Glicogeno, amido, saccarosio. Glucosio
|
|
|
- Rosina Santini
- 10 anni fa
- Просмотров:
Транскрипт
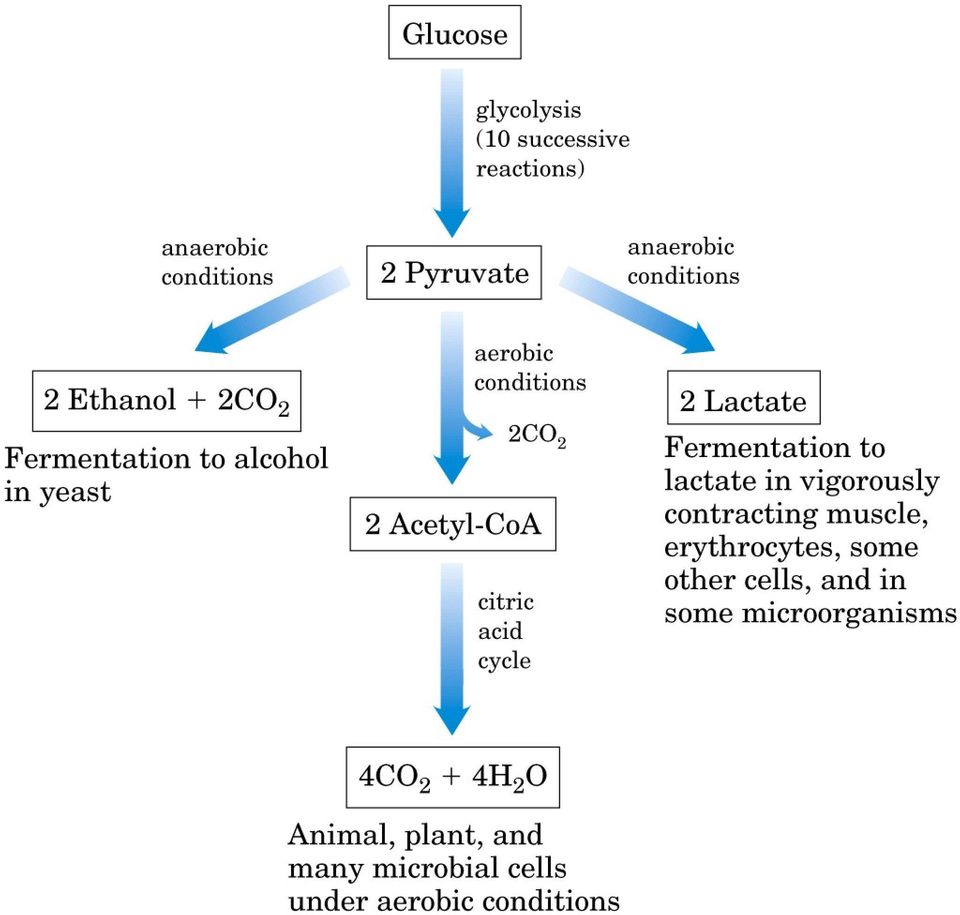
1 Glicogeno, amido, saccarosio Riserva Ossidazione mediante la via del pentosio fosfato Glucosio Ossidazione mediante la glicolisi Ribosio 5-fosfato5 Piruvato

2
3
4 Fase preparatoria

5 Fase di recupero energetico

6 Tutti gli intermedi glicolitici sono fosforilati. 1) Gli intermedi fosforilati non possono uscire dalla cellula. 2) I composti fosforilati ad alta energia che si formano nel corso della glicolisi (1,3-bisfosfoglicerato e fosfoenolpiruvato) ) donano i loro gruppi fosforici all ADP e formano ATP. 2) Il gruppo fosforico favorisce il legame del substrato all enzima.
 ) donano i loro gruppi fosforici all ADP e")
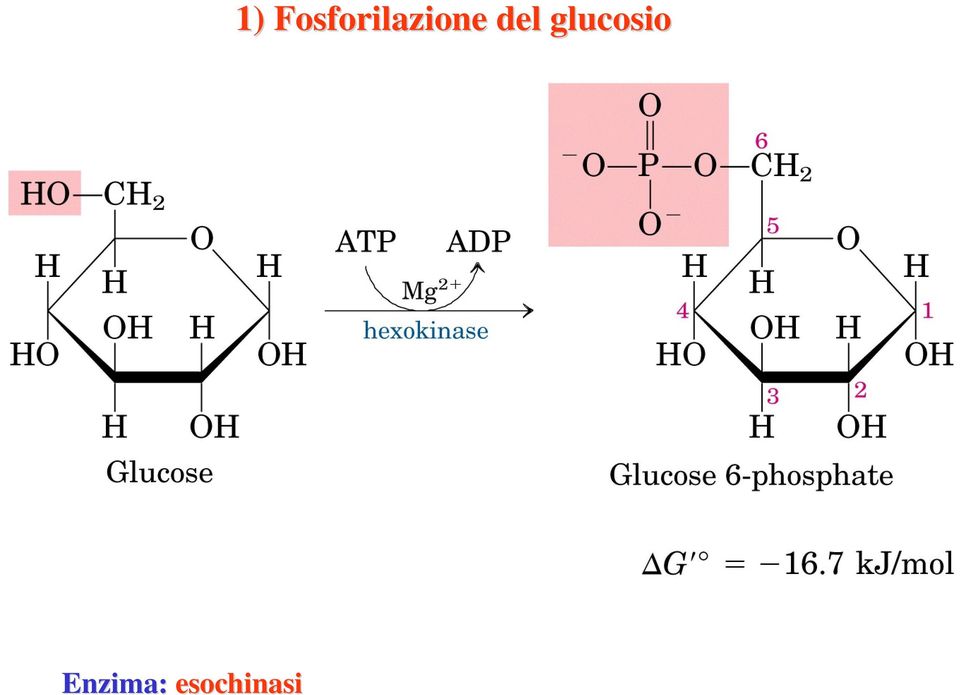
7 Enzima: esochinasi 1) Fosforilazione del glucosio
8 I substrati di quasi tutti gli enzimi NTP-dipendenti sono i complessi formati da magnesio (o manganese) e nucleoside trifosfato Lo ione magnesio neutralizza alcune cariche negative del NTP, riducendo le interazioni ioniche non specifiche tra l enzima l e il gruppo polifosforico del nucleotide Lo ione magnesio mantiene il nucleotide in una conformazione ben definita che può essere riconosciuta dall enzima. Lo ione magnesio fornisce ulteriori punti di interazione tra il complesso ATP- Mg 2+ e l enzima. l

9 Enzima

10 Adattamento indotto dell esochinasi

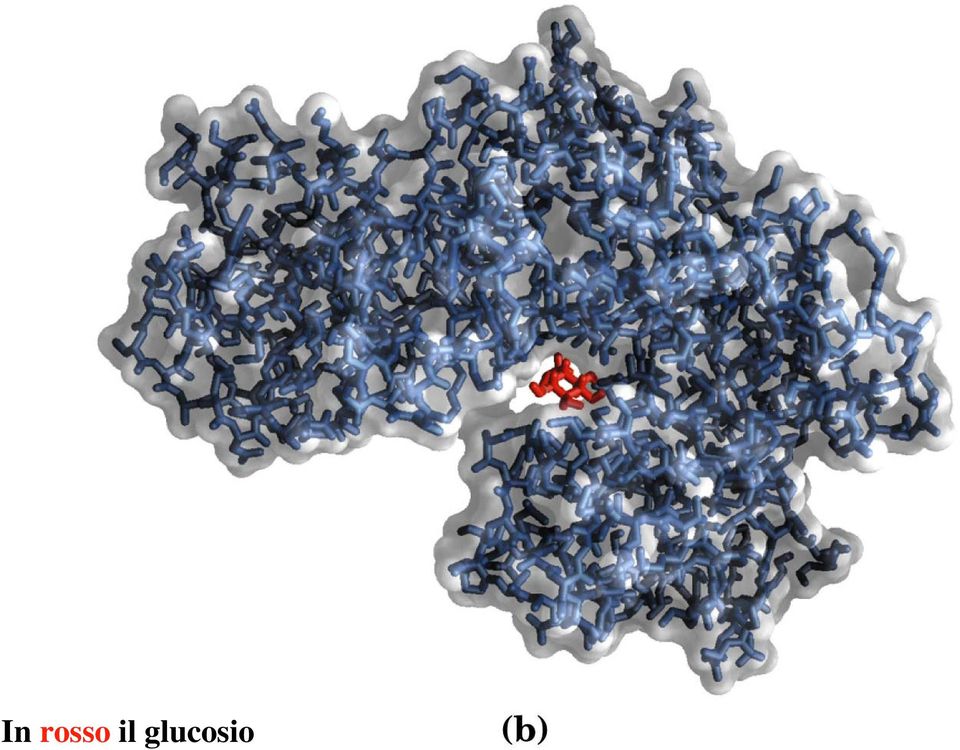
11 In rosso il glucosio
12 Adattamento indotto nell esochinasi esochinasi Le modificazioni conformazionali indotte dal legame del substrato creano un ambiente molto più apolare intorno al glucosio il che favorisce la donazione del gruppo fosforico terminale dell ATP. L adattamento indotto assicura che si formi una conformazione cataliticamente attiva dell enzima enzima solo quando sono legati sia il donatore che l accettorel del fofosfato, impedendo trasferimenti dispendiosi del gruppo fosforico all acqua acqua

13 2) Conversione del glucosio-6-fosfato in fruttosio 6-fosfato6 Enzima: fosfoesosio isomerasi o fosfoglucosio isomerasi

14 Il glucosio-6-fosfato, un aldoso,, viene trasformato in fruttosio-6-fosfato, fosfato, un chetoso.

15 3) Fosforilazione del fruttosio-6-fosfato fosfato a fruttosio-1,6 1,6-bisfosfato Enzima: fosfofruttochinasi 1

16 Formazione della gliceraldeide 3-fosfato per scissione (4) e isomerizzazione (5) Enzima: aldolasi
")
17 Rottura del fruttosio 1,6-bisfosfato
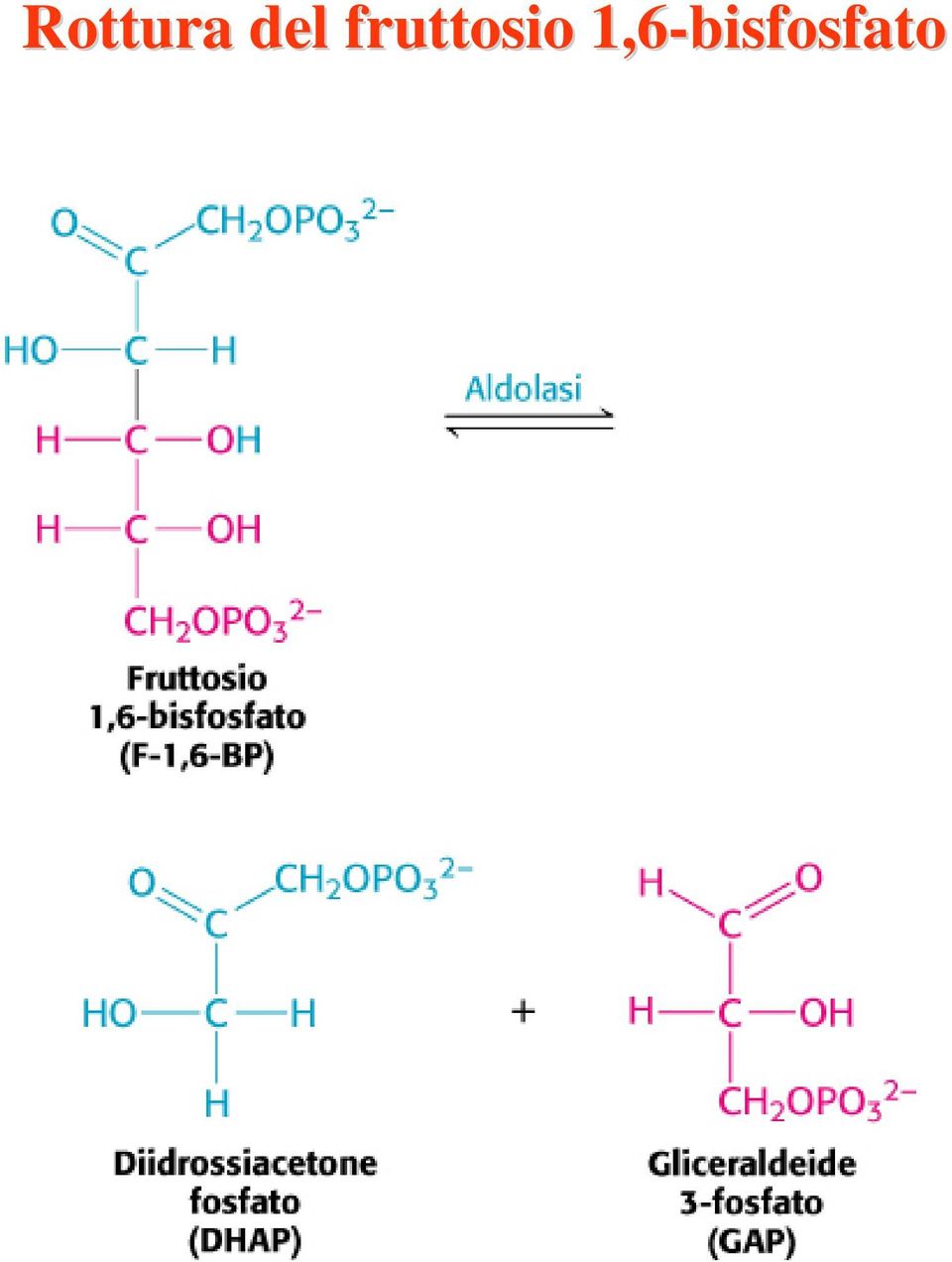
18 Scissione del fruttosio 1,6-bisfosfato in glicaraldeide 3-fosfato e diidrossiacetone fosfato. I prodotti delle restanti tappe della glicolisi sono costituiti da unità a tre atomi di carbonio anziché da unità a sei atomi di carbonio. L enzima aldolasi è così chiamato in base alla natura della reazione inversa, una condensazione aldolica. DHAP= diidrossiacetone fosfato GAP= gliceraldeide 3-fosfato

19 5) Interconversione dei trioso fosfato Enzima: trioso fosfato isomerasi

20 Struttura della trioso fosfato isomerasi 8 filamenti β circondati da 8 α eliche: il barile αβ

21 Meccanismo catalitico della trioso fosfato isomerasi Il glutammato 165 trasferisce un protone tra gli atomi 1 e 2 con l ausilio l dell istidina 95. L istidina 95 facilita la catalisi donando un protone per stabilizzare la carica negativa che si sviluppa sul gruppo carbonilico sull atomo C-2. C La forma rara dell istidina carica negativamente è stabilizzata da interazioni con altre parti dell enzima enzima
22 Struttura della trioso fosfato isomerasi L ansa impedisce all intermedio instabile enediolo di abbandonare l enzima l chiudendo il sito attivo sino a quando l isomerizzazione l non è completata
23
24 Gli atomi C 1, C 2 e C 3 diventano indistinguibili rispettivamente dagli atomi C 6, C 5 e C 4.
25 6) Ossidazione della gliceraldeide 3-fosfato a 1,3-bisfosfoglicerato Enzima: Gliceraldeide 3-fosfato deidrogenasi La fosforilazione è accoppiata all ossidazione della gliceraldeide 3-fosfato
26 La reazione catalizzata dalla gliceraldeide 3-fosfato deidrogenasi è la somma di due reazioni: 1) Ossidazione dell aldeide a un acido carbossilico ad opera del NAD + (termodinamicamente favorevole) 2) Unione dell acido carbossilico e dell ortofosfato a formare un acilfosfato (termodinamicamente sfavorevole) Se queste reazioni si svolgessero in successione la seconda avrebbe un energia di attivazione troppo alta per avvenire a velocità significativa. I due processi devono essere accoppiati.
27 Struttura della gliceraldeide 3-fosfato deidrogenasi Il sito attivo include un residuo di cisteina e un residuo di istidina adiacenti a un NAD + legato
28 Meccanismo catalitico della gliceraledeide 3-fosfato deidrogenasi La reazione procede attraverso un intermedio tioestere,, che permette che l ossidazione della gliceraldeide sia accoppiata alla fosforilazione del 3-3 fosfoglicerato
29 L intermedio tioestere ha un energia libera più alta di quella dell acido carbossilico libero Reazione non accoppiata Reazione accoppiata
30 Reazione catalizzata dalla gliceraldeide 3-fosfato deidrogenasi
31 Lo iodoacetato è un potente inibitore della gliceraldeide 3- fosfato deidrogenasi
32 7) Formazione dell ATP dall 1,3-bisfosfoglicerato Enzima: Fosfoglicerato chinasi
33 Idrolisi dell 1,3-bisfosfoglicerato Il prodotto dell idrolisi è il 3-fosfoglicerato 3 il cui gruppo carbossilico, dopo la formazione, può andare incontro a dissociazione. La ionizzazione e le strutture di risonanza rendono possibile la stabilizzazione dei prodotti della reazione idrolitica. Un ulteriore contributo alla variazione di energia libera è dovuto alla stabilizzazione per risonanza del P i.
34 .formazione del piruvato e generazione della seconda molecola di ATP (tappe 8, 9 e 10).
35 8) Conversione del 3-fosfoglicerato 3 in 2-fosfoglicerato2 Enzima: Fosfoglicerato mutasi
36 Meccanismo catalitico della fosfoglicerato mutasi L enzima viene inizialmente fosforilato mediante trasferimento di un gruppo fosforico dal 2,3 BPG.. Il 2,3 BPG si comporta da cofattore: è necessario in piccole quantità per iniziare il ciclo e poi viene continuamente rigenerato dal ciclo stesso. Solo negli eritrociti il 2,3 BPG è presente in alte concentrazioni (5 mm) ) e regola l affinitl affinità dell emoglobina emoglobina per l ossigeno l Sono necessarie quantità catalitiche di 2,3-bisfosfoglicerato per mantenere un residuo di istidina del sito attivo in forma fosforilata
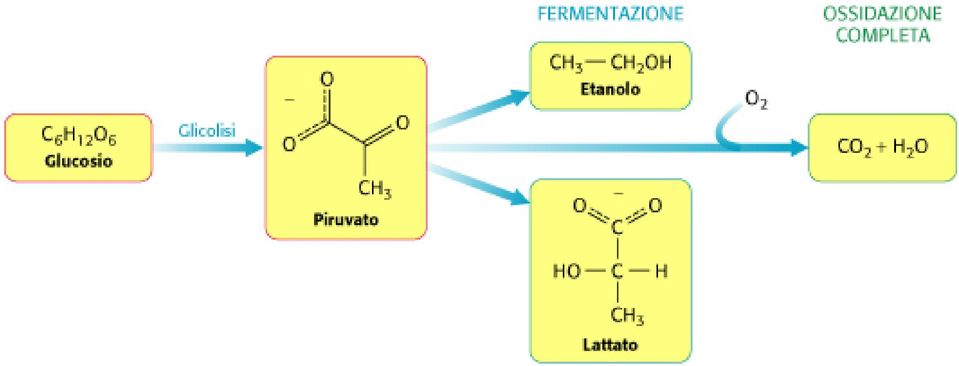
37 9) Deidratazione del 2-fosfoglicerato 2 a fosfoenolpiruvato Enzima: Enolasi Si forma un enolo. Un enolfosfato ha un elevato potenziale di trasferimento del gruppo fosforico; Un estere fosforico di un normale alcol ha un potenziale di trasferimento del gruppo fosforico basso.
38 Idrolisi del fosfoenolpiruvato Piruvato (forma enolica) Piruvato (forma chetonica) Il prodotto dell idrolisi può esistere in due forme tautomeriche, mentre il reagente ha una sola forma, la forma enolica. Il prodotto è più stabile del reagente.
39 10) Trasferimento del gruppo fosforico dal fosfoenolpiruvato all ADP (formazione della seconda molecola di ATP).
40 Tautomerizzazione del piruvato
41 Consumo e produzione di ATP nella glicolisi Reazione Molecole di ATP Glucosio Glucosio 6-fosfato6-1 Fruttosio 6-fosfato 6 Fruttosio 1,6-bisfosfato x 1,3-bisfosfoglicerato 2 x 3-fosfoglicerato x Fosfoenolpiruvato 2 x piruvato + 2 Resa complessiva + 2
42
43 Glicolisi Glucosio + 2 ATP + 2 NAD ADP + 2 P i 2 Piruvato + 2 ADP + 2 NADH + 2 H ATP + 2 H 2 O semplificando i termini comuni Glucosio + 2 NAD ADP + 2 P i 2 Piruvato + 2 NADH + 2 H ATP + 2 H 2 O ΔG o = - 85 kj/mole
44 Gli intermedi glicolitici sono incanalati fra i siti attivi degli enzimi Gli enzimi della glicolisi vengono considerati normalmente componenti solubili nel citosol delle cellule, ma vi sono numerosi risultati sperimentali che tendono t a suggerire che questi enzimi all interno della cellula esistano come complessi multienzimatici. Quando una cellula viene aperta, per purificare gli enzimi citosolici,, la diluizione della soluzione del citosol contenente complessi proteici non covalenti favorisce la dissociazione dei complessi nei suoi costituenti.
45 Incanalamento di un substrato tra due enzimi della via glicolitica
46 Differenti destini del piruvato. Si forma etanolo a partire dal piruvato nei lieviti e in parecchi altri microorganismi Si forma lattato a partire dal piruvato in un ampia gamma di microorganismi,, ed anche nelle cellule degli organismi superiori quando la quantità di ossigeno è limitante come nel tessuto muscolare durante l attivitl attività fisica intensa Una quantità di energia molto maggiore può essere estratta aerobicamente mediante il ciclo dell acido citrico e la catena di trasporto degli elettroni
47 La formazione e il consumo di NADH, localizzati entro la via glicolitica.
48 La fermentazione indica la degradazione anaerobica del glucosio (e di altri nutrienti organici) per ottenere energia sotto forma di ATP.
49 La fermentazione lattica Nella fermentazione lattica (o lattacida) ) il NAD + viene rigenerato dal NADH mediante la riduzione del piruvato a lattato.
50
51 La fermentazione alcolica I lieviti ed alcuni microorganismi fermentano il glucosio ad etanolo e CO 2.
52 La tiamina pirofosfato (TPP) e il suo ruolo nella decarbossilazione del piruvato Protone acido, la sua perdita genera un carbanione La tiamina pirofosfato è la forma coenzimatica della vitamina B 1 (tiamina). La TPP ha una funzione rilevante nelle reazioni in cui si ha la rottura di un legame adiacente a un gruppo carbonilico.
53 La tiamina pirofosfato (TPP) e il suo ruolo nella decarbossilazione del piruvato
54 La tiamina pirofosfato (TPP) e il suo ruolo nella decarebossilazione del piruvato 1) Il carbanione della TPP agisce come un nucleofilo,, attaccando il carbonio carbonilico del piruvato 2) In seguito alla decarbossilazione si forma un carboanione.. L anello L tiazolico della TPP si comporta come una trappola per elettroni, stabilizzando il carbanione per risonanza Dopo la protonazione (3), il prodotto della reazione, l acetaldeide, viene rilasciato (4) e un protone si dissocia per rigenerare il carbanione della TPP (5)
55 Alcune reazioni in cui la tiamina pirofosfato è un cofattore essenziale
56
57 Il sito attivo dell acol deidrogenasi Il sito attivo contiene uno ione zinco legato a due residui di cisteina e a un residuo di istidina. Lo ione zinco lega il substrato acetaldeidico attraverso il suo atomo di ossigeno, polarizzandolo in modo che possa accettare più facilmente un idruro dal NADH
58 Una fermentazione su scala industriale
59 Mantenimento dell equilibrio equilibrio redox
60 Vie di alimentazione della glicolisi
61 Rimozione del residuo di glucosio terminale dall estremit estremità non riducente di una catena di glicogeno ad opera della glicogeno fosforilasi.
62 La glicogeno fosforilasi catalizza la reazione in cui il legame glicosidico (α1 4), viene attaccato dal fosfato inorganico producendo il rilascio o di glucosio 1-fosfato1 fosfato. Parte dell energia energia del legame glicosidico viene conservata mediante la formazione dell estere estere fosforico nel glucosio 1-fosfato. 1
63 Demolizione del glicogeno vicino a un punto di ramificazione (α1 6)( Dopo la rimozione sequenziale dei residui terminali di glucosio da parte della glicogeno fosforilasi,, i residui di glucosio vicini a una ramificazione sono staccati mediante un processo a due tappe che richiede l azione l di un enzima deramificate bifunzionale.
64 Il glucosio 1-fosfato1 viene poi convertito in glucosio 6-fosfato 6 dall enzima fosfoglucomutasi. Glucosio 1-fosfato 1 Glucosio 6-fosfato6 Nel fegato è presente l enzima l Glucosio 6-fosfatasi6 che catalizza la reazione Glucosio 6-fosfato 6 + H 2 O Glucosio + P i
65 La maggior parte del fruttosio viene metabolizzato dal fegato usando la via del fruttosio 1, fosfato. Fruttosio + ATP Fruttosio 1-fosfato 1 + ADP enzima: fruttochinasi - Il fruttosio 1-fosfato1 viene quindi convertito in gliceraldeide e diidrossiacetone fosfato dalla fruttosio 1-fosfato 1 aldolasi. - Il diidrossiacetone fosfato viene poi convertito in gliceraldeide 3-fosfato dall enzima glicolitico trioso fosfato isomerasi. - La gliceraldeide viene poi trasformata dalla trioso chinasi: Gliceraldeide + ATP Gliceraldeide 3-fosfato + ADP.
66 Il fruttosio può anche essere fosforilato a fruttosio 6-fosfato6 dall esochinasi esochinasi. Nel muscolo, nel tessuto adiposo e nel rene questa è la via principale di utilizzo del fruttosio.
67 Mannosio enzima: esochinasi Mannosio + ATP Mannosio 6-fosfato + ADP Il Mannosio 6-fosfato viene poi isomerizzato ad opera della fosfomannosio isomerasi in fruttosio 6-fosfato. 6
68 Via di conversione del D-Galattosio 1-fosfato 1 in D-Glucosio 1-fosfato1
69 I disaccaridi della dieta vengono idrolizzati a monosaccaridi enzima: maltasi Maltosio + H 2 O 2 D-GlucosioD enzima: lattasi Lattosio + H 2 O D-Galattosio + D-GlucosioD enzima: saccarasi Saccarosio + H 2 O D-Fruttosio + D-GlucosioD enzima: trealasi Trealosio + H 2 O 2 D-GlucosioD
70 L intolleranza al lattosio La lattasi nell endotelio endotelio intestinale: Sebbene tutti i bambini possiedano una lattasi intestinale, molti i di loro smettono di produrre questo enzima da adulti diventando intolleranti al lattosio. l La lattasi può essere identificata trattando una fettina di tessuto intestinale con un anticorpo che si lega in modo specifico a questo enzima. a) Adulto che ha conservato alti livelli di lattasi b) Adulto con un intolleranza al lattosioi
71 Vie di alimentazione della glicolisi
72
73 Cancro e glicolisi Le condizioni ipossiche esistenti all interno di una massa tumorale determinano l attivazione l del fattore di trascrizione inducibile dall ipossia (HIF-1), che induce l adattamento l metabolico (aumento degli enzimi glicolitici) e attiva i fattori angiogenici che stimolano la crescita di nuovi vasi sanguigni.
74 Regolazione del catabolismo dei carboidrati
75 In ogni via metabolica vi è almeno una reazione che è lontana dall equilibrio a causa della bassa attività dell enzima enzima cha la catalizza. La velocità di questa reazione non è limitata dalla disponibilità del substrato, ma soltanto dall attivit attività dell enzima. enzima. La reazione viene detta limitata dall enzima e la velocità di questa tappa determina quella della via nel suo complesso. In genere queste tappe che limitano la velocità sono reazioni molto esoergoniche e quindi irreversibili nelle condizioni intracellulari. Gli enzimi che catalizzano queste tappe esoergoniche che limitano la velocità sono di solito il bersaglio delle regolazioni metaboliche.
76 Glicolisi e gluconeogenesi sono regolate in modo coordinato PFK-1 = fosfofruttochinasi -1; FBPase-1 = fruttosio 1,6-bisfosfatasi. Tre reazioni della glicolisi sono così esoergoniche da essere praticamente irreversibili; sono quelle catalizzate dalla esochinasi,, dalla PFK-1 e dalla piruvato chinasi.
77 L esochinasi del muscolo viene inibita allostericamente dal suo prodotto, il glucosio 6-6 fosfato I mammiferi hanno diverse forme di esochinasi,, tutte capaci di catalizzare la conversione del glucosio in glucosio 6-fosfato. 6 Proteine diverse in grado di catalizzare la stessa reazione sono dette isoenzimi. La forma predominante di esochinasi nel fegato è l esochinasi D, detta anche glucochinasi La glucochinasi continua a fosforilare il glucosio anche se i livelli di glucosio 6-fosfato 6 sono elevati. La glucochinasi ha una elevata K M per il glucosio e diventa efficiente quando il glucosio è abbondante. La funzione della glucochinasi epatica è quella di fornire glucosio 6-fosfato 6 per la sintesi del glicogeno. Il glucosio 6-fosfato 6 non è solo un intermedio della glicolisi ( glicogeno, via dei pentosio-fosfato fosfato). La prima reazione irreversibile utilizzata dalla sola via glicolitica è la fosforilazione del fruttosio 6-fosfato 6 a fruttosio 1,6-bisfosfato. La fosfofruttochinasi-1 è il principale sito di controllo della glicolisi
78 La fosfofruttochinasi - 1 è il principale elemento di regolazione della via glicolitica nei mammiferi. L enzima del fegato viene inibito da elevate concentrazioni di ATP.
79 Struttura della fosfofruttochinasi 1
80 Elevate concentrazioni di ATP fanno diminuire l affinitl affinità dell enzima enzima per il substrato fruttosio 6-fosfato. 6 L azione inibitrice dell ATP viene abolita dall AMP. Perché l AMP e non l ADP? l Quando l ATP l viene utilizzato rapidamente, l adenilatol chinasi può formare ATP a partire da ADP: ADP + ADP ATP + AMP L AMP è il segnale per lo stato a bassa energia.
81 -Elevate concentrazioni di ATP fanno diminuire l affinitl affinità dell enzima enzima per il substrato fruttosio 6-fosfato. 6 -L azione inibitrice dell ATP viene abolita dall AMP. -La fosfofruttochinasi -1 è inibita dal citrato,, il primo intermedio del ciclo dell acido citrico. Elevati livelli di citrato indicano che le necessità energetiche e la richiesta di metaboliti per le vie biosintetiche sono soddisfatte.
82
83 Il principale regolatore allosterico della fosfofruttochinasi-1 1 (PFK-1) è il fruttosio 2,6 bisfosfato. Il fruttosio 2,6 bisfosfato è un potente attivatore della PFK-1. Il fruttosio 2,6 bisfosfato si forma dal fruttosio 6-fosfato 6 ad opera della fosfofruttochinasi 2 (PFK-2) Il fruttosio 2,6 bisfosfato viene idrolizzato a fruttosio 6-fosfato 6 da una specifica fosfatasi la fruttosio 2,6 bisfosfatasi (Fbasi-2)
84 La fosfofruttochinasi 2 e la fruttosio 2,6 bisfosfatasi sono presenti in una unica catena polipeptidica (detta enzima tandem) ) e sono reciprocamente regolate dalla fosforilazione di un residuo di serina. Proteina fosforilata attività fosfatasica Proteina defosforilata attività chinasica
85 Regolazione della sintesi e della degradazione del fruttosio 2,6-bisfosfato Una diminuzione della concentrazione del glucosio ematico,, segnalato dal glucagone, determina la fosforilazione dell enzima enzima bifunzionale (aumento attività fosfatasica) ) e quindi fa diminuire la concentrazione di fruttosio 2,6-bisfosfato, rallentando la glicolisi. Un aumento della concentrazione di fruttosio 6-fosfato6 fosfato,, che segue ad un aumento del glucosio ematico, accellera la formazione di fruttosio 2,6-bisfosfato facilitando la defosforilazione dell enzima enzima (aumento( dell attivit attività chinasica).
86 La piruvato chinasi viene inibita da concentrazioni elevate di ATP L ATP inibisce la piruvato chinasi allostericamente,, diminuendo l affinitl affinità dell enzima enzima per il suo substrato, il fosfoenolpiruvato. La piruvato chinasi viene anche inibita dall Acetil Acetil-CoA e dagli acidi grassi a catena lunga,, importanti combustibili per il ciclo dell acido citrico.
87 Regolazione della glicogeno fosforilasi La glicogeno fosforilasi è regolata da più effettori allosterici che segnalano lo stato energetico della cellula nonché dalla fosforilazione reversibile che risponde a segnali ormonali quali l insulina, l il glucagone e l adrenalina. l
88 Strutture della fosforilasi a e della fosforilasi b La glicogeno fosforilasi a è fosforilata sulla serina 14 di ciascuna subunità. Questa modificazione favorisce la struttura dello stato R più attivo.
89
90
91
92 Attivazione della fosforilasi chinasi
93 Nel muscolo il fine ultimo della glicolisi è la produzione di ATP Il fegato ha il ruolo di mantenere costante il livello di glucosio nel sangue producendo ed esportando questo metabolita ai tessuti che lo richiedono, e importandolo e conservandolo quando viene introdotto in eccesso con la dieta.
94 Muscolo La fosforilasi b muscolare è attiva in presenza di elevate concentrazioni di AMP,, che si lega a un sito di legame per i nucleotidi presente su entrambe le subunità e stabilizza la conformazione della fosforilasi b nello stato R.
95 La glicogeno fosforilasi del fegato è un sensore per il glucosio. Il legame del glucosio sposta l equilibrio allosterico della glicogeno fosforilasi a epatica dallo stato R (attivo) allo stato T (meno attivo). Se è presente glucosio libero derivante da qualche altra fonte come la dieta, non è necessario mobilitare il glicogeno.
96 Il metabolismo secondario del glucosio Vie cataboliche che, partendo dal glucosio, portano alla formazione di prodotti specializzati necessari alla cellula.
97 La via del pentoso fosfato - produce NAPDH e Ribosio 5-fosfato5 fosfato. Glucosio 6-fosfato NADP + + H 2 O Ribosio 5-fosfato 5 + CO NADPH + 2 H + Il NAPDH è necessario per le reazioni di biosintesi Il Ribosio 5-fosfato5 è un precursore nella sintesi dei nucleotidi (presente in: ATP, CoA, NAD +, FAD, DNA, RNA)
98 I trasportatori di elettroni derivati dalla nicotinamide Il NADH viene ossidato dalla catena respiratoria per generare ATP Il NADPH serve come donatore di elettroni nelle biosintesi riduttive
99 Le reazioni ossidative della via del pentoso fosfato (a)
100 Le reazioni ossidative della via del pentoso fosfato (b)
101 La via del pentoso fosfato è molto attiva nel tessuto adiposo in cui è attiva la biosintesi degli acidi grassi che richiede potere riducente nella a forma di NADPH. L attività della via del pentoso fosfato è molto bassa nel muscolo scheletrico.
102 La via del pentoso fosfato Reazione Enzima Parte ossidativa Glucosio 6-fosfato 6 + NADP + 6-fosfoglucono-δ-lattone lattone + NADPH + H + Glucosio 6-fosfato6 deidrogenasi 6-fosfoglucono-δ-lattone lattone + H 2 O 6-fosfogluconato + H + Lattonasi 6-fosfogluconato + NADP + ribuloso 5-fosfato + CO 2 + NADPH 6-fosfogluconato deidrogenasi
103 La via del pentoso fosfato Nei tessuti che richiedono più NADPH per le biosintesi riduttive che ribosio 5-5 fosfato da incorporare nei nucleotidi,, il ribosio 5-fosfato 5 viene convertito in gliceraldeide 3-fosfato e fruttosio 6-fosfato6 (reazioni non ossidative della via del pentoso fosfato). 3 ribosio 5-fosfato 5 2 fruttosio 6-fosfato 6 + gliceraldeide 3-fosfato Il ribosio 5-fosfato 5 formato in eccesso dalla via del pentoso fosfato può essere completamente convertito in intermedi glicolitici. Il fruttosio 6-fosfato 6 fosfato e la gliceraldeide 3-fosfato sono anche intermedi della via gluconeogenetica.
104 La via del pentoso fosfato Reazione Enzima Parte non ossidativa riboso 5-fosfato ribuloso 5-fosfato Fosfopentoso isomerasi ribuloso 5-fosfato xiluloso 5-fosfato Fosfopentoso epimerasi xiluloso 5-fosfato + riboso 5-fosfato sedoeptulosio 7-fosfato + gliceraldeise 3-fosfato Transchetolasi sedoeptulosio 7-fosfato + gliceraldeise 3-fosfato fruttoso 6-fosfato + eritroso 4-fosfato Transaldolasi Xiluloso 5-fosfato + eritroso 4-.fosfato fruttoso 6-fosfato + glicaraldeide 3-fosfato Transchetolasi
105 Parte non ossidativa della via del pentoso fosfato transchetolasi C 5 + C 5 C 3 + C 7 transaldolasi C 7 + C 3 C 4 + C 6 transchetolasi C 5 + C 4 C 3 + C 6
106 Parte non ossidativa della via del pentoso fosfato transchetolasi C 5 + C 5 C 3 + C 7 transaldolasi C 7 + C 3 C 4 + C 6 transchetolasi C 5 + C 4 C 3 + C 6 Il risultato complessivo di queste reazioni è la formazione di due esosi e di un triosio a partire da tre pentosi : 3 C 5 2 C 6 + C 3
107 C 5 + C 5 C 3 + C 7 transchetolasi Il donatore dell unit unità bicarboniosa è lo xilulosio 5-fosfato, un epimero del ribulosio 5-fosfato.. Un chetosio è un substrato della transchetolasi soltanto se il gruppo ossidrilico su C-3 C 3 ha la configurazione dello xilulosio anziché del ribulosio. C 3 Il ribulosio 5-fosfato viene convertito nell epimero epimero appropriato per la reazione della transchetolasi dalla fosfopentosio epimerasi.
108 C 5 + C 5 C 3 + C 7 transchetolasi
109 C 7 + C 3 C 4 + C 6 transaldolasi Intermedio glicolitico
110 C 5 + C 4 C 3 + C 6 transchetolasi Intermedi glicolitici
111 La via del pentoso fosfato
112 Via del pentosio fosfato La somma delle reazioni della via del pentosio fosfato è: 2 Xilulosio 5-fosfato + ribosio 5-fosfato 5 2 fruttosio 6-fosfato 6 + gliceraldeide 3-fosfato Lo xilulosio 5-fosfato può essere formato a partire da ribosio 5-fosfato 5 ad opera dell azione sequenziale della fosfopentosio isomerasi e della fosfopentosio epimerasi pertanto: 3 Ribosio 5-fosfato 5 2 fruttosio 6-fosfato 6 + gliceraldeide 3-fosfato L eccesso eccesso di ribosio 5-fosfato 5 formato dalla via del pentosio fosfato può essere completamente convertito in intermedi glicolitici. Inoltre, grazie alla via del pentosio fosfato: - Ogni molecola di ribosio ingerita nella dieta può essere convertita in intermedi glicolitici. - Gli scheletri carboniosi degli zuccheri possono essere riarrangiati per soddisfare esigenze fisiologiche
113 Regolazione della via del pentoso fosfato Il glucosio 6-fosfato 6 viene metabolizzato sia dalla via glicolitica sia dalla via del pentosio fosfato. La regolazione della via del pentoso fosfato avviene al livello della prima reazione, la deidrogenazione del glucosio 6-fosfato, 6 che è una reazione irreversibile. Il fattore di regolazione più importante è il livello di NADP +, l accettorel di elettroni in questa reazione catalizzata dalla glucosio 6-fosfato 6 deidrogenasi. La parte non ossidativa della via del pentoso principalmente dalla disponibilità dei substrati. fosfato è controllata
114 Metabolismo del glucosio 6-fosfato 6 in 4 differenti condizioni metaboliche: Quattro modi della via del pentoso fosfato
115 Modo 1: E E necessario molto più ribosio 5-fosfato 5 che NADPH Es.: cellule in rapida divisione necessitano di ribosio 5-fosfato 5 per la sintesi di DNA
116 Modo 2: Il fabbisogno di NADPH e quello di ribosio 5-fosfato 5 sono bilanciati
117 Modo 3: E E necessario più NADPH che ribosio 5-fosfato5 Es.: il tessuto epatico richiede una elevata concentrazione di NADPH per la sintesi di acidi grassi. La via gluconeogenetica risintetizza glucosio 6-fosfato
118 Modo 4: Sono necessari sia NADPH che ATP
119
120
121 Il glucosio può essere trasformato in acido glucuronico
122 L acido glucuronico è importante nella detossificazione e nell escrezione escrezione di composti organici estranei L UDP-glucuronato è il donatore di unità glucuronosidiche utilizzato da una famiglia di enzimi detossificanti che agiscono su composti non polari come molte tossine ambientali e carcinogeni La coniugazione di questi composti con il glucuronato (glucuronazione) produce derivati più polari rimossi più facilmente dal sangue da parte dei reni.
123
124 Un modo alternativo per generare energia è una variazione della respirazione in cui l accettorel finale di elettroni è diverso dall ossigeno (respirazione anaerobica)
125 Un secondo sistema di generazione di energia prevede il ricorso a sostanze chimiche inorganiche invece che organiche. Gli organismi in grado di utilizzare sostanze chimiche inorganiche come donatori di elettroni sono un tipo di chemiotrofi detti chemiolitotrofi. Esempi di donatori di elettroni inorganici sono : l idrogeno l solforato (H 2 S), l idrogeno l gassoso (H 2 ), il ferro ferroso (Fe 2+ ) e l ammoniaca l (NH 3 ) I chemiolitotrofi hanno dei sistemi di trasporto degli elettroni simili a quelli dei chemioorganotrofi e determinano una forza protomotrice che porta alla sintesi di ATP.
126 L ossidazione dell idrogeno. Diversi chemiolitotrofi sono in grado di utilizzare come donatore di elettroni l idrogeno, l H 2, un prodotto moilto comune del metabolismo microbico. Bioenergetica negli idrogenobatteri aerobi
127 La riduzione del solfato. Oltre all idrogeno (H 2 ) l idrogenasi l può anche sfruttare H 2 che si genera dal catabolismo dei composti organici come il lattato e il piruvato.
128 Un gran numero di microorganismi,, così come le piante verdi, sono fototrofi,, usano cioè la luce come fonte di energia nel processo di fotosintesi.
METABOLISMO DEL Glucosio
 METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
METABLISM DEL Glucosio Il metabolismo del glucosio può essere suddiviso nelle seguenti vie metaboliche: Glicolisi ssidazione del glucosio in acido piruvico e acido lattico. Via del pentoso fosfato Via
GLUCONEOGENESI. Sintesi (GENESI) di nuove (NEO) molecole di glucosio
 di nuove (NEO) molecole di glucosio") GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
GLUCONEOGENESI Sintesi (GENESI) di nuove (NEO) molecole di glucosio CATABOLISMO ANABOLISMO OSSIDAZIONI Produzione di ATP RIDUZIONI Consumo di ATP La GLUCONEOGENESI è un PROCESSO ANABOLICO La gluconeogenesi
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
GLICOLISI e CATABOLISMO degli ESOSI
 GLICOLISI e CATABOLISMO degli ESOSI 1 ph= 7.0 2 Digestione dei polisaccaridi Amilasi salivare e pancreatica = endoglucosidasi 3 Digestione degli oligosaccaridi maltosio, maltotriosio, destrine, Saccarosio,
GLICOLISI e CATABOLISMO degli ESOSI 1 ph= 7.0 2 Digestione dei polisaccaridi Amilasi salivare e pancreatica = endoglucosidasi 3 Digestione degli oligosaccaridi maltosio, maltotriosio, destrine, Saccarosio,
Il ciclo dell acido citrico
 Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE. L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO
 GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
GLUCONEOGENESI SINTESI DI NUOVO GLUCOSIO A PARTIRE DA FONTI NON GLUCIDICHE L UOMO CONSUMA QUASI 160 g DI GLUCOSIO AL GIORNO 75% DI TALE GLUCOSIO E NEL CERVELLO I FLUIDI CORPOREI CONTENGONO SOLO 20 g DI
Regolazione del metabolismo del glucosio
 Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
SISTEMI ENERGETICI. L ATP privato di uno dei suoi 3 radicali fosforici diventa ADP (adenosindifosfato).
.") SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA
 CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
GLICOGENOLISI & GLICOGENOSINTESI
 GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
GLICOGENOLISI & GLICOGENOSINTESI ALIMENTARE Glucosio GLUCONEOGENESI GLICOGENOLISI (epatica) Glicemia: 70-90 mg/100ml ~ 5 mm GLICOGENO Nel fegato fino al 6-10% della massa epatica (~100 grammi) Nel muscolo
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
Bioenergetica & metabolismo (1)
") Bioenergetica & metabolismo (1) Tutte le vie metaboliche sono interconnesse (come le linee di una grande metropolitana) La maggior parte degli organismi viventi hanno vie metaboliche molto simili 1 Vie
Bioenergetica & metabolismo (1) Tutte le vie metaboliche sono interconnesse (come le linee di una grande metropolitana) La maggior parte degli organismi viventi hanno vie metaboliche molto simili 1 Vie
glicogeno Glucosio 6-P Piruvato Acetil CoA Intermedi del ciclo di Krebs
 PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
PIRUVAT glicogeno Glicogeno sintesi glicogenolisi Glucosio Glucosio 6-P Ribosio 5-P glicolisi Gluconeogenesi Amino acidi Piruvato Acetil CoA lattato Intermedi del ciclo di Krebs Concentrazione di glicogeno
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
METABOLISMO DEL GLICOGENO Struttura del glicogeno DEMOLIZIONE DEL GLICOGENO: GLICOGENOLISI 1) distacco progressivo di unità glucosidiche con formazione di glucosio-1- fosfato: enzima: glicogeno fosforilasi
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2! 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Metabolismo degli amminoacidi
 Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
Metabolismo degli amminoacidi Gli amminoacidi derivati in gran parte dalla degradazione delle proteine della dieta o intracellulari possono essere ossidati per generare e energia. La quantità di energia
BIOLOGIA GENERALE 22-24 ottobre 2007
 Biologia generale Massolo Alessandro [email protected]; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
Biologia generale Massolo Alessandro [email protected]; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA. Angela Chambery Lezione 24
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 24 Una panoramica del metabolismo Concetti chiave: Organismi diversi utilizzano strategie differenti per catturare l'energia
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 24 Una panoramica del metabolismo Concetti chiave: Organismi diversi utilizzano strategie differenti per catturare l'energia
Da dove prendono energia le cellule animali?
 Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
Da dove prendono energia le cellule animali? La cellula trae energia dai legami chimici contenuti nelle molecole nutritive Probabilmente le più importanti sono gli zuccheri, che le piante sintetizzano
metano alcool metilico formaldeide acido formico CO 2
 -4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
-4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Mais, riso, patate, granozucchero di canna o barbabietola Latte da zucchero Polisaccaride Amido - - Disaccaride Maltosio Saccarosio Lattosio
 Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
Abbiamo visto che i carboidrati sono la principale fonte di energia nell'alimentazione umana. Nella nostra dieta, sono sopratutto presenti nella forma di amido, ma anche il saccarosio è presente in una
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara
 Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
Giovanni Di Bonaventura, Ph.D., B.Sc. Università di Chieti-Pescara Richieste metaboliche: elementi essenziali In generale, i batteri per la loro crescita hanno bisogno di elementi essenziali. In particolare
dieta vengono convertiti in composti dei corpi chetonici.
 Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
ogni anno 200 miliardi di tonnellate di CO 2 vengono convertite in biomassa
 ogni anno 200 miliardi di tonnellate di CO 2 vengono convertite in biomassa La maggior parte di questo carbonio viene incorporato in composti organici attraverso le reazioni di riduzione del carbonio associate
ogni anno 200 miliardi di tonnellate di CO 2 vengono convertite in biomassa La maggior parte di questo carbonio viene incorporato in composti organici attraverso le reazioni di riduzione del carbonio associate
Prof. Maria Nicola GADALETA
 Prof. Maria Nicola GADALETA E-mail: [email protected] Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: [email protected] Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Metabolismo degli aminoacidi
 Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Valitutti, Taddei, Kreuzer, Massey, Sadava, Hills, Heller, Berenbaum
 Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
Dal carbonio agli OGM VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono evidenziate
Il ciclo nutrizione / digiuno
 Il ciclo nutrizione / digiuno Aspetti generali La fase di assorbimento è il periodo che va dalle 2 alle 4 ore successive all assunzione di un pasto. Durante questo intervallo si verifica un transitorio
Il ciclo nutrizione / digiuno Aspetti generali La fase di assorbimento è il periodo che va dalle 2 alle 4 ore successive all assunzione di un pasto. Durante questo intervallo si verifica un transitorio
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO 1 2 DESTINI DEL GLUCOSIO 6-FOSFATO 3 RAPPRESENTAZIONE SCHEMATICA DEL METABOLISMO DEL GLUCOSIO 4 Glucosio-6-fosfatasi Omeostasi del glicogeno Nell'organismo animale, il glicogeno
METABOLISMO DEL GLICOGENO 1 2 DESTINI DEL GLUCOSIO 6-FOSFATO 3 RAPPRESENTAZIONE SCHEMATICA DEL METABOLISMO DEL GLUCOSIO 4 Glucosio-6-fosfatasi Omeostasi del glicogeno Nell'organismo animale, il glicogeno
Niccolò Taddei Biochimica
 Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
Niccolò Taddei Biochimica VERSO L UNIVERSITÀ Le domande sono tratte dalle prove di ammissione emesse annualmente dal Ministero dell Istruzione, dell Università e della Ricerca (MIUR) e le soluzioni sono
Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone = C 6 O 6 (H 2 O) 6
 6") Carboidrati Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone Glucosio Carboidrati modificati: membrane cellulari, acidi nucleici, Da carbonio idrato: glucosio= C 6 H 12
Carboidrati Carboidrati puri : zucchero e amido nei cibi, cellulosa nel legno, carta e cotone Glucosio Carboidrati modificati: membrane cellulari, acidi nucleici, Da carbonio idrato: glucosio= C 6 H 12
A livello della loro struttura chimica, come i grassi, anche i carboidrati sono composti ternari, formati cioè da tre molecole:
 I carboidrati costituiscono la fonte di energia principale per lo svolgimento di tutte le funzioni organiche (mantenimento della temperatura corporea, battito cardiaco, funzioni cerebrali, digestione,
I carboidrati costituiscono la fonte di energia principale per lo svolgimento di tutte le funzioni organiche (mantenimento della temperatura corporea, battito cardiaco, funzioni cerebrali, digestione,
Metabolismo degli amminoacidi. Ciclo dell urea
 Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
Metabolismo degli amminoacidi Ciclo dell urea Biosintesi di amminoacidiporfirine, creatina, carnitina, ormoni, nucleotidi Gli amminoacidi possono subire una degradazione ossidativa in 3 diverse situazioni
FUNZIONI DEI MITOCONDRI
 FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
FUNZIONI DEI MITOCONDRI La funzione principale dei mitocondri è di compiere le trasformazioni energetiche indispensabili per le funzioni cellulari. Metabolismo energetico: insieme delle reazioni chimiche
GLUCOSIO GLUCOSIO-6-FOSFATO ISOMERASI FRUTTOSIO-6-FOSFATO ATP FRUTTOSIO-1,6-DIFOSFATO ALDOLASI ISOMERASI
 GLUCOSIO ESOCHINASI (costitutivo) inibizione feed-back GLUCOCHINASI (induttivo) GLUCOSIO-6-FOSFATO ISOMERASI - 2 FRUTTOSIO-6-FOSFATO FOSFOFRUTTOCINASI allosterico reg +: e Fr-2,6-dP reg -:, citrato FRUTTOSIO-1,6-DIFOSFATO
GLUCOSIO ESOCHINASI (costitutivo) inibizione feed-back GLUCOCHINASI (induttivo) GLUCOSIO-6-FOSFATO ISOMERASI - 2 FRUTTOSIO-6-FOSFATO FOSFOFRUTTOCINASI allosterico reg +: e Fr-2,6-dP reg -:, citrato FRUTTOSIO-1,6-DIFOSFATO
unità C2. Le trasformazioni energetiche nelle cellule
 unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
Il glicogeno (riserva di glucosio) è immagazzinato nel fegato e nei muscoli (con finalità diverse )
 è immagazzinato nel fegato e nei muscoli (con finalità diverse )") Glicogeno Glicogeno...1 Glicogenolisi...3 Glicogenosintesi...7 Regolazione ormonale della glicogenolisi e delle glicogenosintesi...13 Il recettore del glucagone e quello dell adrenalina sono 2 GPCR (G-Protein
Glicogeno Glicogeno...1 Glicogenolisi...3 Glicogenosintesi...7 Regolazione ormonale della glicogenolisi e delle glicogenosintesi...13 Il recettore del glucagone e quello dell adrenalina sono 2 GPCR (G-Protein
Nota dell editore Presentazione
 00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
00PrPag 3-08-2007 11:42 Pagina V Autori Nota dell editore Presentazione XI XIII XV Parte I Chimica 1 Struttura dell atomo 3 Teorie atomiche 3 Costituenti dell atomo 4 Numeri quantici 5 Tipi di orbitali
Le Biomolecole I parte. Lezioni d'autore di Giorgio Benedetti
 Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Biosintesi non ribosomiale di metaboliti peptidici bioattivi
 Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Biologia Molecolare della Cellula 08 7: Regolazione ormonale del metabolismo del glicogeno e della glicolisi Desensitizzazione
 Biologia Molecolare della Cellula 08 7: Regolazione ormonale del metabolismo del glicogeno e della glicolisi Desensitizzazione camp Proteina-CHINASI camp DIPENDENTE (PKA) Proteine target : ε metabolici
Biologia Molecolare della Cellula 08 7: Regolazione ormonale del metabolismo del glicogeno e della glicolisi Desensitizzazione camp Proteina-CHINASI camp DIPENDENTE (PKA) Proteine target : ε metabolici
CELLULE EUCARIOTICHE
 CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
I Composti Organici. Le Biomolecole
 I Composti Organici I composti organici sono molecole tutte contenenti carbonio. Essi comprendono. 1. composti di interesse energetico che sono gli Idrocarburi ( i derivati del petrolio), 2. composti a
I Composti Organici I composti organici sono molecole tutte contenenti carbonio. Essi comprendono. 1. composti di interesse energetico che sono gli Idrocarburi ( i derivati del petrolio), 2. composti a
I carboidrati La maggior parte dei carboidrati, detti anche saccaridi o glucidi, è riconducibile alla formula chimica generale C n (H 2 O) n.
 n.") MODULO 3 IL METABOLISMO GLUCIDICO RICHIAMI SULLA CHIMICA DEI CARBOIDRATI Introduzione I carboidrati sono macromolecole organiche molto abbondanti in natura. Essi svolgono funzioni metaboliche e strutturali
MODULO 3 IL METABOLISMO GLUCIDICO RICHIAMI SULLA CHIMICA DEI CARBOIDRATI Introduzione I carboidrati sono macromolecole organiche molto abbondanti in natura. Essi svolgono funzioni metaboliche e strutturali
Carboidrati! Oligosaccaridi Polisaccaridi. B) Carboidrati complessi
 Carboidrati complessi") Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
Carboidrati! Sono i composti organici più abbondanti sulla terra Contengono: C, H, O Sono Aldeidi o Chetoni poliossidrilici Possono essere suddivisi in: A) Zuccheri semplici Monosaccaridi B) Carboidrati
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
http://sds.coniliguria.it 1
 Aspetti metabolici dell esercizio fisico Genova 22 gennaio 2011 A cura di Attilio TRAVERSO http://sds.coniliguria.it 1 L allenamento produce modificazioni fisiologiche (adattamenti) in quasi tutti i sistemi
Aspetti metabolici dell esercizio fisico Genova 22 gennaio 2011 A cura di Attilio TRAVERSO http://sds.coniliguria.it 1 L allenamento produce modificazioni fisiologiche (adattamenti) in quasi tutti i sistemi
La fotosintesi: energia dal Sole
 La fotosintesi: energia dal Sole Gli organismi fotosintetici usano la luce del Sole, l acqua del suolo e il CO 2 dell atmosfera per produrre composti organici e liberare O 2 grazie alla fotosintesi. Sadava
La fotosintesi: energia dal Sole Gli organismi fotosintetici usano la luce del Sole, l acqua del suolo e il CO 2 dell atmosfera per produrre composti organici e liberare O 2 grazie alla fotosintesi. Sadava
TIAMINA (vitamina B1, aneurina)
") TIAMINA (vitamina B1, aneurina) Fonti alimentari: cereali integrali (germe e crusca), carne (maiale), lievito di birra, legumi Abbondanza nell organismo umano : 30-100 mg RDA (assunzione giornaliera di
TIAMINA (vitamina B1, aneurina) Fonti alimentari: cereali integrali (germe e crusca), carne (maiale), lievito di birra, legumi Abbondanza nell organismo umano : 30-100 mg RDA (assunzione giornaliera di
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici
 I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
I composti organici della vita: carboidrati, lipidi, proteine e acidi nucleici La seta della tela di ragno è un insieme di macromolecole, dette proteine. Sono le caratteristiche fisico-chimiche di queste
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato
 2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
2 a reazione della glicolisi: isomerizzazione del glucosio 6-fosfato in fruttosio 6-fosfato Glucosio 6-fosfato isomerasi Glucosio 6-fosfato anomero α Fruttosio 6-fosfato anomero α Glucosio 6-fosfato Fruttosio
LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI PROTEINE. sono ACIDI NUCLEICI. molecole complesse = POLIMERI. formate dall'unione di
 LE BIOMOLECOLE LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI LE BIOMOLECOLE sono LIPIDI PROTEINE ACIDI NUCLEICI molecole complesse = POLIMERI formate dall'unione di molecole semplici = MONOMERI
LE BIOMOLECOLE LE BIOMOLECOLE DETTE ANCHE MOLECOLE ORGANICHE; CARBOIDRATI LE BIOMOLECOLE sono LIPIDI PROTEINE ACIDI NUCLEICI molecole complesse = POLIMERI formate dall'unione di molecole semplici = MONOMERI
Degradazione delle proteine della dieta. Catabolismo degli aminoacidi
 Degradazione delle proteine della dieta Catabolismo degli aminoacidi LE PROTEINE DELLA DIETA SONO DEGRADATE ENZIMATICAMENTE AD AMINOACIDI LIBERI L ingresso di una proteina nello stomaco stimola la mucosa
Degradazione delle proteine della dieta Catabolismo degli aminoacidi LE PROTEINE DELLA DIETA SONO DEGRADATE ENZIMATICAMENTE AD AMINOACIDI LIBERI L ingresso di una proteina nello stomaco stimola la mucosa
METABOLISMO E SISTEMI ENERGETICI
 METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
METABOLISMO E SISTEMI ENERGETICI 1 Obiettivi della lezione 1) Capire come l organismo converta il cibo che ingeriamo in ATP per fornire ai muscoli l energia che essi necessitano per contrarsi. 2) Esaminare
METABOLISMO DEI CARBOIDRATI I: PROCESSI ANAEROBICI NELLA PRODUZIONE DELL'ENERGIA METABOLICA
 AITL 13 METABLISM DEI ARBIDRATI I: RESSI ANAERBII NELLA RDUZINE DELL'ENERGIA METABLIA Il nostro studio dettagliato del metabolismo inizierà considerando le fasi anaerobiche del metabolismo dei carboidrati
AITL 13 METABLISM DEI ARBIDRATI I: RESSI ANAERBII NELLA RDUZINE DELL'ENERGIA METABLIA Il nostro studio dettagliato del metabolismo inizierà considerando le fasi anaerobiche del metabolismo dei carboidrati
PROTEINE. sono COMPOSTI ORGANICI QUATERNARI
 PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
DNA - RNA. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi.
 DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA
 PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA Svolgono la loro funzione legando reversibilmente l OSSIGENO. Aumentano la solubilità dell ossigeno nel plasma, da 3ml/L a 220 ml/l. La mioglobina
PROTEINE RESPIRATORIE DEI VERTEBRATI EMOGLOBINA E MIOGLOBINA Svolgono la loro funzione legando reversibilmente l OSSIGENO. Aumentano la solubilità dell ossigeno nel plasma, da 3ml/L a 220 ml/l. La mioglobina
N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi
 Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Glicina (Gly) Alanina (Ala) N 2, malgrado la sua abbondanza, è un fattore limitante la crescita della maggior parte degli organismi La digestione delle proteine endopeptidasi H O R H O R R H 3+ N -C-C-NH-C-C-NH-C-C-NH-C-C-NH-C-COO
Transferasi che hanno come coenzima FOSFOPANTOTEINA E COENZIMA A (derivati dalla VITAMINA B3 )
") Transferasi che hanno come coenzima FSFPANTTEINA E ENZIMA A (derivati dalla VITAMINA B3 ) Altre transferasi hanno come oe: Vitamina B3 (sintetizzata in vegetali e molti procarioti anche ospiti dell intestino
Transferasi che hanno come coenzima FSFPANTTEINA E ENZIMA A (derivati dalla VITAMINA B3 ) Altre transferasi hanno come oe: Vitamina B3 (sintetizzata in vegetali e molti procarioti anche ospiti dell intestino
Esempio di domande possibili alla prova orale di Biochimica Generale ed Applicata AA 2009-10 Farmacia - FERIOTTO
 Esempio di domande possibili alla prova orale di Biochimica Generale ed Applicata AA 2009-10 Farmacia - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali
Esempio di domande possibili alla prova orale di Biochimica Generale ed Applicata AA 2009-10 Farmacia - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali
Biosintesi dei triacilgliceroli
 Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
Biosintesi dei triacilgliceroli Destino della maggior parte degli acidi grassi sintetizzati o ingeriti: Ø triacilgliceroli (riserva) Ø fosfolipidi (membrane) in base alle necessità metaboliche I triacilgliceroli
Domande relative alla specializzazione in: Biochimica clinica
 Domande relative alla specializzazione in: Biochimica clinica Domanda #1 (codice domanda: n.403) : Nell'ibridazione fluorescente in situ (Fluorescent in situ hybridization "FISH"), che tipo di sonde fluorescenti
Domande relative alla specializzazione in: Biochimica clinica Domanda #1 (codice domanda: n.403) : Nell'ibridazione fluorescente in situ (Fluorescent in situ hybridization "FISH"), che tipo di sonde fluorescenti
Le idee della chimica
 G. Valitutti A.Tifi A.Gentile Seconda edizione Copyright 2009 Zanichelli editore Capitolo 25 Le basi della biochimica 1. I carboidrati 2. I lipidi 3. Gli amminoacidi, i peptidi e le proteine 4. La struttura
G. Valitutti A.Tifi A.Gentile Seconda edizione Copyright 2009 Zanichelli editore Capitolo 25 Le basi della biochimica 1. I carboidrati 2. I lipidi 3. Gli amminoacidi, i peptidi e le proteine 4. La struttura
Principali vie di utilizzo del Glucosio
 Principali vie di utilizzo del Glucosio GLICOGENO (Polimero di riserva) Polimero lineare e ramificato di glucosio che si accumula nel fegato e nel muscolo. Nella catena lineare le molecole di glucosio
Principali vie di utilizzo del Glucosio GLICOGENO (Polimero di riserva) Polimero lineare e ramificato di glucosio che si accumula nel fegato e nel muscolo. Nella catena lineare le molecole di glucosio
La nutrizione e il metabolismo
 La nutrizione e il metabolismo 1. Le sostanze nutritive, o nutrienti 2. Definizione di metabolismo 3. Il metabolismo dei carboidrati 4. Il metabolismo dei lipidi 1 L apparato respiratorio 5. Il metabolismo
La nutrizione e il metabolismo 1. Le sostanze nutritive, o nutrienti 2. Definizione di metabolismo 3. Il metabolismo dei carboidrati 4. Il metabolismo dei lipidi 1 L apparato respiratorio 5. Il metabolismo
GENERALITA SUL METABOLISMO. DIGESTIONE e ASSORBIMENTO dei CARBOIDRATI GLICOLISI
 GENERALITA SUL METABOLISMO DIGESTIONE e ASSORBIMENTO dei CARBOIDRATI GLICOLISI ENERGIA per Ø gradiente elettrochimico (Na + /K + ATPasi) Ø processi biosintetici (es. sintesi proteica) Ø trasporto transmembrana
GENERALITA SUL METABOLISMO DIGESTIONE e ASSORBIMENTO dei CARBOIDRATI GLICOLISI ENERGIA per Ø gradiente elettrochimico (Na + /K + ATPasi) Ø processi biosintetici (es. sintesi proteica) Ø trasporto transmembrana
Fosforilazione ossidativa
 Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
di glucosio da parte di tutte le cellule, facilitandone il trasporto transmembranario 2. aumenta l utilizzazione
 non c Ormone ipoglicemizzante, causa un forte abbassamento della glicemia (70-100 mg/100 ml) perché esalta i processi responsabili della sottrazione di glucosio dal sangue e inibisce i processi responsabili
non c Ormone ipoglicemizzante, causa un forte abbassamento della glicemia (70-100 mg/100 ml) perché esalta i processi responsabili della sottrazione di glucosio dal sangue e inibisce i processi responsabili
Attenzione : lunedì 29 aprile NON ci sarà lezione
 Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
Attenzione : lunedì 29 aprile NON ci sarà lezione Metabolismo dei lipidi a) Ossidazione degli acidi grassi Triacilgliceroli (90% del totale) Gruppi metilenici o metilici Richiedono molto O 2 per essere
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Le proteine. Le proteine sono i mattoncini che costituiscono gli organismi viventi.
 Le proteine Le proteine sono i mattoncini che costituiscono gli organismi viventi. Per avere un idea più precisa, basti pensare che tutti i muscoli del nostro corpo sono composti da filamenti di proteine
Le proteine Le proteine sono i mattoncini che costituiscono gli organismi viventi. Per avere un idea più precisa, basti pensare che tutti i muscoli del nostro corpo sono composti da filamenti di proteine
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI
 LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
LIPIDI e METABOLISMO DEGLI ACIDI GRASSI Lipidi I lipidi sono i principali costituenti delle membrane biologiche. Sono biomolecole insolubile in acqua ma solubili in solventi organici. La loro idrofobicità
L adattamento dei batteri. Strategie di adattamento
 L adattamento dei batteri Strategie di adattamento mutazione trasferimento genico orizzontale regolazione dell espressione genica regolazione della trascrizione regolazione della traduzione regolazione
L adattamento dei batteri Strategie di adattamento mutazione trasferimento genico orizzontale regolazione dell espressione genica regolazione della trascrizione regolazione della traduzione regolazione
scaricato da www.sunhope.it
 Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Βeta ossidazione degli acidi grassi BETA OSSIDAZIONE DEGLI ACIDI GRASSI 13/12/2013 SEMINARIO. Acidi grassi (2)
") Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
SECREZIONE DI INSULINA:
 SECREZIONE DI INSULINA: O Ca ++ VGCC K-ATP GLUT2 Ca ++ K + O Ca ++ HK I P GK ATP O ADP piruvato acidi grassi corpi chetonici aminoacidi secretina glucagone incretine: colecistochinina (CCK) peptide inibitore
SECREZIONE DI INSULINA: O Ca ++ VGCC K-ATP GLUT2 Ca ++ K + O Ca ++ HK I P GK ATP O ADP piruvato acidi grassi corpi chetonici aminoacidi secretina glucagone incretine: colecistochinina (CCK) peptide inibitore
Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO
 Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali delle biomolecole. -
Esempio di domande possibili alla prova orale di Biochimica AA 2011-12 Farmacia1 - FERIOTTO Lo studente spieghi: - Geometria dei legami del carbonio. - Principali gruppi funzionali delle biomolecole. -
che è caratteristico degli alcoli (es CH 3 -CH 2 -OH etanolo) gruppo carbonilico che si trova in composti detti chetoni;
 gruppo carbonilico che si trova in composti detti chetoni;") LE BIOMOLECOLE 1. IL CARBONIO E I SUOI COMPOSTI Il carbonio è l'elemento base del mondo vivente. Ciò si deve alla sua particolare capacità di formare 4 legami covalenti, in genere molto forti, o con altri
LE BIOMOLECOLE 1. IL CARBONIO E I SUOI COMPOSTI Il carbonio è l'elemento base del mondo vivente. Ciò si deve alla sua particolare capacità di formare 4 legami covalenti, in genere molto forti, o con altri
METABOLISMO DEL GLICOGENO MUSCOLO, FEGATO
 METABOLISMO DEL GLICOGENO MUSCOLO, FEGATO DEGRADAZIONE DEL GLICOGENO Glicogeno fosforilasi Scinde i legami glicosidici α(1 4) uno alla volta, attraverso una FOSFOROLISI a partire dalle estremità non riducenti
METABOLISMO DEL GLICOGENO MUSCOLO, FEGATO DEGRADAZIONE DEL GLICOGENO Glicogeno fosforilasi Scinde i legami glicosidici α(1 4) uno alla volta, attraverso una FOSFOROLISI a partire dalle estremità non riducenti
PROGRAMMA DI BIOLOGIA. CLASSE 2^ F a. s. 2014 2015. Prof.ssa RUBINO ALESSANDRA
 ISTITUTO TECNICO INDUSTRIALE DI STATO "ENRICO FERMI" Via Luosi n. 23-41124 Modena Tel. 059211092 059236398 - (Fax): 059226478 E-mail: [email protected] Pagina web: www.fermi.mo.it PROGRAMMA DI BIOLOGIA
ISTITUTO TECNICO INDUSTRIALE DI STATO "ENRICO FERMI" Via Luosi n. 23-41124 Modena Tel. 059211092 059236398 - (Fax): 059226478 E-mail: [email protected] Pagina web: www.fermi.mo.it PROGRAMMA DI BIOLOGIA
ENZIMI CINETICA ENZIMATICA
 ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
ENZIMI PERCHE UNA REAZIONE AVVENGA, SI DEVONO SODDISFARE TRE CONDIZIONI I SUBSTRATI DEVONO ENTRARE IN COLLISIONE LA COLLISIONE DEVE AVVENIRE CON ORIENTAMENTO CORRETTO I REAGENTI DEVONO AVERE ENERGIA SUFFICIENTE
Macromolecole Biologiche. I domini (III)
") I domini (III) Domini α/β La cross over connection è l unità costitutiva su cui si basa la topologia di 3 tipi di domini α/β osservati nelle proteine: - α/β barrel - motivi ricchi di Leu (fold a ferro
I domini (III) Domini α/β La cross over connection è l unità costitutiva su cui si basa la topologia di 3 tipi di domini α/β osservati nelle proteine: - α/β barrel - motivi ricchi di Leu (fold a ferro
NUCLEOTIDI e ACIDI NUCLEICI
 NUCLEOTIDI e ACIDI NUCLEICI Struttura dei nucleotidi Il gruppo fosfato conferisce carica negativa e proprietà acide FUNZIONI DEI NUCLEOTIDI MOLECOLE DI RISERVA DI ENERGIA L idrolisi dei nucleosidi trifosfato
NUCLEOTIDI e ACIDI NUCLEICI Struttura dei nucleotidi Il gruppo fosfato conferisce carica negativa e proprietà acide FUNZIONI DEI NUCLEOTIDI MOLECOLE DI RISERVA DI ENERGIA L idrolisi dei nucleosidi trifosfato
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 29 La via del pentosio fosfato Concetti chiave: La via del pentosio fosfato è costituita da tre fasi in cui viene prodotto
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 29 La via del pentosio fosfato Concetti chiave: La via del pentosio fosfato è costituita da tre fasi in cui viene prodotto
Bioenergetica e fisiologia dell esercizio 8. Risposte all allenamento: adattamenti biochimici e effetti sulla resistenza
 Bioenergetica e fisiologia dell esercizio 8. Risposte all allenamento: adattamenti biochimici e effetti sulla resistenza Prof. Carlo Capelli, Fisiologia Generale e dell Esercizio, Facoltà di Scienze Motorie,
Bioenergetica e fisiologia dell esercizio 8. Risposte all allenamento: adattamenti biochimici e effetti sulla resistenza Prof. Carlo Capelli, Fisiologia Generale e dell Esercizio, Facoltà di Scienze Motorie,
 forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
GLICOLISI ADP ATP. Condizioni aerobiche. Condizioni anaerobiche ATP
 GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
GLICOLISI È la parte iniziale del processo di degradazione ossidativa dei carboidrati. Durante la loro ossidazione l energia immagazzinata nel loro scheletro carbonioso è utilizzata per produrre ATP e
I RECETTORI ACCOPPIATI A PROTEINE G
 I RECETTORI ACCOPPIATI A PROTEINE G I recettori accoppiati a proteine G sono costituiti da un unica subunità formata da un filamento di aminoacidi che attraversa 7 volte la membrana plasmatica Il sito
I RECETTORI ACCOPPIATI A PROTEINE G I recettori accoppiati a proteine G sono costituiti da un unica subunità formata da un filamento di aminoacidi che attraversa 7 volte la membrana plasmatica Il sito
Cap.16 GLICOLISI. Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP H 2 O 2 NAD + 2 NADH + 2H +
 Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
OBESITA. 11 Aprile 2013
 OBESITA 11 Aprile 2013 L obesità è una patologia cronica caratterizzata dall eccessivo accumulo di tessuto adiposo nell organismo ed è causata da fattori ereditari e non ereditari che determinano un introito
OBESITA 11 Aprile 2013 L obesità è una patologia cronica caratterizzata dall eccessivo accumulo di tessuto adiposo nell organismo ed è causata da fattori ereditari e non ereditari che determinano un introito