Infatti, a parte alcuni canali che sono costantemente aperti (resting o leak, rispettivamente del Cl- e del K+), i canali sono di norma in modalità chiusa e si aprono solo in presenza di uno stimolo appropriato, rimanendo aperti per pochi msec. Subito dopo l apertura, molti canali ionici vanno incontro ad un altra modificazione conformazionale che consiste nell occlusione del poro acquoso e nella contemporanea incapacità di aprirsi nuovamente, anche qualora si ripresentasse lo stimolo che ne determina l apertura. Questo stato indicato come inattivo o refrattario viene mantenuto di norma per alcuni msec, dopodiché il canale ionico torna in modalità chiuso.


7 I CANALI IONICI Poiché canali dedicati al passaggio dello stesso ione possono avere organizzazioni strutturali diverse, si preferisce di norma classificare i canali ionici in base alla modalità di regolazione della loro apertura. RECETTORI IONOTROPI Seguendo questi criteri i canali vengono raggruppati in famiglie di canali relativamente omogenei fra loro sia dal punto di vista funzionale che strutturale. Possiamo avere: Canali regolati dal potenziale di membrana Canali regolati da ligando Canali regolati da altre modalità (tensione meccanica, luce, etc )

8 Canali regolati dal potenziale di membrana Vengono anche denominati come canali voltaggio-dipendenti o canali regolati da voltaggio. Conferiscono alle cellule eccitabili la capacità di variare in pochi msec il contenuto intracellulare di cariche elettriche rispetto all esterno. I canali voltaggio-dipendente sono così denominati perché si aprono solo quando il potenziale di membrana della cellula raggiunge un determinato valore, chiamato valore soglia.
 che si dispongono in maniera simmetrica così da formare il tetramero che compone il")
9 Canali regolati dal potenziale di membrana Queste capacità dipendono dalla struttura del canale, che comprende 4 domini di circa amminoacidi. Ciascun dominio comprende 6 diversi segmenti transmembrana (da S1 ad S6 in struttura secondaria di alphaeliche) che si dispongono in maniera simmetrica così da formare il tetramero che compone il canale. Dei 6 segmenti, s5 ed s6 compongono l apertura del poro acquoso e fra loro c è un ansa di circa 20 amminoacidi che insieme alle altre 3 anse, forma il filtro di selettività. I segmenti da s1 ad s4 sono formati da amminoacidi carichi elettricamente e formano il sensore di voltaggio. Alcuni canali possono infine avere anche altre sub-unità accessorie che mediano fenomeni di grande importanza per il canale come la sua corretta posizione sulla membrana plasmatica
10 Canali regolati dal potenziale di membrana Questa tipologia di canali ha un alto livello di selettività per il Na+, il K+, il Ca+, Cl- e H+. Ed ogni canale specifico per questi ioni può presentare diverse isoforme o sotto-tipologie, per via del gran numero di geni differenti che codificano per questi canali. Ad esempio: Na+ = 10 geni K+ = >100 geni Ca+ = 16 geni
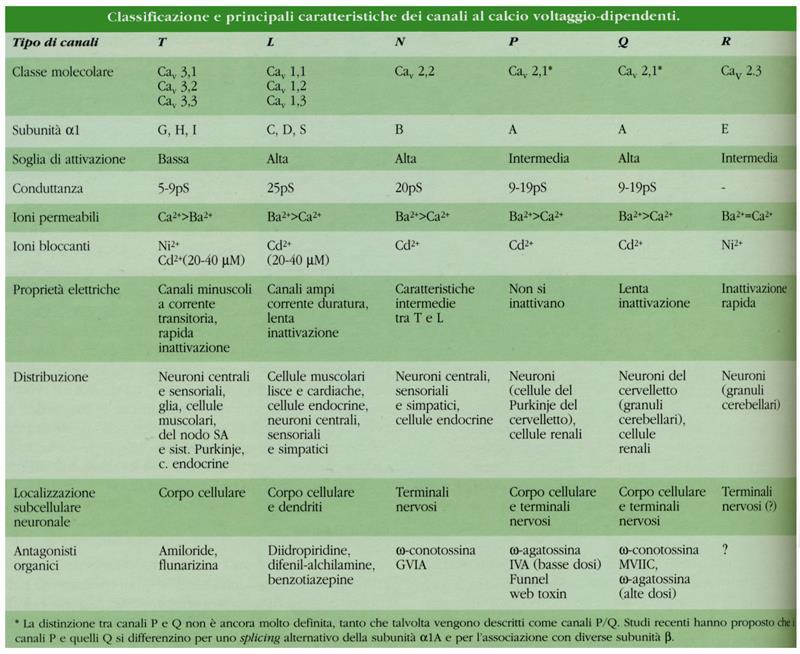
11 Canali voltaggio-dipendenti del calcio

12 Canali voltaggio-dipendenti del sodio
13 Canali regolati da ligando RECETTORI IONOTROPI Sono complessi proteici di membrana, caratterizzati dalla presenza sulla loro superficie non solo del poro acquoso, ma anche di una regione specifica, detta sito recettoriale (sito di legame), corrispondente ad una sorta di tasca che accoglie in maniera stereo-specifica una molecola di segnalazione, generalmente indicata come ligando. Si contrappongono ai recettori metabotropi che invece indicano una categoria di recettori, privi di poro acquoso e che agiscono tramite una cascata di eventi, sui recettori ionotropi.

14 RECETTORI IONOTROPI Alcuni di questi recettori hanno il sito di legame sul versante extracellulare quindi il ligando deve arrivare dall esterno, in altri invece quest ultimo è posto all interno della cellula, ed il ligando viene prodotto nel citoplasma. L incontro casuale fra canale e ligando porta alla formazione del complesso recettore-ligando, ed è quindi, del tutto analogo alla formazione del complesso enzima substrato. In entrambi i casi infatti, il legame avviene grazie a forze deboli, e porta ad un cambiamento conformazionale dell intero sistema proteico ed in questo all apertura del canale. Vi sono inoltre anche altre importanti analogie: La formazione del complesso sito di legame-ligando avviene in maniera del tutto casuale e quindi dipendente dalla concentrazione del ligando nello spazio extra-/intracellulare. Questo fenomeno è di grande importanza nelle sinapsi chimiche.
15 RECETTORI IONOTROPI In molti casi, il sito di legame può ospitare molecole diverse dal ligando. Queste sostanze possono avere origine endogena (molecole di regolazione) o esogena (farmaci). E possono essere ulteriormente classificati in: Agonisti Antagonisti competitivi Fig Antagonisi non-competitivi Una volta formatosi, il complesso recettoreligando è reversibile, con il ligando che può essere nuovamente rilasciato nell ambiente o degradato
16 RECETTORI IONOTROPI Alcuni recettori sono provvisti di siti di legame addizionali che riconoscono molecole diverse dal ligando. In analogia con il fenomeno della regolazione allosterica enzimatica, anche in questo caso il sito è definito allosterico e può essere occupato da modulatori allosterici, classificati come: Attivatore allosterico Fig Inibitore allosterico Anche in questi casi, l origine dei modulatori allosterici, può essere endogena o esogena.
17 RECETTORI IONOTROPI Anche per i recettori ionotropi, vale la regola generale dei canali ionici circa il tempo di refrattarietà fra un apertura e la successiva. Fig. 5.16
18 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca2+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )


19 Recettori ionotropi della superfamiglia dei recettori nicotinici Sono tutti costituiti da 5 sub-unità principali, aventi entrambi i terminali nell ambiente extracellulare. Per questo motivo vengono anche denominati recettori pentamerici. Queste sub-unità possono essere uguali o diverse fra loro e combinarsi in modo vario a seconda del recettore in questione. Ogni sub-unità contiene 4 regioni ad alpha-elica transmembrana (M1-M4), connesse fra loro da brevi anse che sulla regione M3 creano il poro acquoso. Inoltre sul versante extra-cellulare, questi recettori presentano uno o più siti di legame per i neurotrasmettitori e/o per i modulatori allosterici. Per quanto concerne la tipologia di ioni trasportati, il loro livello di selettività è relativamente basso e trasportano quindi cationi (Na+, K+ e Ca+) oppure anioni (Cl-) I recettori ionotropi nicotinici presentano una grande varietà di isoforme e quindi un numero molto elevato di sottotipi recettoriali.





20 Recettori ionotropi della superfamiglia dei recettori nicotinici Recettori nicotinici Acetilcolina Serotonina GABA Glicina Eccitatori - depolarizzazione Inibitori - iperpolarizzazione

21 Recettore nicotinico dell Acetilcolina I recettori nicotinici dell Acetilcolina (ACh), prendono il loro nome per via del loro potente agonista nicotina. Essendo stati i primi recettori ionotropi ad essere studiati e classificati essi fungono da paradigma per tutti i recettori ionotropi pentamerici. Fig. 5.17


22 Recettore nicotinico dell Acetilcolina La struttura e la funzionalità dei recettori per l Ach sono diverse a seconda del tessuto in cui essi sono espressi. È possibile infatti avere: Recettori muscolari (con subunità apha, beta, gamma, delta ed epsilon) Recettori neuronali (con subunità apha e beta)
23 Recettore nicotinico della serotonina Il recettore 5-HT3 è un recettore-canale attivato dal ligando serotonina (5-idrossitriptamina) che permette il flusso di Na + e K + ; ha una struttura di membrana simile a quella del recettore colinergico nicotinico, con 5 sub-unità chiamate 5-HT3a-e. La subunità 5-HT3a è portatrice del sito di legame con la serotonina ed è quindi presente in ogni recettore in combinazione con le altre subunità. Il legame della serotonina su due siti recettoriali determina l apertura di un canale permeabile ai cationi Na +, K + e Ca ++ con conseguente depolarizzazione desensibilizzante. Fig Questi recettori sono localizzati sulle terminazioni parasimpatiche nel tratto gastrointestinale e anche nelle afferenze vagali e splancniche. Nel sistema nervoso centrale (SNC), invece, vi è una elevata densità di recettori 5-HT3 nel nucleo del tratto solitario e nell area postrema (dove c è il centro del vomito), ma anche nel nucleus accumbens, amigdala, ippocampo, corteccia entorinale e frontale) I recettori 5-HT3 presenti nel canale gastroenterico e nel SNC sono coinvolti nella risposta emetica e costituiscono la base anatomica per le proprietà antiemetiche degli antagonisti recettoriali 5-HT3.
24 Recettore nicotinico del GABA Il GABA-A è il recettore per l acido gammaamminobutirrico (GABA) che è il più importante neurotrasmettitore inibitorio del cervello. Il GABA è agonista endogeno del recettore e si lega sul sito di legame, mediando una modificazione allosterica che fa aprire il canale per gli anioni, principalmente Cl-. Fig Il canale è formato da diverse subunità, leggermente diverse a seconda del distretto nervoso in cui si trova: 2 subunità α 2 subunità β 1 subunità γ
25 Recettore nicotinico del GABA Esistono molti agonisti ed antagonisti del recettore GABA A, che si legano alle diverse subunità su diversi siti di legame specifici, a loro dedicati, i più importanti comprendono: - Benzodiazepine (ansiolitici) - Barbiturici (sedativi, ipnotici) - Steroidi (ormoni derivanti dal cortisolo) - Etanolo (effetto simil-ansiolitico) - Picrotossina (antagonista) Fig e 5.21
26 Recettore nicotinico del GABA Anche nel caso del GABA-A, le varie subunità possono presentare diverse isoforme. In particolare abbiamo 6 isoforme per l unità alpha, 4 isoforme per la beta, e 4 isoforme per la gamma. Tab. 5.2 Queste isoforme si combinano fra loro in modo vario dando origine ad un ampio repertorio di sottotipi recettoriali espressi in modo specifico nei vari distretti del SNC


 e 2 beta.")
27 Recettore nicotinico della glicina Il recettore per la glicina (GlyR) è un recettore simile a quello del Gaba, infatti gli stessi agonisti e antagonisti agiscono anche su questo tipo di recettore. Tuttavia, nel cervello questo recettore è presente in quantità di gran lunga inferiore rispetto al GABA-A., con una distribuzione limitata al tronco dell encefalo ed al midollo spinale ed in aggiunta, nella retina. In fase embrionale il recettore della glicina è composto da 5 subunità alfa. È composto da 3 subunità alfa (4 isoforme) e 2 beta. Le 5 subunità recettoriali sono assemblate a formare una canale centrale permeabile allo ione Cl-. Il loro malfunzionamento provoca una malattia chiamata come iperekplessia (eccessiva reazione d allarme).
28 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca2+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )
, sono fra loro diversi sia per la struttura che per la funzione.")
29 Recettori ionotropi del glutammato Questa classe di recettori comprende 3 diversi tipi, chiamati AMPA, Kainato ed NMDA, che pur essendo attivati dallo stesso neurotrasmettitore amminoacido glutammato (acido glutammico), sono fra loro diversi sia per la struttura che per la funzione. I loro nomi dipendono dagli agonisti specifici che ne permisero l identificazione: AMPA = acido alpha-amino-3-idrossi-5-metil-4-isoxa-zolpropionico Kainato = acido Kainico NMDA = N-metil-D-aspartato La loro importanza deriva dal fatto che il glutammato è il neurotrasmettitore in assoluto più presente nel cervello e costituisce il principale agente della neurotrasmissione eccitatoria. Questi recettori mediano importanti funzioni come la plasticità sinaptica (apprendimento e memoria), ma se stimolati eccessivamente (ictus, epilessia) possono dare il via a processi di neurotossicità che porta alla morte cellulare. Per tale motivo è possibile che questi recettori abbiano un ruolo alla base delle malattie neurodegenerative.

30 Recettori ionotropi del glutammato Per quanto concerne la specificità, i pori acquosi dei recettori AMPA e Kainato hanno una minore specificità e permettono il passaggio sia di K+ che di Na+, ed in quantità minore di Ca2+. Di contro il recettore NMDA ha una spiccata specificità per il Ca2+ e molto più bassa per gli altri cationi. NMDA AMPA, Kainato

31 Recettori ionotropi del glutammato Dal punto di vista strutturale, questi recettori sono composti da 4 o 5 sub-unità principali, caratterizzate da una comune organizzazione di base della catena polipeptidica, con l estremità N-terminale in ambiente extracellulare e l estremità C-terminale in ambiente intra-cellulare. Ogni subunità contiene sempre 3 regioni trans-membrana (M1, M3 ed M4) ed un ansa (M2) situata nel versante intracellulare il cui scopo è controllare la permeabilità del poro acquoso. Fig. 5.22
32 Recettori ionotropi del glutammato La specificità e l importanza di questi recettori è indirettamente confermata dalla molteplicità dei tipi di subunità che li compongono. Queste sub-unità possono essere divise in gruppi sulla base delle somiglianze nelle loro sequenze amminoacidiche Tab. 5.3
33 Recettori ionotropi del glutammato Recettori del glutammato AMPA Kainato NMDA GluR1 GluR3 GluR3 GluR4 KA1, 2 GluR5 GluR6 GluR7 NR1 NR2A,B C,D NR3A,B Eccitatori - depolarizzazione
34 AMPA Gli AMPA sono i recettori ionotropi più presenti nel nostro cervello ed i principali mediatori della trasmissione eccitatoria veloce. Sono composti da 4 sub-unità (GluR1-4), di cui 2 sempre di tipo GluR1. Ciascun tipo di sub-unità si presenta con diverse varianti e sottotipi che hanno origine da modificazioni post-trascrizionali dell RNA, come: Q= glutammina permeabile a tutti i cationi R= arginina permeabile a Na e K, ma non al Ca Fig Splicing alternativo Editing RNA Oppure da modificazioni post-traduzionali della catena polipeptidica, come la fosforilazione.

35 AMPA A fronte di soli 4 geni che codificano per i 4 tipi di base di sub-unità AMPA, le molecole di RNA vanno incontro a varie modificazioni da cui consegue la produzione di una vasto numero di tipi e sottotipi recettoriali, ciascuno dei quali ha caratteristiche funzionali proprie e può essere espresso in modo differenziale nei vari distretti cerebrali. Al fine di studiare questa vasta possibilità recettoriale, vengono utilizzate coltivazioni in vitro oppure modelli animali nei quali i geni che codificano per gli AMPA vengono selettivamente mutati. Attraverso questi studi si è potuto dimostrare che gli antagonisti AMPA riescono a prevenire la morte cellulare a seguito di un ictus, inibendo l eccessiva stimolazione degli AMPA. In conclusione, gli AMPA svolgono un ruolo fondamentale di stimolazione dell attività elettrica e di regolazione indiretta dei processi di morte neuronale.

36 Kainato Il Kainato (acido kainico) è un agente convulsivante, che oltre ad essere un agonista dei recettori AMPA, riesce ad attivare dei canali che hanno come agonista selettivo questa molecola, denominati appunto canali del kainato. Questi canali sono generalmente formati da un complesso tetramerico di 5 possibili sub-unità KA 1-2 e iglur 5-7 che si combinano tra loro. Queste subunità possono variamente combinarsi tra di loro ma è stabile la presenza di GluR 5 o GluR 6 per ottenere un recettore che, attivato dall agonista selettivo, sia in grado di generare una corrente cationica intensa. Fig Entrambe queste due subunità possono esistere in due varianti diverse, cioè nella sequenza di aminoacido della parete del poro-canale si trova sia la variabile con arginina che rende il recettore poco permeabile al calcio, sia residui di glutammina che lo rendono altamente permeabile a sodio e calcio.


37 Kainato Meno abbondanti dei recettori AMPA nel SNC, questi recettori sono prevalentemente espressi nello striato, nei nuclei reticolari del talamo, nell ipotalamo, negli strati profondi della corteccia cerebrale, nello strato delle cellule granulari del cervelletto, nel giro dentato e nello strato lucido dell area CA3 dell ippocampo. I recettori del kainato sembrano svolgere un ruolo importante nello sviluppo e nella plasticità del SNC in quanto alla nascita variano nel numero e nell espressione delle diverse sub-unità. I recettori KA coesistono con gli altri sottotipi recettoriali a livello postsinaptico. L attivazione di questi recettori sembra anche capace di modulare il rilascio di glutammato dalle vescicole sinaptiche (autorecettori)

38 NMDA I recettori NMDA hanno una cinetica molto più lenta (nell ordine di centinaia di millisecondi) dei recettori AMPA e KA e sono altamente permeabili al calcio. Di norma, agiscono insieme ai recettori AMPA e Ka, ma le loro caratteristiche specifiche sono alla base del coinvolgimento di questi in tutti i processi cognitivi superiori ma anche di patologie quali la psicosi o la schizofrenia. Sono in genere composti da 4 sub-unità, ognuna delle quali presenta variante causate esclusivamnte dallo splicing alternativo. Le diverse subunità comprendono: NR1, NR2A-D (comuni) e NR3A-B, NR4 (inibitorie). La subunità NR1 è sempre presente in tutti i recettori di tipo NMDA. La particolarità di tale subunità è data dalla sequenza di aminoacidi che delimita la parete del poro-canale ove sono presenti dei siti di asparagina che rendono questo recettore altamente permeabile al calcio e conferiscono a questo recettore delle altre proprietà come quella di legare ioni magnesio, che in condizioni di non apertura del recettore, si legano all interno del poro acquoso bloccandone completamente la funzionalità. Fig La subunità NR 1 si trova in maniera ubiquitaria in tutte le regioni cerebrali mentre le subunità NR 2A-D sono presenti preferenzialmente nella isocorteccia, nell ippocampo e nel cervelletto.

39 NMDA I recettori NMDA hanno due diversi siti di legame principali che legano rispettivamente i ligandi endogeni L- glutammato, L-aspartato, L-omocistinato e chinolinato, e come ligandi coattivatori, la glicina, D-serina e D-alanina. Siti per inibitori e antagonisti: in questi siti possono legarsi vari inibitori, nella fattispecie Mg2+ e Zn2+, o antagonisti del glutammato che legandosi generano modifiche nella struttura della proteina recettoriale tali da impedire il legame col neurotrasmettitore. Sito fosforilabile: si trova sul versante endoplasmatico ed è fosforilabile dalla PKC. Questo sito è responsabile dell'interazione tra recettori ionotropici e metabotropici. L attivazione del recettore NMDA può avvenire solo se contemporaneamente sia il glutammato che la glicina interagiscono su di esso nei loro siti di legame, oltre a queste condizioni sono però necessarie ulteriori contingenze.
40 NMDA In totale sono tre, quindi, le situazioni necessarie all attivazione del canale: 1. Legame al recettore del ligando glutammato o agonisti. 2. Legame della glicina sul secondo sito di legame Fig Rimozione degli ioni Mg++. Ciò è possibile grazie ad un cambiamento di polarizzazione della membrana plasmatica In conclusione, i recettori NMDA sono sottoposti sia al controllo del ligando che al controllo del potenziale di membrana
41 NMDA Il blocco dovuto al magnesio può essere rimosso anche da poliamine endogene: spermina e spermidina; a basse ed alte concentrazioni, rispettivamente, potenziano ed inibiscono le attività del recettore. Altre sostanze, invece come la ketamina e la fenciclidina agiscono da antagonisti non competitivi, causando fenomeni simili ai sintomi positivi della schizofrenia. Fig Più in generale, gli effetti finali sono di tipo trofico (durante la formazione di contatti sinaptici), di tipo regolatorio della trasmissione sinaptica: nei processi di apprendimento e memorizzazione, potenziamento e depressione sinaptica di lunga durata e sono inoltre coinvolti nella long-term potentiation (LTP) ed, infine, di tipo tossico che possono produrre necrosi neuronale.
42 Potenziamento a lungo termine Bliss e Lomo (1973) Descritto per la prima volta nelle cellule dell ippocampo: consiste nell aumento stabile e duraturo dell'intensità della risposta neuronale dopo che le cellule afferenti di quell'area erano state sollecitate con stimoli elettrici ad alta frequenza. Il neurone postsinaptico permane in una condizione 'attivata' per intervalli di tempo molto più lunghi di quanto avvenga nella trasmissione dell'impulso nervoso.
, grazie all eliminazione del blocco dovuto agli ioni magnesio.")
43 Potenziamento a lungo termine Il LTP avviene in due fasi: 1) la normale trasmissione sinaptica attiva i recettori metabotropici del glutamato (mglur) e i recettori AMPA (AMPAR); 2) scariche ad alta frequenza inducono il potenziamento a lungo termine attivando anche i recettori NMDA (NMDAR), grazie all eliminazione del blocco dovuto agli ioni magnesio. L ingresso di ioni calcio produce reazioni di trasduzione del segnale che fanno aumentare la sensibilità dei recettori AMPA, mediante fosforilazione, e il loro numero complessivo, migliorando l efficienza di risposta della cellula postsinaptica.
, che diffonde nella terminazione presinaptica e produce una regolazione verso l alto della sua efficienza neurotrasmettitoriale")
44 Potenziamento a lungo termine Un altro effetto degli ioni calcio è l induzione della sintesi di un segnale ad azione retrograda, quale l ossido nitrico (NO), che diffonde nella terminazione presinaptica e produce una regolazione verso l alto della sua efficienza neurotrasmettitoriale (upregulation).
45 Potenziamento a lungo termine L LTP coinvolge meccanismi di modificazione della espressione genica e a conferma di ciò è soppresso dagli inibitori della trascrizione e della sintesi proteica. Questi meccanismi prevedono anche la sintesi e il rilascio da parte dei neuroni postsinaptici di un fattore neurotrofico (BDNF) che agisce in maniera retrograda in modo da aumentare l efficienza del neurone presinaptico. L espressione di nuovi prodotti produce aumenti nel numero e nelle dimensioni dei contatti sinaptici. Grazie a meccanismi di questo tipo, la complessità del nostro sistema nervoso aumenta con l esperienza.

46 PLT precoce (memoria a breve termine) Le proteinchinasi attivate agiscono su proteine preesistenti in attesa di essere attivate. Ad es., fosforilazione di recettori AMPA risposta postsinaptica più intensa a parità di glutammato liberato fosforilazione AMPAR *CaMK Processi di rilascio *PKC *PKC NMDAR Ca ++ Messaggero retrogrado * = attivazione persistente

47 PLT tardivo (memoria a lungo termine) Implica la sintesi di nuove proteine 47 spina dendrite camp CaMK MAPK PKA corpo cellulare sintesi proteica CREB nucle o espressione genica CREB = Cyclic AMP-Response Element Binding Protein MAPK = mitogen-activated protein kinase

48 Recettori ionotropi con ligando extracellulare
49 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca2+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )
50 Recettori ionotropi di nucleotidi ciclici Recettori di nucleotidi ciclici CNG HCN Cyclic Nucleotide- Gated Hyperpolarization and Cyclic Nucleotide- Gated Eccitatori - depolarizzazione
51 CNG HCN Sono recettori dei nucleotidi ciclici (camp e cgmp) a struttura tetramerica in cui 2 delle subunità sono omomeri. Le estremità amino- e carbossi-terminale sono intracellulari. Questi recettori sono formati da 6 eliche transmembrana che la attraversano a tutto spessore. C è una sequenza di amminoacidi che entra dal lato extracitoplasmatico, la attraversa in parte, ripiega su se stessa, e fuoriesce dallo stesso lato della membrana (ANSA P). Questa sequenza aminoacidica con l elica S6 partecipa alla formazione della parete del porocanale. Il legame dei ligandi avviene sul versante intracellulare ed è stato osservato che si legano alla sequenza aminoacidica carbossi-terminale in continuità con l elica S6 del recettore. D altra parte, camp e cgmp si formano all interno della cellula e il loro sito di legame deve essere intracellulare. Fig Nei recettori HCN la regione S4 contiene un sensore di voltaggio del tutto analogo a quelli dei recettori regolati da voltaggio. I recettori dei nucleotidi ciclici svolgono compiti importanti nei fotorecettori della retina, nelle cellule sensorie dell epitelio olfattivo oltre che in vari distretti del SNC e P. I recettori HCN sono anche particolarmente importanti per la regolazione del battito cardiaco.
52 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca2+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )

53 Recettori ionotropi regolati dal Ca2+ Si tratta di un gruppo eterogeneo di recettori ionotropi il cui poro acquoso è selettivo per il K+ e si apre quando il sito di legame intracellulare, lega una ione Ca2+. La loro struttura molecolare è simile a quella dei canali regolati da voltaggio, sono quindi formati da complessi tetramerici con 6 domini transmembrana. La subunità α1 ed è la responsabile di tutte le proprietà elettrofisiologiche e farmacologiche di questi canali dato che il poro e il sensore del voltaggio si trovano su di essa. Ogni dominio è formato da 6 α-eliche transmembrana. Il 4 segmento rappresenta il sensore del potenziale mentre tra la 5a e la 6a α- elica c è il sito di legame che caratterizza la selettività ionica. Esistono 9 isoforme della subunità α1, da cui derivano diversi tipi di canali per il calcio È interessante notare che rispondendo all ingresso di Ca2+ all interno dell ambiente citoplasmatico, questi recettori costituiscono importanti sistema di risposta (effettori) all attivazione dei recettori metabotropi che hanno il Ca2+ come secondo messaggero.
 Intermedia permeabilità (recettori IK) Bassa permeabilità (recettori")
54 Recettori ionotropi regolati dal Ca2+ I recettori regolati dal Ca2+, vengono di norma classificati in base alla loro permeabilità per gli ioni K+, nello specifico possiamo avere: Elevata permeabilità (recettori BK) Intermedia permeabilità (recettori IK) Bassa permeabilità (recettori SK)
55 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )
56 Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc ) Questa tipologia di recettori, comunemente detta anche meccanocettori, è attivata da una varietà di stimolo pressori come quelli relativi alla percezione tattile, uditiva dell equilibrio e del posizionamento nello spazio del nostro corpo. Partecipano inoltre, alla regolazione omeostatica nei reni, ed al controllo della pressione sanguigna. Fig Per quanto riguarda i recettori posti nella nostra cute, questi agiscono come rilevatori di pressione grazie ad un contatto diretto con la matrice extracellulare (tessuto connettivo) e le proteine del complesso recettoriale.
57 Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc ) Una seconda tipologia di meccanocettori è composta da quelli posti nell organo del Corti, nell orecchio interno. Questi recettori costituiscono i sensori finali di una serie di vibrazioni che a partono dal timpano, e seguendo la trasmissione attraverso la catena degli ossicini, fino alla finestra ovale, giungono infine a muovere il liquido contenuto nell organo del Corti. Fig Questi spostamenti muovono una membrana chiamata membrana tettoria, che è in diretto contatto con delle cellule ciliate (stereociglia) che a seconda del loro movimento, aprono e chiudono i loro canali, sensibili al Na+ o al Ca2+.
, che da")
58 RECETTORI IONOTROPI Rispetto ai canali ionici voltaggio-dipendenti, i recettori ionotropi presentano una assai maggiore variabilità strutturale. Possono essere costituiti da 3 a 5 subunità, e vengono di norma classificati in classi, comprensive sia dei recettori regolati da neurotrasmettitori (rilasciati nelle giunzioni sinaptiche), che da secondi messaggeri (contenuti all interno delle cellule, Es. Ca+ e camp). Tra le varie classi, i seguenti sono i più importanti per il sistema nervoso: 1. Recettori ionotropi della superfamiglia dei recettori nicotinici 2. Recettori ionotropi del glutammato 3. Recettori ionotropi di nucleotidi ciclici 4. Recettori ionotropi regolati dal Ca+ 5. Recettori ionotropi regolati da altre modalità (luce, tensione meccanica, etc )
59 RECETTORI NON-IONOTROPI, RECETTORI METABOTROPI Si tratta di recettori che rispondono all arrivo di ligandi extracellulari e danno il via ad una cascata di processi metabolici all interno del citoplasma. Sono molto eterogenei dal punto di vista strutturale e possono essere costituiti da una singola catena polipeptidica, o da 2 diverse sub-unità che in presenza del ligando si uniscono fra loro. Dal punto di vista funzionale possiamo classificarli in due grandi famiglie: Recettori legati ad attività enzimatica Recettori legati alla proteina G


60 Recettori legati ad attività enzimatica I recettori legati ad attività enzimatica sono molto eterogenei dal punto di vista strutturale, anche se i più noti ed importanti sono di norma costituiti da due catene polipeptidiche, che a seguito del legame con il ligando uniscono le loro zone nel dominio extracellulare in un dimero. I ligandi in genere comprendono i fattori di crescita (pelle, neurotrofici, fibroblasti e piastrine) o gli ormoni (insulina). La formazione del dimero extra-recettoriale, consente alle porzioni proteiche intracellulari di dare inizio ad una attività enzimatica, che nella maggioranza dei casi è una proteinchinasi.


61 Recettori legati ad attività enzimatica A seconda dell amminoacido che viene fosforilato cambia la denominazione dei recettori: Tirosin-chinasi Istidina-chinasi Serina/treonina chinasi La chinasi in questi casi generalmente consiste: nell autofosforilazione del recettore: I residui autofosforilati servono poi come sito di legame ad alta affinità per altre proteine intracellulari nella fosforilazione di altre proteine.
 dalla proteina con dominio SH2, Grb, che viene fosforilata dal")
62 Recettori legati ad attività enzimatica La via Ras/Raf media l'effetto di molti fattori di crescita e di fattori mitogeni. Ras, che è il prodotto di un protoncogene, funziona come una proteina G e trasferisce il segnale (per mezzo di uno scambio GDP/GTP) dalla proteina con dominio SH2, Grb, che viene fosforilata dal recettore tirosinchinasico. L'attivazione del Ras, a sua volta, attiva Raf,che rappresenta la prima di una sequenza di chinasi serina/treonina,ognuna delle quali fosforila e attiva la chinasi successiva, in ordine: (RAS-RAF- MEK-ERK ), quest'ultimo si lega a c- Fas e c-jun traslocando nel nucleo e legandosi al promoter richiamerà l'rna polimerasi che tradurrà la sequenza dando origine alla ciclina D attivando il processo di riproduzione cellulare.
, e la loro attivazione da il via ad un processo chiamato trasduzione del")
63 Recettori legati alla proteina G I recettori metabotropici sono proteine di membrana legate sul versante citoplasmatico a proteine G (GPCR), e la loro attivazione da il via ad un processo chiamato trasduzione del segnale.


64 Recettori legati alla proteina G Le proteine G sono proteine enterotrimeriche ad attività GTPasica formate da subunità α, β, γ. Nella forma inattiva, la subunità αlpha lega il GDP ed è associata strettamente al complesso formato dalle subunità β e γ. Quando attivata dall interazione con il recettore, la subunità α subisce un cambiamento conformazionale che ne provoca il distacco dal complesso βγ e lo scambio del GDP, ad esso già legato, con una molecola di GTP. In questo modo tale subunità diventa attiva e può interagire con proteine bersaglio (effettore primario) La subunità αlpha non resta nella conformazione attiva per molto tempo, in virtù della sua attività GTPasica intrinseca, che stacca rapidamente il gruppo fosfato terminale dal GTP, trasformandolo in GDP. Questa attività assicura una risposta cellulare strettamente dipendente dall attivazione del recettore metabotropico.
65 Ligando La trasduzione del segnale Recettore metabotropo Enzima - inattivo GDP Proteina G
66 La trasduzione del segnale Unione del ligando con il canale primo messaggero Cambiamento conformazionale del recettore che lo porta a legarsi con il recettore
67 La trasduzione del segnale A seguito del legame con il recettore, la proteina espelle il GDP e si lega invece ad un GTP GDP GTP
68 La trasduzione del segnale La presenza del GTP invece che del GDP causa il distacco della proteina dal recettore e la divisione in due subunità la G-Alpha-GTP e l eterodimero Betagamma. Che restano però ancorati alla membrana plasmatica e sono liberi di muoversi nei dintorni del recettore.
69 La trasduzione del segnale Il complesso G-alpga-GTP incontrerà un enzima che svolgerà il ruolo di effettore primario nel processo di trasduzione. L enzima inizierà ad utilizzare l energia derivante dall idrolisi del GTP in GDP per produrre nuove molecole che agiranno da secondi messaggeri per andare ad attivare l effettore secondario Allo stesso modo anche il complesso betagamma si legherà ad altri effettori primari per dare il via alla produzione di altri processi metabolici
70 Una volta completata l idrolisi del GTP in GDP il complesso alpha si stacca dall effettore primario che terminerà la produzione di secondi messaggeri. Una volta tornata libera nel citoplasma il complesso alpha può incontrare un complesso beta-gamma libero e riformare la proteina G originaria. La trasduzione del segnale
71 La trasduzione del segnale

72


73 I recettori I GPCR possiedono 7 domini ad α-elica transmembrana, un sito di legame per il neurotrasmettitore posto in profondità al centro della porzione che si affaccia nell ambiente extracellulare e un dominio intracitoplasmatico che prende contatto con proteine G. E attraverso l interazione con queste proteine che i recettori metabotropici esercitano i propri effetti. Fig. 6.2
, quali i potenziali postsinaptici eccitatori o inibitori, sia risposte ad insorgenza lenta e di lunga durata.")
74 I recettori La maggior parte dei neurotrasmettitori classici attiva sia recettori ionotropici che metabotropici. Ognuno di questi trasmettitori può indurre sia risposte rapide (msec), quali i potenziali postsinaptici eccitatori o inibitori, sia risposte ad insorgenza lenta e di lunga durata. Tale possibilità fornisce al sistema nervoso la capacità di elaborare l informazione nel tempo e di modularla in risposta alle varie esigenze ambientali. Questo consente ai recettori metabotropi di svolgere ruoli chiave nel SNC e P. La loro importanza è ulteriormente confermata dall elevato numero di geni (nell ordine delle migliaia) che codificano per queste proteine, di cui 350 sono recettori per neutrasmettitori o altri ligandi noti, mentre 150 non hanno ancora avuto una identificazione certa e vengono quindi chiamati recettori orfani.

75 I recettori In generale, i GPCR possono essere raggruppati in 3 famiglie: La famiglia A, è di gran lunga la più numerosa e comprende la maggior parte dei recettori per le monoamine e i neuropeptidi. La famiglia B è costituita dai recettori della secretina, del glucagone e della calcitonina. La famiglia C è costituita principalmente dai recettori metabotropici del glutammato e dai recettori sensibili al Ca2+ Questi recettori interagiscono con il neurotrasmettitore attraverso una regione concava rivolta verso l esterno della cellula. Uno dei ripiegamenti intracellulari di queste molecole media l interazione con le proteine G. Il recettore attiva molto male le proteine G in assenza di ligando, ma lo fa efficacemente nella conformazione stabilizzata dalla sua presenza. Un recettore attivato dal ligando può attivare molte copie di proteine G con un effetto a cascata che amplifica il segnale extracellulare.
76 Le proteine G Esistono diverse classi di proteine G con specificità sia nel bersaglio che nell attività, delle quali le proteine Gs, Gi, e Gq sono quelle meglio conosciute. Stimolatoria CREB Inibitoria Tab. 6.1 Più abbondante nel cervello, recettori muscarinici Processi di percezione nella retina Inibitoria

77 Le proteine G

78 L effettore primario Il più classico fra gli effettori primari durante la trasduzione del segnale è L adenilato ciclasi o adenilico ciclasi, spesso abbreviata come AC, è un enzima della classe delle liasi che catalizza la seguente reazione: ATP = AMP 3',5'-ciclico + pirofosfato Fenomeni biologici che sono stati dimostrati dipendere dall'attività delle ACs comprendono: funzioni cerebrali mediate da neurotrasmettitori; differenziamento cellulare embrionale; fenomeni di resistenza alle azioni molecolari dell'insulina; maturazione degli spermatogoni a spermatozoi; fenomeni di dipendenza da farmaci e/o droghe; soppressione di certe funzioni immunitarie.

79 L effettore primario L'attivazione delle PLC è uno dei primi eventi della risposta cellulare alla stimolazione da parte di un gran numero di segnali extracellulari. Grazie al loro intervento la risposta cellulare si separa in due vie, regolate dai secondi messaggeri DAG e IP3. Il DAG rimane localizzato sulla membrana, così da mediare l'associazione della proteinchinasi C (PKC) alla membrana cellulare, seguita dalla sua attivazione, dando avvio ad una catena di fosforilazioni, che concorreranno alle risposte funzionali della cellula. IP3 è un messaggero intracellulare citoplasmatico, che interagisce con il corrispondente recettore del reticolo endoplasmatico liscio causando la liberazione del Ca2+ immagazzinato e, quindi, l'innalzamento della concentrazione del Ca2+ intracellulare. Il Ca2+ è uno dei principali messaggeri intracellulari ed avvia una ampia serie di risposte cellulari (contrazione, secrezione, ecc.).
 e, mediante interazione con la calmodulina, un altra proteina regolativa, la protein chinasi calcio-calmodulina dipendente (CAMK).")

80 I secondi messaggeri I secondi messaggeri sono responsabili dell induzione di nuove attività cellulari: il camp attiva la protein chinasi A (PKA), gli ioni calcio attivano, insieme al diacilglicerolo, la protein chinasi C (PKC) e, mediante interazione con la calmodulina, un altra proteina regolativa, la protein chinasi calcio-calmodulina dipendente (CAMK). Tutte queste chinasi, a cascata, fosforilano numerosi bersagli proteici all interno della cellula, modificandone l attività. Un altra importante funzione della PKA è l attivazione del fattore di trascrizione CREB che attiva la trascrizione di geni che codificano per proteine che una volta sintetizzate, possono legarsi a canali ionici, enzimi e/o proteine strutturali, modificandone l attività.

81 Classificazione sulla base del ligando I recettori metabotropi possono essere classificati non solo sulla base delle diverse funzioni della sub-unità G-alpha-GTP o sugli effetti relativi all effettore primario o dei secondi messaggeri, ma anche sulla base del ligando a cui questi sono sensibili. Come detto molti di questi recettori sono sensibili agli stessi neurotrasmettitori dei recettori ionotropici e nello specifico possiamo avere: recettore metabotropico del glutammato recettore muscarinico recettore adrenergico recettore GABA B recettore della serotonina recettore della dopamina

82 recettore metabotropico del glutammato I recettori metabotropici glutammatergici (o del glutammato, mglu), fungono da modulatori della sinapsi glutammatergica, in quanto reclutati solo in condizioni di elevate concentrazioni extracellulari del neurotrasmettitore. Si conoscono 8 sottotipi di mglu (mglu1-8) suddivisi in tre gruppi in base all omologia di sequenza, al profilo farmacologico e ai meccanismi di trasduzione del segnale.

83 recettore muscarinico I recettori muscarinici possono essere suddivisi in cinque sottotipi recettoriali: M1, M2, M3, M4, M5 (solo parzialmente caratterizzato), divisi in base alle omologie della sequenza peptidica. I recettori M1, M3 e M5 hanno come secondo messaggero il diacilglicerolo o DAG e l'inositolo trifosfato o IP3. L'aumento della concentrazione di questi due secondi messaggeri nella cellula ne provoca l'attivazione per apertura dei canali del calcio. I recettori M2 e M4 invece, inibiscono l'enzima adenilato ciclasi provocando diminuzione della concentrazione di camp nella cellula. Questo provoca inibizione della cellula neuronale.
84 recettore adrenergico I recettori adrenergici sono recettori di membrana che interagiscono con l'adrenalina e altre catecolamine. A seconda dell'accoppiamento, i recettori adrenergici sono classificati in α (alfa) e β (beta), a loro volta ripartiti in sottotipi: α¹ e α² e β¹, β² e β³. I recettori α¹ sono accoppiati alla fosfolipasi C e producono i loro effetti principalmente grazie al rilascio di Ca 2+ intracellulare I recettori α² sono accoppiati a una proteina G (proteina G inibitrice) la quale inibisce l'azione dell'adenilato ciclasi riducendo pertanto la formazione di camp così come inibiscono i canali del calcio. I recettori β¹ sono accoppiati a Proteine G stimolatrici che innalzano il livello di camp attivando delle proteinchinasi che innalzano il livello di calcio intracellulare. I recettori β² sono accoppiati a una proteina G stimolatrice e attivatrice della Proteinchinasi A che ha come substrato la chinasi della catena leggera della miosina (MLCK), generando così un rilassamento muscolare. Tipo eccitatorio postsinaptico presente in prevalenza sulla muscolatura liscia dei piccoli vasi (resistenze periferiche), la cui stimolazione genera contrazione della muscolatura liscia vasale generando un aumento della pressione recettore presinaptico, presente sulle terminazioni nervose, è deputato alla regolazione della secrezione di neurotrasmettitori sia catecolaminergici che colinergici: la sua attivazione determina una diminuzione della produzione di noradrenalina (feedback negativo) e acetilcolina recettore di tipo eccitatorio accoppiato a proteina Gs come tutti i beta recettori; è presente a livello della muscolatura liscia di alcuni apparati: muscolatura liscia bronchiale, muscolatura liscia gastrointestinale ed è inoltre presente sulla muscolatura liscia di coronarie e grandi vasi che irrorano la muscolatura scheletrica è un recettore di tipo eccitatorio, presente soprattutto a livello del tessuto adiposo. Qui attiva l'enzima lipasi che libera acidi grassi dai trigliceridi

85 recettore GABA B Il recettore GABA B è uno dei due recettori per il GABA (acido γ- amminobutirrico) che sono presenti a livello del sistema nervoso, assieme al recettore GABA-A (recettore ionotropo). I meccanismi effettori associati sono il sistema dell'adenilatociclasi e i canali del calcio e del potassio. In seguito alla loro attivazione, questi recettori provocano diminuzione della conduttanza al calcio e aumento di quella al potassio nelle membrane neuronali. L'effetto sulla conduttanza del calcio è principalmente associato alle correnti presinaptiche mediata da canali del calcio di tipo P/Q e N; la riduzione del calcio intracellulare nel terminale presinaptico è il meccanismo con cui questi recettori regolano il rilascio di neurotrasmettitori. La conduttanza del potassio è invece associata a recettori GABA B localizzati a livello postsinaptico; l'aumento della permeabilità al potassio, quindi la sua uscita dalla cellula, si traduce in un effetto iperpolarizzante della membrana, con conseguente innalzamento della soglia di eccitabilità della stessa cellula.

86 recettore della serotonina I recettori metabotropi serotoninergici (o della serotonina) sono sei e Si trovano nel sistema nervoso centrale e periferico, sulla membrana cellulare delle cellule nervose e in altri tipi di cellule. Regolano sia l'eccitazione sia l'inibizione della trasmissione sinaptica. Modulano il rilascio di neurotrasmettitori, tra cui l'acido glutammico, GABA, dopamina, adrenalina / noradr enalina e acetilcolina, così come molti ormoni, tra cui l'ossitocina, prolattina, vasopressina, cortisolo, corti cotropina, la sostanza P, ed altri. Influenzano vari processi biologici e neurologici, come l'aggressività, ansia, appetito, conoscenza, appren dimento, memoria, umore, nausea, sonno e termor egolazione. I recettori della serotonina sono il bersaglio di una varietà di farmaci e sostanze stupefacenti, tra cui molti antidepressivi, antipsicotici, anoressizzanti, e allucinogeni.

87 recettore della dopamina Nella classe dei recettori dopaminergici esistono cinque sottotipi recettoriali, codificati da geni diversi. Ad oggi sono stati clonati 5 geni che codificano per recettori dopaminergici, di conseguenza sono stati identificati i recettori dopaminergici D1, D2, D3, D4, D5. I recettori dopaminergici sono stati classificati in due sottoclassi (D1-like) e (D2-like), sulla base della classificazione filogenetica e al tipo di proteina G a cui si accoppiano. D1-like In questa famiglia si trovano i recettori D1 e D5. L'attivazione di questi recettori porta all'attivazione dell'adenilato ciclasi, in quanto accoppiati a proteine G stimolatorie. D2-like I recettori D2, D3, D4 appartengono alla sottoclasse dei recettori dopaminergici D2-like. Questi recettori sono accoppiati a proteine G inibitorie. L'attivazione di questi recettori ad opera della dopamina ed altri agonisti comporta l'inibizione dell'adenilato ciclasi e quindi ad una diminuzione del secondo messaggero camp.
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
TRASDUZIONE DEL SEGNALE CELLULARE
 TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
Glutamatergic pathways in the brain
 Glutamatergic pathways in the brain Principali vie glutammatergiche Vie cortico-corticali Vie corticotalamiche Vie extrapiramidali(corticostriatali) Vie ippocampali Altre vie glutammatergiche Proiezioni
Glutamatergic pathways in the brain Principali vie glutammatergiche Vie cortico-corticali Vie corticotalamiche Vie extrapiramidali(corticostriatali) Vie ippocampali Altre vie glutammatergiche Proiezioni
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE
 COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Fondamenti di Chimica Farmaceutica. Recettori
 Fondamenti di Chimica Farmaceutica Recettori Recettori In biochimica, un recettore è una proteina, transmembrana o intracellulare, che si lega ad uno specifico ligando, causando nel recettore una variazione
Fondamenti di Chimica Farmaceutica Recettori Recettori In biochimica, un recettore è una proteina, transmembrana o intracellulare, che si lega ad uno specifico ligando, causando nel recettore una variazione
Neurotrasmettitori. I neurotrasmettitori appartengono a due grandi categorie:
 Neurotrasmettitori e recettori sinaptici Neurotrasmettitori I neurotrasmettitori appartengono a due grandi categorie: Neurotrasmettitori classici: molecole a basso peso molecolare: Acetilcolina Monoamine
Neurotrasmettitori e recettori sinaptici Neurotrasmettitori I neurotrasmettitori appartengono a due grandi categorie: Neurotrasmettitori classici: molecole a basso peso molecolare: Acetilcolina Monoamine
Cellula presinaptica e cellula postsinaptica. Il messaggio nervoso segue la direzione mostrata dalle frecce.
 La trasmissione sinaptica Cellula presinaptica e cellula postsinaptica. Il messaggio nervoso segue la direzione mostrata dalle frecce. 1 Microfotografia a scansione elettronica del corpo cellulare di un
La trasmissione sinaptica Cellula presinaptica e cellula postsinaptica. Il messaggio nervoso segue la direzione mostrata dalle frecce. 1 Microfotografia a scansione elettronica del corpo cellulare di un
FATTORI DI CRESCITA COME MESSAGGERI
 RECETTORI ASSOCIATI A PROTEINE CHINASI Sono proteine che hanno sia funzione di recettore che attività enzimatica chinasica. L interazione con il ligando specifico stimola l attività della chinasi e il
RECETTORI ASSOCIATI A PROTEINE CHINASI Sono proteine che hanno sia funzione di recettore che attività enzimatica chinasica. L interazione con il ligando specifico stimola l attività della chinasi e il
Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre
, dall interazione con altre") Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre cellule (contatto cellula-cellula) o con strutture extracellulari
Una risposta cellulare specifica può essere determinata dalla presenza di mediatori chimici (ormoni o altre molecole), dall interazione con altre cellule (contatto cellula-cellula) o con strutture extracellulari
I RECETTORI PER NEUROTRASMETTITORI E ORMONI: CARATTERISTICHE GENERALI. 1.Localizzazione: recettori di membrana e recettori intracellulari
 I RECETTORI PER NEUROTRASMETTITORI E ORMONI: CARATTERISTICHE GENERALI Classificazione per: 1.Localizzazione: recettori di membrana e recettori intracellulari 2.Struttura 3.Modalità di funzionamento 4.Agonista
I RECETTORI PER NEUROTRASMETTITORI E ORMONI: CARATTERISTICHE GENERALI Classificazione per: 1.Localizzazione: recettori di membrana e recettori intracellulari 2.Struttura 3.Modalità di funzionamento 4.Agonista
Trasduzione del segnale
 Trasduzione del segnale Modalità di segnalazione tra cellule JUXTACRINA proteine transmembrana mediatori locali ormoni Il legame di una sostanza al suo recettore di membrana porta la cellula a cambiare
Trasduzione del segnale Modalità di segnalazione tra cellule JUXTACRINA proteine transmembrana mediatori locali ormoni Il legame di una sostanza al suo recettore di membrana porta la cellula a cambiare
Differenze tra sinapsi neuromuscolare e sinapsi centrali
 Sinapsi centrali ed Integrazione sinaptica Differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore solo Ach. Le sinapsi centrali utilizzano
Sinapsi centrali ed Integrazione sinaptica Differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore solo Ach. Le sinapsi centrali utilizzano
Il Glutammato ed i suoi recettori
 Il Glutammato ed i suoi recettori Ionotropi NMDA (N- me%l- D- aspartato) : controlla canali ionici Na + /K + /Ca ++ AMPA (alfa- Amino- 3- Idrossi- 5- Me%l- 4- isoxazolone propionato) : controlla canali
Il Glutammato ed i suoi recettori Ionotropi NMDA (N- me%l- D- aspartato) : controlla canali ionici Na + /K + /Ca ++ AMPA (alfa- Amino- 3- Idrossi- 5- Me%l- 4- isoxazolone propionato) : controlla canali
Io Non sono un recettore!
 I recettori portano il messaggio extracellulare direttamente nel citoplasma (recettori di membrana) o nel nucleo (recettori nucleari) Io Non sono un recettore! È essenziale per la loro sopravvivenza che
I recettori portano il messaggio extracellulare direttamente nel citoplasma (recettori di membrana) o nel nucleo (recettori nucleari) Io Non sono un recettore! È essenziale per la loro sopravvivenza che
SISTEMA NERVOSO. Sistema nervoso periferico. Sistema nervoso centrale. simpatico parasimpatico Sezione afferente.
 SISTEMA NERVOSO Sistema nervoso periferico Sistema nervoso centrale Sezione efferente Sezione afferente Sistema autonomo Sistema somatico simpatico parasimpatico INNERVAZIONE SIMPATICA E PARASIMPATICA
SISTEMA NERVOSO Sistema nervoso periferico Sistema nervoso centrale Sezione efferente Sezione afferente Sistema autonomo Sistema somatico simpatico parasimpatico INNERVAZIONE SIMPATICA E PARASIMPATICA
GLUTAMMATO E ACIDO γ-aminobutirrico (GABA)
") GLUTAMMATO E ACIDO γ-aminobutirrico (GABA) GLUTAMMATO E ACIDO γ-aminobutirrico GLUTAMMATO E GLI AMINOACIDI ECCITATORI Ossalacetato Glioxilato Glicina Modulatore allosterico positivo dei recettori NMDA
GLUTAMMATO E ACIDO γ-aminobutirrico (GABA) GLUTAMMATO E ACIDO γ-aminobutirrico GLUTAMMATO E GLI AMINOACIDI ECCITATORI Ossalacetato Glioxilato Glicina Modulatore allosterico positivo dei recettori NMDA
Il neurone. I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi.
 attraverso le sinapsi.") Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
Il neurone I neuroni trasmettono l informazione ad altri neuroni o alle cellule effettrici (cellule muscolari) attraverso le sinapsi. Trasmissione sinaptica Ingresso: segnali sinaptici Integrazione: segnali
Richiede comunicazione bidirezionale tra compartimento pre- e postsinaptico finalizzata all attivazione di diversi meccanismi cellulari e molecolari
 Plasticità sinaptica: Sinaptogenesi Rimodellamento sinaptico Richiede comunicazione bidirezionale tra compartimento pre- e postsinaptico finalizzata all attivazione di diversi meccanismi cellulari e molecolari
Plasticità sinaptica: Sinaptogenesi Rimodellamento sinaptico Richiede comunicazione bidirezionale tra compartimento pre- e postsinaptico finalizzata all attivazione di diversi meccanismi cellulari e molecolari
Il Glutammato ed i suoi recettori
 Il Glutammato ed i suoi recettori Ionotropi NMDA (N- me%l- D- aspartato) : controlla canali ionici Na + /K + /Ca ++ AMPA (alfa- Amino- 3- Idrossi- 5- Me%l- 4- isoxazolone propionato) : controlla canali
Il Glutammato ed i suoi recettori Ionotropi NMDA (N- me%l- D- aspartato) : controlla canali ionici Na + /K + /Ca ++ AMPA (alfa- Amino- 3- Idrossi- 5- Me%l- 4- isoxazolone propionato) : controlla canali
Dipendente da contatto Segnali di posizione (migrazione dei leucociti) Paracrina Raggio breve (neurotrasmettitori)
 Paracrina Raggio breve (neurotrasmettitori)") Dipendente da contatto Segnali di posizione (migrazione dei leucociti) Paracrina Raggio breve (neurotrasmettitori) Autocrina Endocrina Agisce sulla stessa cellula che Fattori rilasciati in circolazione
Dipendente da contatto Segnali di posizione (migrazione dei leucociti) Paracrina Raggio breve (neurotrasmettitori) Autocrina Endocrina Agisce sulla stessa cellula che Fattori rilasciati in circolazione
I RECETTORI CANALE: IL RECETTORE NICOTINICO PER L ACETILCOLINA E IL RECETTORE-CANALE PER IL GABA
 I RECETTORI CANALE: IL RECETTORE NICOTINICO PER L ACETILCOLINA E IL RECETTORE-CANALE PER IL GABA ASPETTI GENERALI SULL ORGANIZZAZIONE MOLECOLARE, SUL MODO DI FUNZIONARE E SULLA MODULAZIONE FARMACOLOGICA
I RECETTORI CANALE: IL RECETTORE NICOTINICO PER L ACETILCOLINA E IL RECETTORE-CANALE PER IL GABA ASPETTI GENERALI SULL ORGANIZZAZIONE MOLECOLARE, SUL MODO DI FUNZIONARE E SULLA MODULAZIONE FARMACOLOGICA
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA)
") Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Modalità di trasporto di membrana (DIFFUSIONE FACILITATA) Classi di proteine di trasporto presenti nella membrana plasmatica (o CARRIER) Porta girevole Porta aperta I canali ionici Sezione trasversale
Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari
 Corso di laurea magistrale in BIOTECNOLOGIE DELLA RIPRODUZIONE UNIVERSITA DEGLI STUDI DI TERAMO Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari Prof.ssa Luisa Gioia Trasmissione
Corso di laurea magistrale in BIOTECNOLOGIE DELLA RIPRODUZIONE UNIVERSITA DEGLI STUDI DI TERAMO Corso di recupero Fisiologia cellulare/ Laboratorio di colture cellulari Prof.ssa Luisa Gioia Trasmissione
comunicazione cellulare Prof. Davide Cervia - Fisiologia Fisiologia della cellula: comunicazione cellulare
 comunicazione cellulare 1 2 3 4 neurotrasmettitori sono più di cento caratteristiche: sintesi immagazzinamento e rilascio diffusione e legame selettività blocco da parte di antagonisti termine dell azione
comunicazione cellulare 1 2 3 4 neurotrasmettitori sono più di cento caratteristiche: sintesi immagazzinamento e rilascio diffusione e legame selettività blocco da parte di antagonisti termine dell azione
Cell Signaling. Qualsiasi tipo di comunicazione tra le cellule
 Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La
Cell Signaling Qualsiasi tipo di comunicazione tra le cellule I segnali intercellulari L evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l altra. La
Glutamatergic pathways in the brain
 Glutamatergic pathways in the brain Principali vie glutammatergiche Vie cortico-corticali Vie corticotalamiche Vie extrapiramidali (corticostriatali) Vie ippocampali Altre vie glutammatergiche Proiezioni
Glutamatergic pathways in the brain Principali vie glutammatergiche Vie cortico-corticali Vie corticotalamiche Vie extrapiramidali (corticostriatali) Vie ippocampali Altre vie glutammatergiche Proiezioni
Laurea Specialistica in Neuropsicobiologia. 1. Quale delle seguenti condizioni non influisce sull assorbimento gastrointestinale dei farmaci?
 Laurea Specialistica in Neuropsicobiologia 1. Quale delle seguenti condizioni non influisce sull assorbimento gastrointestinale dei farmaci? a. grado di ionizzazione b. la legge d azione di massa c. l
Laurea Specialistica in Neuropsicobiologia 1. Quale delle seguenti condizioni non influisce sull assorbimento gastrointestinale dei farmaci? a. grado di ionizzazione b. la legge d azione di massa c. l
Comunicazione cellulare 1) Se le cellule sono in contatto diretto tra loro, esse possono comunicare attraverso l interazione tra una molecola di
 Se le cellule sono in contatto diretto tra loro, esse possono comunicare attraverso l interazione tra una molecola di") Comunicazione cellulare 1) Se le cellule sono in contatto diretto tra loro, esse possono comunicare attraverso l interazione tra una molecola di superficie di una cellula e recettori presenti sulla superficie
Comunicazione cellulare 1) Se le cellule sono in contatto diretto tra loro, esse possono comunicare attraverso l interazione tra una molecola di superficie di una cellula e recettori presenti sulla superficie
BIOSEGNALAZIONE. La ricezione e la trasmissione delle informazioni extracellulari Parte I
 BIOSEGNALAZIONE La ricezione e la trasmissione delle informazioni extracellulari Parte I La coordinazione delle attività metaboliche nei diversi tessuti e organi dei mammiferi avviene mediante il sistema
BIOSEGNALAZIONE La ricezione e la trasmissione delle informazioni extracellulari Parte I La coordinazione delle attività metaboliche nei diversi tessuti e organi dei mammiferi avviene mediante il sistema
La maggior parte dei farmaci però produce i suoi effetti in seguito all interazione con specifici componenti tessutali macromolecolari
 FARMACODINAMICA È quella branca della Farmacologia che studia i meccanismi con cui i farmaci sono in grado di modificare le funzioni dell organismo. È la scienza che contribuisce a costruire e solidificare
FARMACODINAMICA È quella branca della Farmacologia che studia i meccanismi con cui i farmaci sono in grado di modificare le funzioni dell organismo. È la scienza che contribuisce a costruire e solidificare
giunzioni comunicanti connessone cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali precoci veloci bidirezionali
 La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
La sinapsi Sinapsi elettriche giunzioni comunicanti ioni attraversano il canale centrale detto connessone presenti tra cellule gliali, muscolo liscio e cardiaco, neuroni del SNC negli stadi embrionali
La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico.
 La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico. N1 N2 Punto di discontinuità Elettriche Chimiche: Eccitatorie Inibitorie La sinapsi I neuroni comunicano
La sinapsi: zona di contatto tra due neuroni responsabile della trasmissione del segnale elettrico. N1 N2 Punto di discontinuità Elettriche Chimiche: Eccitatorie Inibitorie La sinapsi I neuroni comunicano
La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi elettro-chimici.
 LA TRASMISSIONE SINAPTICA Il neurone : unità elementare responsabile dei messaggi nervosi La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi
LA TRASMISSIONE SINAPTICA Il neurone : unità elementare responsabile dei messaggi nervosi La più nota proprietà del neurone è quella di ricevere e trasmettere segnali costituiti principalmente da eventi
Tipi di neurotrasmettitori
 Tipi di neurotrasmettitori Neurotrasmettitori classici: molecole a basso peso molecolare, di varia natura: acetilcolina amine biogene (dopamina, adrenalina, noradrenalina) istamina aminoacidi (GABA, glicina,
Tipi di neurotrasmettitori Neurotrasmettitori classici: molecole a basso peso molecolare, di varia natura: acetilcolina amine biogene (dopamina, adrenalina, noradrenalina) istamina aminoacidi (GABA, glicina,
1) Sostanze liberate A B Recettori ormoni altre 2) Conduzione di segnali elettrici
 Sostanze liberate A B Recettori ormoni altre 2) Conduzione di segnali elettrici") MECCANISMI DI TRASMISSIONE DEI MESSAGGI LA COMUNICAZIONE CELLULARE 1) Sostanze liberate A B Recettori ormoni altre 2) Conduzione di segnali elettrici CONDUZIONE SALTATORIA PROPAGAZIONE DI UN PdA LUNGO
MECCANISMI DI TRASMISSIONE DEI MESSAGGI LA COMUNICAZIONE CELLULARE 1) Sostanze liberate A B Recettori ormoni altre 2) Conduzione di segnali elettrici CONDUZIONE SALTATORIA PROPAGAZIONE DI UN PdA LUNGO
FARMACODINAMICA FARMACOCINETICA
 FARMACODINAMICA FARMACOCINETICA Farmacodinamica La farmacodinamica studia gli effetti biochimici e fisiologici e il meccanismo d azione dei farmaci ² identificare i siti d azione dei farmaci ² delineare
FARMACODINAMICA FARMACOCINETICA Farmacodinamica La farmacodinamica studia gli effetti biochimici e fisiologici e il meccanismo d azione dei farmaci ² identificare i siti d azione dei farmaci ² delineare
LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA
 LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA Lezione_6 1 STUTTURA MOTONEURONE FIBRA MUSCOLARE GIUNZIONE NMUSCOLARE Lezione_6 2 Sintesi e degradazione di acetilcolina ACh è rilasciata da
LA GIUNZIONE NEUROMUSCOLARE COME ESEMPIO DI SINAPSI CHIMICA Lezione_6 1 STUTTURA MOTONEURONE FIBRA MUSCOLARE GIUNZIONE NMUSCOLARE Lezione_6 2 Sintesi e degradazione di acetilcolina ACh è rilasciata da
scaricatoda Come è noto il SNC controlla i muscoli e gli altri organi attraverso la branca efferente del SNP.
 Come è noto il SNC controlla i muscoli e gli altri organi attraverso la branca efferente del SNP. Il sistema parasimpatico controlla la routine e le attività tranquille del vivere quotidiano, come per
Come è noto il SNC controlla i muscoli e gli altri organi attraverso la branca efferente del SNP. Il sistema parasimpatico controlla la routine e le attività tranquille del vivere quotidiano, come per
Principi di farmacodinamica Filippo Caraci
 Università degli Studi di Catania Modulo di Psicofarmacologia Corso di Laurea Magistrale in Psicologia Principi di farmacodinamica Filippo Caraci Farmacodinamica Farmacodinamica e la descrizione delle
Università degli Studi di Catania Modulo di Psicofarmacologia Corso di Laurea Magistrale in Psicologia Principi di farmacodinamica Filippo Caraci Farmacodinamica Farmacodinamica e la descrizione delle
POTENZIALE DI MEMBRANA A RIPOSO. Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI
 POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
POTENZIALE DI MEMBRANA A RIPOSO Le informazioni all'interno del sistema nervoso vengono scambiate mediante SEGNALI ELETTRICI La membrana plasmatica delle cellule è POLARIZZATA (esiste una differenza nella
Il sistema nervoso periferico: divisione efferente
 Il sistema nervoso periferico: divisione efferente 7 1 7 2 7 3 Sistema nervoso autonomo Parasimpatico Simpatico iride ghiandola lacrimale ghiandola salivare cuore polmoni stomaco e pancreas colon Midollare
Il sistema nervoso periferico: divisione efferente 7 1 7 2 7 3 Sistema nervoso autonomo Parasimpatico Simpatico iride ghiandola lacrimale ghiandola salivare cuore polmoni stomaco e pancreas colon Midollare
1. Nel potenziale d azione delle cellule nervose, la fase di ripolarizzazione è dovuta a:
 1. Nel potenziale d azione delle cellule nervose, la fase di ripolarizzazione è dovuta a: a) chiusura dei canali voltaggio-dipendenti del K + e apertura dei canali del Na + b) chiusura dei canali del Na
1. Nel potenziale d azione delle cellule nervose, la fase di ripolarizzazione è dovuta a: a) chiusura dei canali voltaggio-dipendenti del K + e apertura dei canali del Na + b) chiusura dei canali del Na
Corso di Laurea Magistrale in Medicina e Chirurgia. Biofisica e Fisiologia I. Muscolo liscio
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Muscolo liscio Muscolo cardiaco scheletrico liscio MUSCOLO LISCIO La muscolatura liscia si trova nelle pareti degli organi cavi.
** Depolarizzazione: sinapsi eccitatoria, l elemento postsinaptico può generare un potenziale d azione.
 Sinapsi chimiche sia eccitatorie che inibitorie. Il legame neurotrasmettitore-recettore può infatti determinare una modificazione di permeabilità ionica che porta a: ** Depolarizzazione: sinapsi eccitatoria,
Sinapsi chimiche sia eccitatorie che inibitorie. Il legame neurotrasmettitore-recettore può infatti determinare una modificazione di permeabilità ionica che porta a: ** Depolarizzazione: sinapsi eccitatoria,
FARMACOLOGIA DEL SISTEMA NERVOSO AUTONOMO SIMPATICO. Il sistema ortosimpatico media le risposte della reazione di lotta e fuga
 FARMACOLOGIA DEL SISTEMA NERVOSO AUTONOMO SIMPATICO Il sistema ortosimpatico media le risposte della reazione di lotta e fuga Le basi anatomiche del sistema nervoso autonomo Reazioni connesse alla digestione
FARMACOLOGIA DEL SISTEMA NERVOSO AUTONOMO SIMPATICO Il sistema ortosimpatico media le risposte della reazione di lotta e fuga Le basi anatomiche del sistema nervoso autonomo Reazioni connesse alla digestione
Principali differenze tra sinapsi neuromuscolare e sinapsi centrali
 Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Sinapsi centrali ed Integrazione sinaptica Principali differenze tra sinapsi neuromuscolare e sinapsi centrali Le sinapsi neuromuscolari utilizzano come neurotrasmettitore Ach Le sinapsi centrali utilizzano
Comunicazione Cellulare
 Comunicazione Cellulare Negli organismi pluricellulari le cellule comunicano a breve medio o lungo raggio mediante molecole segnale Rilasciate da cellula che le produce e ricevute da cellula bersaglio
Comunicazione Cellulare Negli organismi pluricellulari le cellule comunicano a breve medio o lungo raggio mediante molecole segnale Rilasciate da cellula che le produce e ricevute da cellula bersaglio
Farmacologia del sistema serotoninergico
 Farmacologia del sistema serotoninergico Trasmissione serotoninergica Effetti della serotonina (5-HT, 5-idrossitriptamina): Regolazione del tono della muscolatura liscia sistema cardiovascolare tratto
Farmacologia del sistema serotoninergico Trasmissione serotoninergica Effetti della serotonina (5-HT, 5-idrossitriptamina): Regolazione del tono della muscolatura liscia sistema cardiovascolare tratto
FARMACODINAMICA. La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci.
 FARMACODINAMICA La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci. La farmacodinamica si propone di: * identificare i siti d azione dei farmaci * delineare le interazioni
FARMACODINAMICA La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci. La farmacodinamica si propone di: * identificare i siti d azione dei farmaci * delineare le interazioni
Sostanza liberata da una cellula nervosa presinaptica, in grado di agire in maniera specifica su una cellula postsinaptica. In alcune sinapsi i
 Neurotrasmettitori e recettori sinaptici Neurotrasmettitore Sostanza liberata da una cellula nervosa presinaptica, in grado di agire in maniera specifica su una cellula postsinaptica. In alcune sinapsi
Neurotrasmettitori e recettori sinaptici Neurotrasmettitore Sostanza liberata da una cellula nervosa presinaptica, in grado di agire in maniera specifica su una cellula postsinaptica. In alcune sinapsi
Il sistema nervoso autonomo: simpatico e parasimpatico
 Il sistema nervoso autonomo: simpatico e parasimpatico Proprietà dell omeostasi: 1- mantenimento della costanza dell ambiente interno 2- aumento o diminuzione del controllo tonico 3- controllo antagonista
Il sistema nervoso autonomo: simpatico e parasimpatico Proprietà dell omeostasi: 1- mantenimento della costanza dell ambiente interno 2- aumento o diminuzione del controllo tonico 3- controllo antagonista
Summary diagram of the acquisition and storage of declarative versus procedural information.
 LA MEMORIA Summary diagram of the acquisition and storage of declarative versus procedural information. Principali regioni encefaliche con funzione regolatrice BASI MOLECOLARI DEI PROCESSI DI MEMORIZZAZIONE
LA MEMORIA Summary diagram of the acquisition and storage of declarative versus procedural information. Principali regioni encefaliche con funzione regolatrice BASI MOLECOLARI DEI PROCESSI DI MEMORIZZAZIONE
Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A
 Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A. 2015-2016 Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente Riflessi nervosi: si attuano
Corso di Laurea Magistrale in Medicina e Chirurgia Fisiologia e Biofisica A.A. 2015-2016 Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente Riflessi nervosi: si attuano
TARGETS DEI FARMACI ATTUALMENTE IN USO
 TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
TARGETS DEI FARMACI ATTUALMENTE IN USO NEURONI Il corpo umano contiene circa 100 miliardi di neuroni. Sono cellule specializzate nel trasmettere e ricevere informazioni. Possono variare forma e dimensione
Recettore canale per il GABA
 Recettore canale per il GABA Il recettore GABA A è un canale ionico permeabile al cloro La stimolazione produce una iperpolarizzazione che riduce la eccitabilità cellulare L attività recettoriale è regolata
Recettore canale per il GABA Il recettore GABA A è un canale ionico permeabile al cloro La stimolazione produce una iperpolarizzazione che riduce la eccitabilità cellulare L attività recettoriale è regolata
** Depolarizzazione: sinapsi eccitatoria, l elemento postsinaptico può generare un potenziale d azione.
 Sinapsi chimiche sia eccitatorie che inibitorie. Il legame neurotrasmettitore-recettore può infatti determinare una modificazione di permeabilità ionica che porta a: ** Depolarizzazione: sinapsi eccitatoria,
Sinapsi chimiche sia eccitatorie che inibitorie. Il legame neurotrasmettitore-recettore può infatti determinare una modificazione di permeabilità ionica che porta a: ** Depolarizzazione: sinapsi eccitatoria,
Cambiamenti durante l esercizio:
 Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della ventilazione polmonare Aumento della gittata cardiaca Aumento della
Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della ventilazione polmonare Aumento della gittata cardiaca Aumento della
Regolazione enzimatica Isoenzimi
 Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
FARMACODINAMICA. La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci. identificare i siti d azione dei farmaci
 FARMACODINAMICA La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci. La farmacodinamica si propone di: identificare i siti d azione dei farmaci delineare le interazioni
FARMACODINAMICA La farmacodinamica studia gli effetti biochimici e il meccanismo d azione dei farmaci. La farmacodinamica si propone di: identificare i siti d azione dei farmaci delineare le interazioni
Lezioni su NT, recettori, sistemi di modulazione del SNC. Farmacologia generale e molecolare
 Lezioni su NT, recettori, sistemi di modulazione del SNC Farmacologia generale e molecolare a cura di F. Clementi e G. Fumagalli Capitoli 4 5 6 Capitoli 18 19 20-21 -22 solo per il SNC SISTEMA NERVOSO
Lezioni su NT, recettori, sistemi di modulazione del SNC Farmacologia generale e molecolare a cura di F. Clementi e G. Fumagalli Capitoli 4 5 6 Capitoli 18 19 20-21 -22 solo per il SNC SISTEMA NERVOSO
Comunicazione cellulare
 Comunicazione cellulare Comunicazione mediante messaggi chimici. Funzioni dei recettori chimici e trasduzione del segnale. Recettori metabotropici. Proteine G trimeriche e monometriche. Adenilato ciclasi
Comunicazione cellulare Comunicazione mediante messaggi chimici. Funzioni dei recettori chimici e trasduzione del segnale. Recettori metabotropici. Proteine G trimeriche e monometriche. Adenilato ciclasi
Meccanismo d azione dei NT nelle sinapsi dirette. 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie
 Meccanismo d azione dei NT nelle sinapsi dirette 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie 1) La giunzione neuromuscolare La prima sinapsi ad essere studiata nel dettaglio
Meccanismo d azione dei NT nelle sinapsi dirette 1.La giunzione neuromuscolare 2.Sinapsi eccitatorie 3.Sinapsi inibitorie 1) La giunzione neuromuscolare La prima sinapsi ad essere studiata nel dettaglio
Trasporti attraverso. Proteine di trasporto
 Trasporti attraverso la membrana Proteine di trasporto Diffusione Facilitata Caratteristiche dei trasporti mediati I carriers sono dotati di specificità Sono soggetti a saturazione Possono essere bloccati
Trasporti attraverso la membrana Proteine di trasporto Diffusione Facilitata Caratteristiche dei trasporti mediati I carriers sono dotati di specificità Sono soggetti a saturazione Possono essere bloccati
I CANALI A CONTROLLO DI POTENZIALE
 I CANALI A CONTROLLO DI POTENZIALE I canali a controllo di potenziale (VOC, voltage operated channels) costituiscono una superfamiglia di proteine transmembrana formanti canali I VOC regolano la permeabilità
I CANALI A CONTROLLO DI POTENZIALE I canali a controllo di potenziale (VOC, voltage operated channels) costituiscono una superfamiglia di proteine transmembrana formanti canali I VOC regolano la permeabilità
BIOCHIMICA DEL SISTEMA NERVOSO
 BIOCHIMICA DEL SISTEMA NERVOSO 2,5-3.0% in peso Per unità di peso, rispetto ad altri tessuti: a) Maggior fabbisogno energetico e maggior consumo di ossigeno b) Maggior richiesta di glucosio c) Elevato
BIOCHIMICA DEL SISTEMA NERVOSO 2,5-3.0% in peso Per unità di peso, rispetto ad altri tessuti: a) Maggior fabbisogno energetico e maggior consumo di ossigeno b) Maggior richiesta di glucosio c) Elevato
5. Trasporto di membrana
 5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
5. Trasporto di membrana contiene materiale protetto da copyright, ad esclusivo uso personale; 1 non è consentita diffusione ed utilizzo di tipo commerciale Le membrane biologiche hanno una permeabilità
La trasduzione del segnale intracellulare
 La trasduzione del segnale intracellulare Chemical signaling mechanisms I RECETTORI Farmaci antagonisti I beta-bloccanti per l ipertensione competono con adrenalina e noradrenalina per il recettore beta-adrenergico.
La trasduzione del segnale intracellulare Chemical signaling mechanisms I RECETTORI Farmaci antagonisti I beta-bloccanti per l ipertensione competono con adrenalina e noradrenalina per il recettore beta-adrenergico.
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
11/10/16. Il concetto di MOSAICO FLUIDO. Fosfolipidi Colesterolo Glicolipidi
 Il concetto di MOSAICO FLUIDO Fosfolipidi Colesterolo Glicolipidi 1 DIFFUSIONE SEMPLICE 2 Pressione osmotica Il problema della pressione osmotica 3 Diverse soluzioni al problema della pressione osmotica
Il concetto di MOSAICO FLUIDO Fosfolipidi Colesterolo Glicolipidi 1 DIFFUSIONE SEMPLICE 2 Pressione osmotica Il problema della pressione osmotica 3 Diverse soluzioni al problema della pressione osmotica
scaricato da Caratteristiche di permeabilità della membrana plasmatica
 Caratteristiche di permeabilità della membrana plasmatica Se un solvente è più concentrato in un punto della soluzione esso tenderà a diffondersi lungo il gradiente di concentrazione, dalla parte più concentrata
Caratteristiche di permeabilità della membrana plasmatica Se un solvente è più concentrato in un punto della soluzione esso tenderà a diffondersi lungo il gradiente di concentrazione, dalla parte più concentrata
1. RECETTORI IONOTROPICI A CONTROLLO DI LIGANDO o ROC
 1. RECETTORI IONOTROPICI A CONTROLLO DI LIGANDO o ROC ATP Acetilcolina Glutammato NMDA AMPA/cainato SUBUNITA': ogni recettore è formato da 5 ( o 4) subunità nicotinico MUSCOLARE (acetilcolina) nicotinico
1. RECETTORI IONOTROPICI A CONTROLLO DI LIGANDO o ROC ATP Acetilcolina Glutammato NMDA AMPA/cainato SUBUNITA': ogni recettore è formato da 5 ( o 4) subunità nicotinico MUSCOLARE (acetilcolina) nicotinico
Università degli Studi di Camerino Laurea Specialistica in Farmacia Scuola di Scienze del Farmaco e dei Prodotti della Salute Facoltà di Farmacia
 Università degli Studi di Camerino Laurea Specialistica in Farmacia Scuola di Scienze del Farmaco e dei Prodotti della Salute Facoltà di Farmacia SINTESI DI TETRAIDRODIBENZOSSAZECINE COME POTENZIALI LIGANDI
Università degli Studi di Camerino Laurea Specialistica in Farmacia Scuola di Scienze del Farmaco e dei Prodotti della Salute Facoltà di Farmacia SINTESI DI TETRAIDRODIBENZOSSAZECINE COME POTENZIALI LIGANDI
COMUNICAZIONE INTERCELLULARE
 COMUNICAZIONE INTERCELLULARE TRASFERIMENTO DIRETTO DI SEGNALI CHIMICI E ELETTRICI ATTRAVERSO GIUNZIONI COMUNICANTI COMUNICAZIONE CHIMICA LOCALE (SOSTANZE PARACRINE E AUTOCRINE) COMUNICAZIONE A LUNGA DISTANZA
COMUNICAZIONE INTERCELLULARE TRASFERIMENTO DIRETTO DI SEGNALI CHIMICI E ELETTRICI ATTRAVERSO GIUNZIONI COMUNICANTI COMUNICAZIONE CHIMICA LOCALE (SOSTANZE PARACRINE E AUTOCRINE) COMUNICAZIONE A LUNGA DISTANZA
Comunicare. 6. La comunicazione cellulare. Messaggio EFFETTO. Recepito e elaborato
 6. La comunicazione cellulare contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale Comunicare Messaggio Recepito e elaborato
6. La comunicazione cellulare contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale Comunicare Messaggio Recepito e elaborato
TEST NEUROPSICOBIOLOGIA 2010
 TEST NEUROPSICOBIOLOGIA 2010 1. Il sistema renina-angiotensina è uno dei principali sistemi regolatori dell'apparato: a. Cardio-vascolare b. Dalle gonadi c. Cardio-vascolare e del rene d. Del rene 2. Il
TEST NEUROPSICOBIOLOGIA 2010 1. Il sistema renina-angiotensina è uno dei principali sistemi regolatori dell'apparato: a. Cardio-vascolare b. Dalle gonadi c. Cardio-vascolare e del rene d. Del rene 2. Il
SISTEMA NEUROENDOCRINO
 Mantenimento dell omeostasi Nervoso: risposta rapida +! Endocrino: risposta lenta SISTEMA NEUROENDOCRINO Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato
Mantenimento dell omeostasi Nervoso: risposta rapida +! Endocrino: risposta lenta SISTEMA NEUROENDOCRINO Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato
sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio in senso lato sinapsi neurone effettori elettriche chimiche
 sinapsi 1 sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio neurone in senso lato effettori cellule muscolari, gh. esocrine, gh. endocrine sinapsi elettriche chimiche 2 sinapsi
sinapsi 1 sinapsi punto in cui il neurone entra in contatto con la sua cellula bersaglio neurone in senso lato effettori cellule muscolari, gh. esocrine, gh. endocrine sinapsi elettriche chimiche 2 sinapsi
Ruolo della membrana plasmatica: - nelle comunicazioni intercellulari. - nella trasduzione del segnale extracellulare
 Ruolo della membrana plasmatica: - nelle comunicazioni intercellulari - nella trasduzione del segnale extracellulare Tra cellule vicine, la comunicazione può avvenire con diverse modalità. Pure la comunicazione
Ruolo della membrana plasmatica: - nelle comunicazioni intercellulari - nella trasduzione del segnale extracellulare Tra cellule vicine, la comunicazione può avvenire con diverse modalità. Pure la comunicazione
LEZIONE 1_TRASPORTI DI MEMBRANA
 LEZIONE 1_TRASPORTI DI MEMBRANA 1 2 I TRASPORTI DI MEMBRANA Diffusione semplice Diffusione facilitata Trasporto attivo 3 La diffusione semplice Lezione_1_trasporti 4 La diffusione semplice Relazione tra
LEZIONE 1_TRASPORTI DI MEMBRANA 1 2 I TRASPORTI DI MEMBRANA Diffusione semplice Diffusione facilitata Trasporto attivo 3 La diffusione semplice Lezione_1_trasporti 4 La diffusione semplice Relazione tra
Comunicazione cellulare
 Comunicazione cellulare Per segnali cellulari (cell signaling) si può intendere sia una catena di segnali tutti all interno di una cellula sia la catena di eventi che consente ad un segnale extracellulare
Comunicazione cellulare Per segnali cellulari (cell signaling) si può intendere sia una catena di segnali tutti all interno di una cellula sia la catena di eventi che consente ad un segnale extracellulare
I recettori di membrana e la trasduzione del segnale
 I recettori di membrana e la trasduzione del segnale I recettori di membrana recepiscono segnali fisici o chimici provenienti dall esterno della cellula fotoni / vibrazioni (recepiti dalle rodopsine nella
I recettori di membrana e la trasduzione del segnale I recettori di membrana recepiscono segnali fisici o chimici provenienti dall esterno della cellula fotoni / vibrazioni (recepiti dalle rodopsine nella
Segnalazione cellulare e trasduzione del segnale. Comunicazione fra le cellule
 Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce
Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce
scaricato da www.sunhope.it
 Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Corso di Fisica Medica 1
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Conduzione nervosa 30/3/2006 Terminazioni pre-sinaptiche Mitocondri Sintesi dei neuro trasmettitori (NT) Attività enzimatica Trasporto
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Conduzione nervosa 30/3/2006 Terminazioni pre-sinaptiche Mitocondri Sintesi dei neuro trasmettitori (NT) Attività enzimatica Trasporto
DOGMA CENTRALE DELLA BIOLOGIA. Secondo il dogma centrale della biologia, il DNA dirige la. sintesi del RNA che a sua volta guida la sintesi delle
 DOGMA CENTRALE DELLA BIOLOGIA Secondo il dogma centrale della biologia, il DNA dirige la sintesi del RNA che a sua volta guida la sintesi delle proteine. Tuttavia il flusso unidirezionale di informazioni
DOGMA CENTRALE DELLA BIOLOGIA Secondo il dogma centrale della biologia, il DNA dirige la sintesi del RNA che a sua volta guida la sintesi delle proteine. Tuttavia il flusso unidirezionale di informazioni
 TRASDUZIONE DEL SEGNALE LE PRINCIPALI MOLECOLE CHE PROVVEDONO ALLO SCAMBIO DI INFORMAZIONI TRA LE CELLULE SONO FATTORI SOLUBILI QUALI NEUROTRASMETTITORI, ORMONI, FATTORI DI CRESCITA, CITOCHINE. UN CASO
TRASDUZIONE DEL SEGNALE LE PRINCIPALI MOLECOLE CHE PROVVEDONO ALLO SCAMBIO DI INFORMAZIONI TRA LE CELLULE SONO FATTORI SOLUBILI QUALI NEUROTRASMETTITORI, ORMONI, FATTORI DI CRESCITA, CITOCHINE. UN CASO
Scaricato da 1
 Muscolo cardiaco scheletrico liscio Eterogeneità: muscolo liscio vascolare gastrointestinale urinario respiratorio riproduttivo oculare Anatomia: le fibre contrattili del muscolo liscio sono disposte in
Muscolo cardiaco scheletrico liscio Eterogeneità: muscolo liscio vascolare gastrointestinale urinario respiratorio riproduttivo oculare Anatomia: le fibre contrattili del muscolo liscio sono disposte in
Tirosin chinasi senza o con funzione recettoriale
 Tirosin chinasi senza o con funzione recettoriale elica di poliprolina Il dominio chinasico di Src è mantenuto nello stato inattivo mediante due distinte interazioni intramolecolari: il legame del dominio
Tirosin chinasi senza o con funzione recettoriale elica di poliprolina Il dominio chinasico di Src è mantenuto nello stato inattivo mediante due distinte interazioni intramolecolari: il legame del dominio
1 PARTE: Recettori accoppiati alle proteine G
 1 PARTE: Recettori accoppiati alle proteine G Circa l 80% degli ormoni, dei neurotrasmettitori e dei neuromodulatori regolano le interazioni cellulari 1. Essi sono i cosiddetti primi messaggeri, che inducono
1 PARTE: Recettori accoppiati alle proteine G Circa l 80% degli ormoni, dei neurotrasmettitori e dei neuromodulatori regolano le interazioni cellulari 1. Essi sono i cosiddetti primi messaggeri, che inducono
Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente WWW.SUNHOPE.IT Tubo neurale WWW.SUNHOPE.IT Ventricoli
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I Sistema nervoso: organizzazione, SNC, sistema periferico afferente ed efferente WWW.SUNHOPE.IT Tubo neurale WWW.SUNHOPE.IT Ventricoli
Cambiamenti durante l esercizio: a mantenere l equilibrio delle funzioni del corpo: OMEOSTASI
 Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della frequenza respiratoria a Aumento della frequenza cardiaca Aumento
Sistema Neuro-endocrino Cambiamenti durante l esercizio: Contrazione dei muscoli sistema nervoso attivato coscientemente Aumento della frequenza respiratoria a Aumento della frequenza cardiaca Aumento
Regolazione dell espressione genica
 Regolazione dell espressione genica Ogni processo che porta dal DNA alla proteina funzionale è soggetto a regolazione. Il primo processo relativo alla trascrizione è il più importante punto di regolazione
Regolazione dell espressione genica Ogni processo che porta dal DNA alla proteina funzionale è soggetto a regolazione. Il primo processo relativo alla trascrizione è il più importante punto di regolazione
Tutor Pietro Bortoluzzi Cell
 Prof.ssa Paola Guandalini ---------------------------------------------------- Università degli Studi di Ferrara Dipartimento di Scienze Biomediche e Chirurgico specialistiche Sezione di Fisiologia Umana
Prof.ssa Paola Guandalini ---------------------------------------------------- Università degli Studi di Ferrara Dipartimento di Scienze Biomediche e Chirurgico specialistiche Sezione di Fisiologia Umana
Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI
 Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI Il percorso di maturazione dei linfociti Sviluppo della specicifità immunologica I linfociti B e T avviano le risposte immunitarie dopo il
Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI Il percorso di maturazione dei linfociti Sviluppo della specicifità immunologica I linfociti B e T avviano le risposte immunitarie dopo il
La trasduzione del segnale intracellulare
 La trasduzione del segnale intracellulare Chemical signaling mechanisms Amplification in signal transduction pathways Un aspetto importante è il controllo temporale del comportamento cellulare: il trasferimento
La trasduzione del segnale intracellulare Chemical signaling mechanisms Amplification in signal transduction pathways Un aspetto importante è il controllo temporale del comportamento cellulare: il trasferimento
Ioni fosfato e proteine (anioni) sono prevalenti nella cellula
 sono prevalenti nella cellula") Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
Equilibrio osmotico tra liquido intracellulare e liquido extracellulare (e plasma) mm Na + K + Cl - HCO 3 - grandi anioni proteine Liquido intracellulare Liquido extracellulare 10 140 140 5 5 35 10 35
MUSCOLO. Muscolo striato scheletrico. FGE aa
 MUSCOLO Muscolo striato scheletrico FGE aa.2015-16 Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli
MUSCOLO Muscolo striato scheletrico FGE aa.2015-16 Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli Muscolo scheletrico Muscolo liscio Muscolo cardiaco Tipi di muscoli