PIRUVATO DEIDROGENASI
|
|
|
- Leona Martini
- 6 anni fa
- Visualizzazioni
Transcript
1
2 PIRUVATO DEIDROGENASI IL PIRUVATO CHE DERIVA DALLA OSSIDAZIONE DEI CARBOIDRATI E UNA DELLE FONTI DI ACETIL-CoA, MOLECOLA CHE ENTRA NEL CICLO TCA PER ESSERE COMPLETAMENTE OSSIDATA LA CONVERSIONE DEL PIRUVATO IN ACETIL-CoA E UNA DECARBOSSILAZIONE OSSIDATIVA OPERATA DA UN COMPLESSO NOTO COME PIRUVATO DEIDROGENASI
3 LA REAZIONE E MOLTO COMPLESSA E PREVEDE NON SOLO LA DECARBOSSILAZIONE DEL PIRUVATO E L ATTIVAZIONE METABOLICA DEI DUE ATOMI DI CARBONIO RESTANTI, MA ANCHE LA PRODUZIONE DI EQUIVALENTI RIDUCENTI SOTTO FORMA DI NADH LA REAZIONE E FORTEMENTE ESOERGONICA E PRATICAMENTE IRREVERSIBILE. IL COMPLESSO DELLA DA TRE ENZIMI: - PIRUVATO DECARBOSSILASI (E1) - DIIDROLIPOAMMIDE TRANSACETILASI (E2) - LIPOAMMIDE DEIDROGENASI (E3) PIRUVATO DEIDROGENASI E COSTITUITO
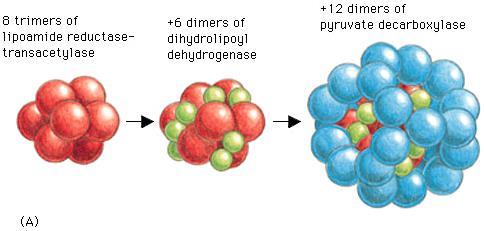
4

5 PER LA SUA ATTIVITA CATALITICA QUESTO COMPLESSO ENZIMATICO NECESSITA DI 5 COENZIMI: TIAMINA PIROFOSFATO (TPP) (Vitamina B1 che diviene TPP dopo una pirofosforilazione ATP-dipendente). Partecipa a tutte le decarbossilazioni di -chetoacidi. La parte attiva del coenzima è rappresentata dall anello tiazolo in cui il carbonio a carattere acido (tra l N e lo S) forma un carbanione che può legare il carbonio carbonilico producendo un composto di addizione. Questo composto va incontro ad una decarbossilazione non ossidativa in cui l anello tiazolico funge da trappola di elettroni dando luogo alla formazione di un eneamina stabilizzata per risonanza. Si produce l IDROSSIETIL- TPP LIPOAMMIDE E tipicamente un trasportatore di elettroni e di gruppi acile che lega allo zolfo della lipoammide all IDROSSIETIL-TPP; contemporaneamente si ossida l aldeide e si riduce lo zolfo. Si genera così un gruppo acilico che verrà successivamente trasferito al Coenzima A
6 COENZIMI FLAVINICI Abbiamo il FLAVIN ADENIN DINUCLEOTIDE (FAD) (Vitamina B2; riboflavina) e il FLAVIN MONONUCLEOTIDE (FMN). Per entrambi i coenzimi la parte funzionale è l ANELLO ISOALLOSSAZINICO che funge da accettore di elettroni. Sono tipicamente coenzimi che partecipano a ossidazioni bielettroniche che possono procedere in due passaggi monoelettronici COENZIMA A Partecipa all attivazione dei gruppi acile tra cui l acetile che deriva dal piruvato. Deriva dalla vitamina ACIDO PANTOTENICO. La parte funzionale è il tiolo libero all estremità NAD Funge da accettore finale di elettroni

7

8 MECCANISMO DI AZIONE DELLA PIRUVATO DEIDROGENASI L OSSIDAZIONE DEL PIRUVATO AD ACETIL- COA COINVOLGE I COENZIMI APPENA ILLUSTRATI CHE AGISCONO DI CONCERTO CON I TRE ENZIMI
9 La lipoammide è legata all E2 ma interagisce con tutti e tre gli enzimi attraverso un braccio flessibile oscillante Infatti la sua catena copre una lunghezza di circa 1.4 nm, ma studi strutturali hanno evidenziato che la distanza tra la TPP legata ad E1 ed il FAD legato ad E3 è di circa nm Ne scaturisce che esistono almeno due bracci oscillanti che coprono questa distanza
10 REGOLAZIONE DELL ATTIVITA DELLA PIRUVATO DEIDROGENASI L attività di questo enzima è regolato mediante inibizione allosterica. - enzima E2 è inibita dall ACETIL-COA ed è attivata dal CoA-SH - E3 è inibita dal NADH ed è attivata dal NAD + - l ATP è un inibitore del complesso mentre l ADP è un attivatore - una delle regolazioni più complesse è quella a carico dell enzima E1 dell intero complesso. Infatti si ha una regolazione attraverso modificazione covalente dell enzima. La presenza di NADH e acetil-coa attivano una componente del complesso, la PIRUVATO DEIDROGENASI CHINASI, che fosforila tre specifici residui di serina di E1 con conseguente perdita di attività della piruvato deidrogenasi
11 Una PIRUVATO DEIDROGENASI FOSFATASI specifica rimuove idroliticamente il fosfato legato e riattiva in questo modo il complesso La fosfatasi è a sua volta attivata dalla concentrazione di ioni Ca e Mg Poiché ATP ed ADP differiscono per l affinità con l Mg, la concentrazione di Mg rispecchia il rapporto ATP/ADP nel mitocondrio. La piruvato deidrogenasi quindi rispecchia il livello di ATP presente venendo inattivata quando l ATP è abbondante

12

13 CICLO DEL TCA 1) L ACETIL-CoA ATTRAVERSO L ENZIMA CITRATO SINTASI VIENE CONDENSATO ALL OSSALACETATO E VIENE SINTETIZZATO ACIDO CITRICO Si tratta di una condensazione aldolica. Il gruppo metilico dell acetile attivato dal CoA perde un protone a cui segue l attacco nucleofilo del carbanione sul carbonio carbonilico dell ossalacetato. La reazione forma un prodotto instabile, il CITRIL-CoA che idrolizza spontaneamente nei prodotti ACIDO CITRICO e CoA La reazione è esoergonica ed è un sito regolatore dell intera via metabolica
14 2) IL CITRATO ATTRAVERSO L AZIONE DELL ACONITASI VIENE ISOMERIZZATO A cis-aconitato E QUINDI AD ISOCITRATO Questa reazione rende possibile la futura decarbossilazione del citrato Infatti i substrati della decarbossilazione sono generalmente - o -chetoacidi, per cui per poter essere ossidato a gruppo carbonilico, il gruppo ossidrilico terziario del citrato deve subire un trasferimento interno alla molecola
15 L enzima coinvolto è l ACONITASI, che produce l ISOCITRATO, un alcol secondario che può essere ossidato La reazione consiste di una disidratazione seguita da una idratazione con il cis-aconitato come intermedio

16 3) L ISOCITRATO ATTRAVERSO LA ISOCITRATO DEIDROGENASI VIENE DECARBOSSILATO ED OSSIDATO (CON CONTEMPORANEA RIDUZIONE DEL NAD) AD -CHETOGLUTARATO Rappresenta la prima delle due reazioni di decarbossilazione ossidativa Anche in questo caso si forma un intermedio, l OSSALSUCCINATO, instabile e legato all enzima che subisce la decarbossilazione prima del rilascio. Questo enzima è NADdipendente anche se esiste una forma NADP-dipendente nel citosol ed anche nel mitocondrio
17 4) L -CHETOGLUTARATO SUBISCE UNA DECARBOSSILAZIONE OSSIDATIVA (SI RIDUCE IL NAD) CON LA CONTEMPORANEA FORMAZIONE DI UN SUCCINIL-CoA. LA REAZIONE E CATALIZZATA DALL ENZIMA -CHETOGLUTARATO DEIDROGENASI Si tratta di una ulteriore decarbossilazione ossidativa di un - chetoacido con contemporanea formazione di un acil-coa L -chetoglutarato deidrogenasi è un complesso multienzimatico dotato di attività enzimatiche simili alla piruvato deidrogenasi e che necessita degli stessi coenzimi (TPP, NAD, FAD, acido lipoico e CoA-SH)
18 A questo punto del ciclo due atomi di carbonio sono stati introdotti come acetil-coa e due sono stati rimossi come CO 2 Nelle successive reazioni del ciclo, il succinil-coa che si è formato, viene convertito nel substrato che prende parte alla prima reazione del ciclo TCA, l ossalacetato
19 5) IL SUCCINIL-CoA E UN COMPOSTO AD ALTO CONTENUTO ENERGETICO CHE IN UNA REAZIONE CATALIZZATA DALLA SUCCINIL-CoA SINTETASI PRODUCE ATP CON UNA FOSFORILAZIONE AL LIVELLO DEL SUBSTRATO E SUCCINATO Il succinil-coa è un composto ad alto livello energetico la cui energia potenziale viene utilizzata per promuovere la formazione di un composto ad alta energia Avviene il processo noto come la fosforilazione al livello del substrato Negli animali non viene prodotto ATP, ma guanosina-trifosfato (GTP) che può essere facilmente trasformata in ATP, mediante scambio di un gruppo fosforico
20 6) IL SUCCINATO ATTRAVERSO UNA DEIDROGENAZIONE OPERATA DALLA SUCCINATO DEIDROGENASI FAD-DIPENDENTE VIENE OSSIDATO A FUMARATO CON PRODUZIONE DI FAD. IN REALTA IL VERO ACCETTORE DI ELETTRONI E RAPPRESENTATO DALL UBICHINONE Si tratta di una reazione di deidrogenazione FADdipendente di due atomi di carbonio con formazione di un doppio legame.
21 La presenza del FAD è legata al fatto che esso è un agente ossidante più potente del NAD in quanto il legame da ossidare è quello C-C L enzima succinato deidrogenasi è saldamente legato alla membrana mitocondriale interna Questa localizzazione è importante in quanto il FAD ridotto deve essere riossidato perché l enzima possa agire di nuovo. La riossidazione avviene grazie all interazione con il sistema di trasporto elettronico mitocondriale, anch esso legato alla membrana
22 7) ATTRAVERSO UNA IDRATAZIONE IL FUMARATO DA IL MALATO AD OPERA DELL ENZIMA FUMARASI La reazione consiste di una idratazione catalizzata dalla fumarasi
23 8) IL MALATO VIENE QUINDI OSSIDATO A OSSALACETATO IN UNA REAZIONE CHE COINVOLGE L ENZIMA MALATO DEIDROGENASI CON RIDUZIONE DEL NAD Il ciclo viene completato dalla deidrogenazione NAD-dipendente del malato ad ossalacetato. La reazione è fortemente endoergonica, ma procede verso destra per l azione della citrato sintasi che mantiene i livelli di OAA estremamente bassi
24 STECHIOMETRIA ED ENERGETICA DEL CICLO TCA Il ciclo inizia con l addizione di un frammento a due atomi di C (ACETIL-CoA) ed un accettore a 4 atomi di C (OAA). Durante il ciclo i due atomi di C vengono rimossi sotto forma di CO 2 ed il citrato viene ulteriormente metabolizzato. La reazione chimica bilanciata è la seguente: Acetil-CoA + 3 H 2 O + 3 NAD + + FAD + ADP + Pi 2 CO NADH + 3 FADH 2 + CoA-SH + ATP QUINDI DA UNA MOLE DI GLUCOSIO CHE PRODUCE DUE MOLI DI ACETIL-CoA SI HANNO: 4 molecole di CO 2, 6 molecole di NADH + H + e 2 molecole di FADH 2 e 2 moli di ATP

25
26 Regolazione del ciclo TCA Ci sono una serie di siti che controllano il ciclo TCA L andamento regolare del ciclo presuppone un input continuo di carbonio, acetil-coa derivato solitamente dal piruvato o dagli acidi grassi, anche se possono essere utilizzati gli amminoacidi e acidi organici Il controllo può essere esercitato quindi sia dall approvvigionamento che dalla velocità di ingresso di questi substrati nei mitocondri Inoltre molti enzimi coinvolti nel ciclo TCA sono regolati da vari metaboliti

27

28
29 Le reazioni catalizzate dal complesso della - chetoglutarato deidrogenasi e dalla citrato sintasi sono irreversibili. L attività di entrambi questi enzimi nei mitocondri è regolata dalla quantità della guanosina nucleotide. La -chetoglutarato deidrogenasi è inibita anche dal succinil-coa, un intermedio del ciclo.
30 L inibizione della citrato sintasi dovuta all ATP è stata spiegata come effetto secondario dell aumento di ATP che causa un inibizione di tipo feed-back della succinil-coa sintetasi, che provoca un aumento nella concentrazione di succinil-coa (che inibisce la -chetoglutarato deidrogenasi) La succinil-coa sintasi catalizza la fosforilazione reversibile, a livello di substrato, dell ADP usando l elevata energia libera di idrolisi del legame tioestere nel succinil-coa
31 La formazione del succinato è dunque strettamente associata alle concentrazioni relative di ATP e ADP Alte concentrazioni di ADP stimolano la formazione del succinato mentre alte concentrazioni dell ATP lo inibiscono Quindi la concentrazione degli adenosina e guanosina nucleotidi influisce enormemente sull attività di piruvato deidrogenasi, citrato sintasi, -chetoglutarato deidrogenasi e succinil-coa sintetasi e dunque è particolarmente importante per controllare il flusso del carbonio del ciclo Anche la succinato deidrogenasi esiste in forma attiva ed inattiva; la forma attiva è ottenuta grazie all interazione con vari metaboliti quali lo stesso succinil-coa, l ubichinone ridotto, l ATP, il succinato, alcuni anioni ed un basso ph
32 Comunque, nonostante l attività enzimatica sia certamente influenzata da questi prodotti, il loro significato fisiologico riguardo alla regolazione del ciclo nei mitocondri non è ancora ben chiarito Alcuni enzimi del ciclo tra cui la piruvato deidrogenasi e la isocitrato deidrogenasi sono soggetti ad inibizioni allosteriche dovute all aumento del NADH La malato deidrogenasi che catalizza la reazione di conversione dal malato ad ossalacetato è inibita sia dal NADH che dall ossalacetato Anche se l attività di molti enzimi del ciclo TCA può essere alterata in vitro cambiando la concentrazione dei metaboliti, il significato di tale attività in vivo è dubbio

33
34 REAZIONI (O VIE METABOLICHE) IN CUI SI UTILIZZANO GLI INTERMEDI DEL CICLO DI TCA Il ciclo TCA è una via anfibolica serva cioè ai processi anabolici e catabolici.gli intermedi del ciclo TCA sono implicati in numerosi altri pathway metabolici ACETIL-CoA - beta-ossidazione (ossidazione dei grassii - biosintesi dei grassi - degradazione della lisina, valina, leucina ed isoleucina - metabolismo fenilanina
35 -CHETO GLUTARATO SUCCINIL-CoA SUCCINATO FUMARATO OSSALACETATO - biosintesi della lisina - metabolismo acido ascorbico - metabolismo glutammato - metabolismo propionato - metabolismo fenilalanina - degradazione valina, leucina ed isoleucina - biosintesi del butanoato - metabolismo tirosina - ciclo dell urea - metabolismo arginina e prolina - metabolismo tirosina - metabolismo gliossilato - metabolismo glutammato ed aspartato - gluconeogenesis

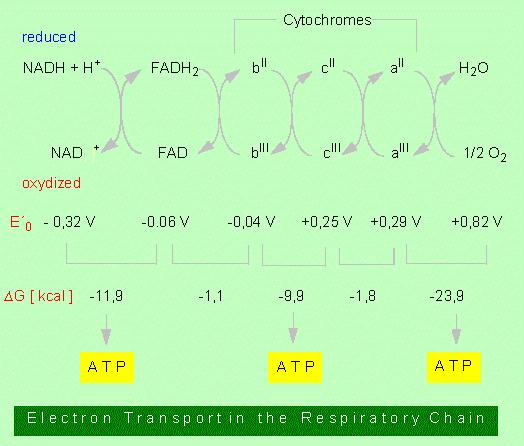
36
37 SEQUENZE ANAPLEROTICHE Il ciclo TCA funge anche come importante fonte di intermedi biosintetici. Queste vie tendono a sottrarre carbonio dal ciclo utilizzando gli intermedi Ad esempio, l OAA e l -CHETOGLUTARATO sono chetoacidi che portano alla sintesi di amminoacidi (aspartato e glutammato) La sottrazione di intermedi dal ciclo determinerebbe il venir meno del procedere del ciclo stesso se non esistessero processi in grado di ripristinare le riserve di intermedi del ciclo stesso. Queste vie sono più propriamente definite ANAPLEROTICHE (= riempimento)
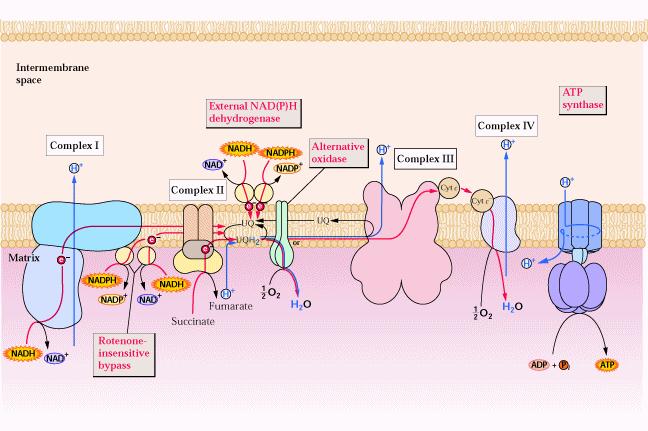
38 a) Ripristino dell OAA In piante e batteri l OAA viene direttamente prodotto dal PEP in una reazione catalizzata dalla FOSFOENOLPIRUVATO CARBOSSILASI:
39 b) Enzima malico Questo enzima è noto anche con il nome di MALATO DEIDROGENASI NADP-DIPENDENTE
40 La catena respiratoria Durante la glicolisi e il ciclo TCA si formano NADH e FADH 2 In condizioni aerobiche è necessaria la loro riossidazione affinché le due vie metaboliche non si blocchino La riossidazione avviene nella membrana interna del mitocondrio, attraverso una serie di carriers che costituiscono una catena di trasporto elettronico nota con il nome di CATENA RESPIRATORIA

41 L ossidazione del piruvato, l ossidazione dei grassi, l ossidazione degli amminoacidi ed il ciclo dell acido citrico avvengono tipicamente nella matrice mitocondriale Sappiamo che il mitocondrio è costituito da una membrana esterna, da una membrana interna, dallo spazio intermembrana e dalla matrice. La membrana interna è ripiegata a formare delle creste
42 Ma indipendentemente da dove avvengono, tutte le ossidazioni portano alla produzione di trasportatori di elettroni ridotti, soprattutto NADH La maggior parte di questo NADH viene riossidato, con concomitante produzione di ATP, attraverso la catena degli enzimi facenti parte della catena respiratoria, saldamente ancorata alla membrana interna E proprio sulla membrana interna che sono dislocati i diversi complessi responsabili della riossidazione del NADH e della contemporanea produzione di ATP

43 L effetto principale della catena è il trasporto di elettroni dal NADH e dal FADH 2 sino all O 2, che costituisce l accettore finale, per formare H 2 O NADH + H + + ½O 2 H 2 O + NAD + FADH 2 + ½O 2 H 2 O + NAD +
 queste reazioni redox sono fortemente esoergoniche ΔG 0 = - n FΔE 0 = -2(96,5)[0,82-(-0,32)] = -220 kj mol -1 Gran parte dell energia disponibile viene utilizzata per la formazione di un")
44 A causa della grande differenza tra i potenziali di ossidoriduzione del donatore (NADH + H + o ubichinone ridotto; ΔE 0 = V) e l accettore (O 2 ΔE 0 = V) queste reazioni redox sono fortemente esoergoniche ΔG 0 = - n FΔE 0 = -2(96,5)[0,82-(-0,32)] = -220 kj mol -1 Gran parte dell energia disponibile viene utilizzata per la formazione di un gradiente protonico che viene a sua volta usato dall ATP sintasi per produrre ATP PER OGNI NADH OSSIDATO SI FORMANO 3 ATP MENTRE PER OGNI FADH2 OSSIDATO SI OTTENGONO 2 ATP
45 Componenti della catena di trasporto elettronico mitocondriale La catena di trasporto degli elettroni avviene attraverso 4 complessi (complessi I, II, III e IV) che si trovano sulla membrana interna e molecole di trasferimento mobili quali l ubichinone (coenzima Q) e il citocromo c Il complesso II è in realtà la succinato deidrogenasi, che in realtà appartiene al ciclo del citrato L ATP sintasi viene denominata complesso V, anche se non partecipa al trasporto degli elettroni
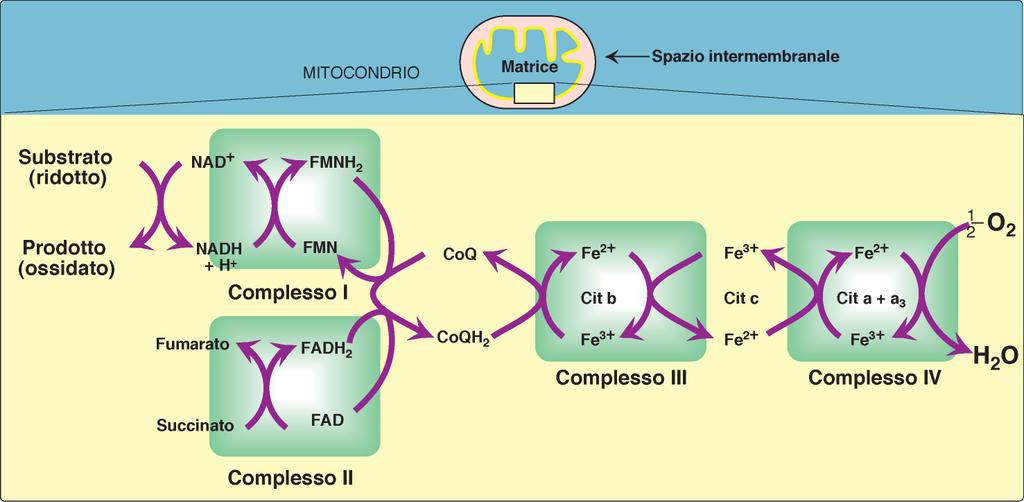
46
47
48
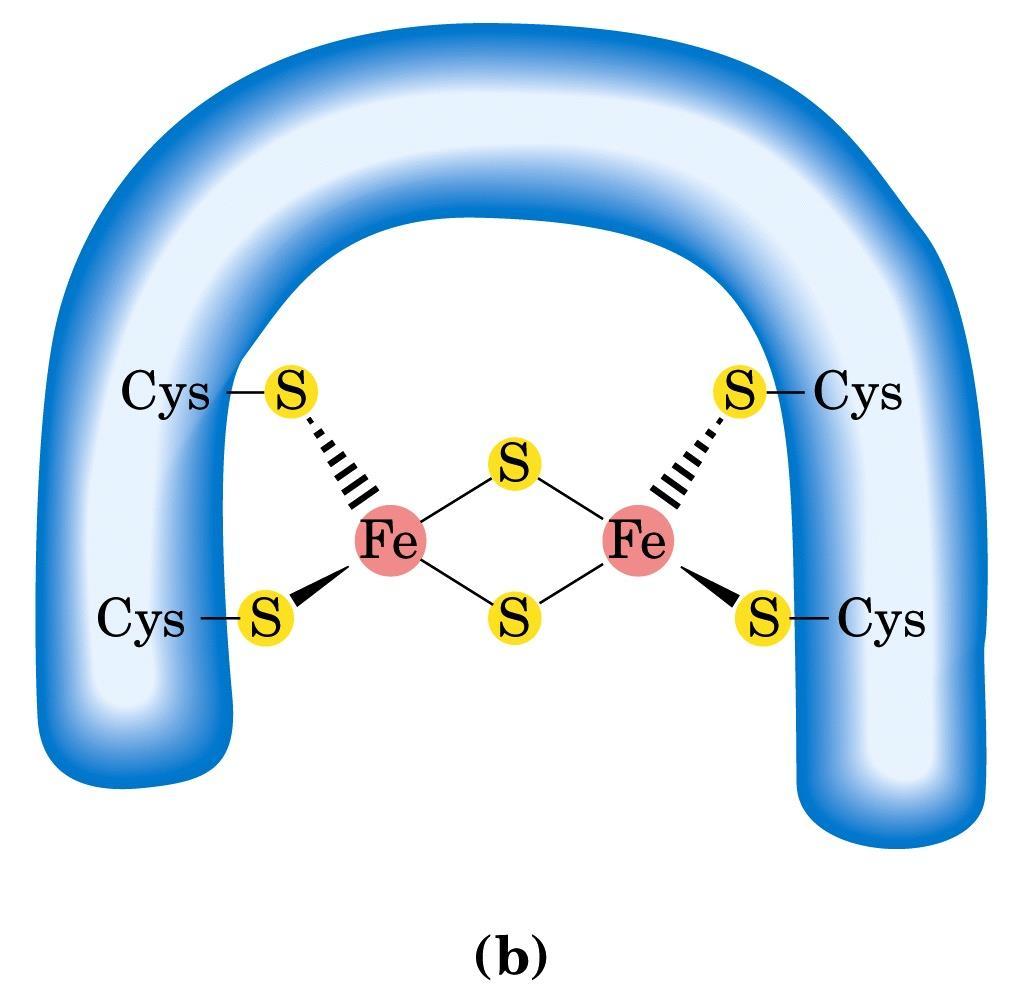
49 Tutti i complessi della catena respiratoria sono formati da numerose subunità polipeptidiche e contengono una serie di coenzimi redox legati alle proteine. Tra questi abbiamo: 1) FLAVOPROTEINE Le deidrogenasi che determinano il movimento degli elettroni nella catena respiratoria sono FLAVOPROTEINE. La NADH DEIDROGENASI che catalizza l ossidazione del NADH è un enzima legato alla membrana che contiene flavinmononucleotide (FMN) come gruppo prostetico 2) PROTEINE FERRO-ZOLFO Differiscono dai citocromi in quanto non contengono il gruppo eme, anche se contengono ferro, il quale è localizzato tra lo zolfo della cisteina e lo zolfo inorganico a formare un complesso 3) UBICHINONI Carrier mobile di elettroni solubile ai lipidi

50 Fe-S proteine
51

52

53 Complesso I: NADH deidrogenasi E costituito da 26 polipeptidi e 7 centri ferro-zolfo. Catalizza il trasporto di elettroni dal NADH al gruppo prostetico FMN della NADH deidrogenasi e quindi all ubichinone. Gli elettroni possono essere introdotti nella catena respiratoria in punti diversi
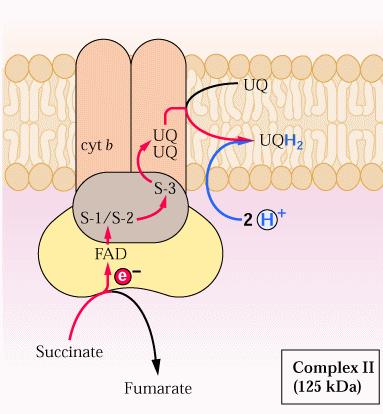
54 Complesso II: Succinato deidrogenasi Anche gli elettroni che derivano dall ossidazione del succinato, dall acetil-coa e da altri substrati vengono trasferiti all ubichinone rigenerando il FAD legato ad un enzima; il processo è mediato dal complesso II o da altre deidrogenasi mitocondriali Il complesso II è noto come succinato-ubichinone ossidoriduttasi ed è legato alla succinato deidrogenasi che partecipa al ciclo TCA. Il complesso contiene proteine Fe-S che accettano elettroni dal FADH 2, nella succinato deidrogenasi. Gli elettroni vengono quindi nuovamente trasferiti all ubichinone. Il trasferimento di elettroni è accompagnato dall assorbimento di 2H + della matrice
55
, tutti legati al complesso, riduce la piccola emoproteina citocromo")
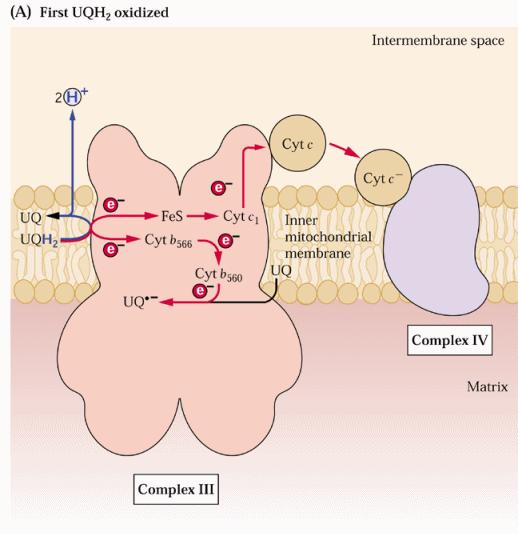
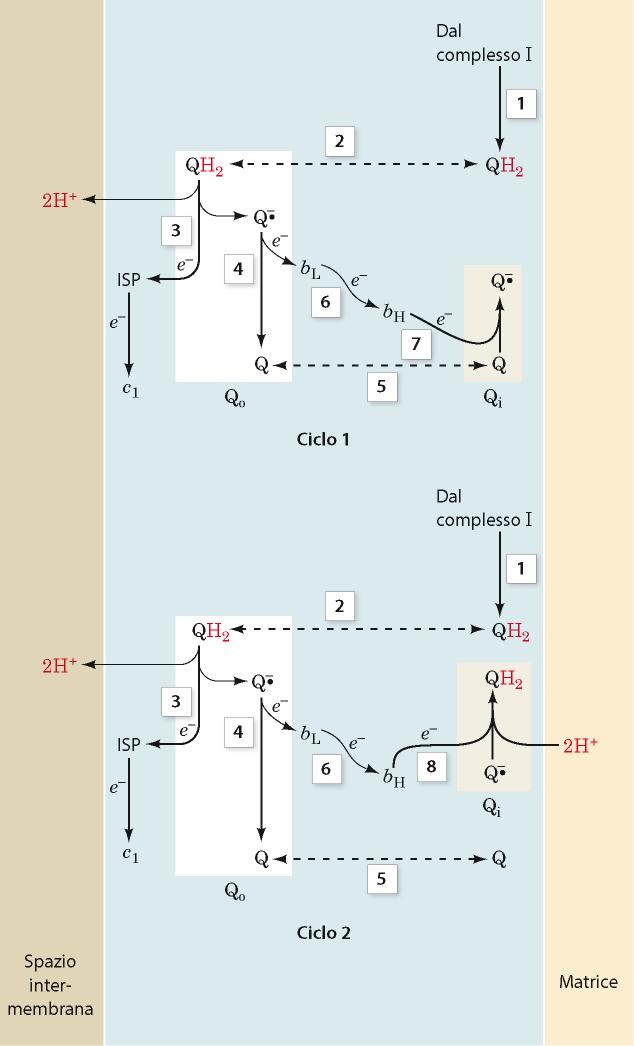
56 Complesso III: citocromo B, c 1 Questo complesso è noto come ubichinone citocromo C ossidoriduttasi L ubichinone ridotto trasferisce i suoi elettroni al complesso III che, a sua volta, attraverso i due gruppi eme di tipo b, un centro Fe/S e un gruppo eme di tipo c (citocromo c 1 ), tutti legati al complesso, riduce la piccola emoproteina citocromo c

57
58 Complesso IV: citocromo ossidasi La citocromo c ossidasi contiene due ioni rame (Cu A e Cu B ) e i gruppi eme a e a 3 attraverso i quali gli elettroni arrivano finalmente all ossigeno Il complesso catalizza il trasferimento di 4 elettroni. Ciò è importante in quanto rende possibile la completa riduzione dell O 2 ad H 2 O senza la generazione di intermedi tossici come l H 2 O 2 Questo complesso utilizza protoni nella matrice e trasferisce protoni nello spazio intermembrana

59
60 Come abbiamo visto, i complessi, compreso il V, sono tutti proteine integrali della membrana interna del mitocondrio e non sono in contatto diretto tra loro L ubichinone si muove liberamente nella membrana a causa della sua lunga catena laterale non polare Il citocromo c invece è solubile in acqua e si lega alla superficie esterna della membrana interna
61
62
63
64 Trasporto elettronico e sintesi dell ATP Abbiamo detto che il trasporto di elettroni nella catena respiratoria determina la liberazione di energia, la quale viene conservata come gradiente protonico TRANSMEMBRANA che è accoppiato alla sintesi dell ATP Si crea un gradiente protonico che determina una variazione del ph tra matrice e spazio intermembrana. Infatti: 1) La matrice è ALCALINA rispetto allo spazio intermembrana 2) I protoni rilasciati sono carichi positivamente per cui determinano una carica positiva nello spazio intermembrana
65 Tutto ciò determina un GRADIENTE ELETTROCHIMICO attraverso la membrana interna definito come FORZA MOTRICE PROTONICA (PMF) Questa PMF è mantenuta anche perché la membrana interna è impermeabile ai protoni che quindi non rientrano nella matrice L energia della PMF è incanalata attraverso un complesso enzimatico nella membrana interna. Il complesso è noto come F 0 F 1 -ATP SINTASI (complesso V) e canalizza la sintesi di ATP a partire da ADP e Pi L unità F 1 contiene il sito attivo per la sintesi di ATP Quando l unità è isolata dalla membrana mitocondriale non determina la sintesi di ATP ma catalizza l idrolisi di ATP in ADP+Pi

66
67 La produzione di ATP Si pensa che 10 siano i protoni trasportati attraverso la membrana per ogni molecola di NADH ossidata Sappiamo che: 3 H + determinano la sintesi di una mole di ATP Inoltre 1 H + è coinvolto nel trasporto di ADP + Pi nella matrice e dell ATP nel citoplasma
68 Quindi le molecole di ATP sintetizzate ogni NADH ossidato sono 2,5--3,3 Il trasporto di elettroni dal FADH 2 all O 2 coinvolge 6 protoni, quindi 1,5--2 moli di ATP Si considera comunque genericamente che sono 3 le moli di ATP prodotte per ogni NADH ossidato e 2 per mole di FADH 2 ossidato
69 In realtà non tutto il NADH è prodotto nella matrice (vedi GLICOLISI) e la membrana interna mitocondriale è impermeabile al NADH Questo problema è ovviato con il sistema navetta (SHUTTLE) Nella malato-aspartato navetta, l ossalacetato è ridotto a malato nel citoplasma dalla MALATO DEIDROGENASI Durante questa reazione il NADH è ossidato a NAD + Il malato viene quindi trasportato nella matrice attraverso un carrier di membrana che scambia il malato citoplasmatico con l α chetoglutarato
70 Una volta nella matrice il NADH è rigenerato dalla ossidazione del malato ad ossalacetato attraverso la malato deidrogenasi Tale navetta è completata da una reazione di transamminazione in cui l α-chetoglutarato è convertito a glutammato e l ossalacetato in aspartato
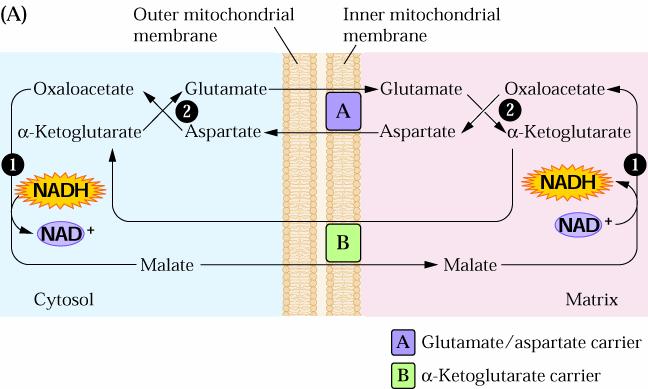
71
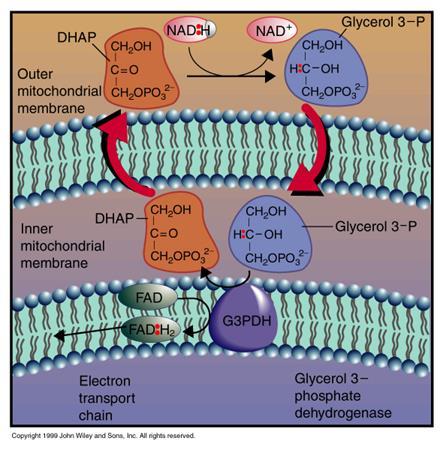
72


73
74 Resa energetica del metabolismo ossidativo Calcoliamo quanta energia si ottiene sotto forma di ATP dal catabolismo ossidativo completo del glucosio Di seguito vengono presentate le equazioni bilanciate per ognuna della tre vie degradative coinvolte GLICOLISI: Glucosio + 2ADP +2Pi +2NAD + 2 Piruvato + 2ATP + 2NADH +2H 2 O + 4H + COMPLESSO DELLA PIRUVATO DEIDROGENASI: 2Acetil-CoA + 6 H2O + 6NAD + + 2FAD + + 2ADP + Pi 4CO2 + 6NADH + 2FADH + 2CoA-SH + 2ATP REAZIONE COMPLESSIVA: Glucosio + 10NAD+ + 2FAD+ + 4H2O + 4ADP + 4 Pi 6CO2 +10NADH + 4H+ + 2FADH2 + 4ATP
75 Quindi i tre processi producono esattamente 4 moli di ATP, oltre 10 moli di NADH e due moli di FADH 2 Considerando che l ossidazione del NADH porta alla sintesi di 3 moli di ATP e quelle del FADH 2 a due moli di ATP si ha una resa totale di 38 moli di ATP Le cellule che utilizzano il sistema di trasporto a navetta del glicerolo fosfato devono pagare un costo energetico Gli elettroni del NADH citoplasmatico entrano nella catena respiratoria sotto forma di FADH 2 ; di conseguenza, la resa in ATP di ognuna di queste due molecole di NADH è pari a 2 e non a 3 Questo diminuisce la resa totale in ATP a 36

76

77

78
Modulo 14 Il ciclo di Krebs
 Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
Modulo 14 Il ciclo di Krebs Il destino ossidativo del piruvato In condizioni aerobie il piruvato viene decarbossilato ad acetato. L acetato è ossidato ulteriormente nel ciclo dell acido citrico. Una serie
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia.
 La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
Seminario. Ciclo di Krebs 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A
 Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico
 Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
Seconda Università degli Studi di Napoli DiSTABiF Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Prof. Antimo Di Maro Anno Accademico 2016-17 Lezione 14 Ciclo di Krebs L ossidazione
Cap.19. Ciclo di Krebs. o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico
 Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico
 Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
prodotto finale della glicolisi
 Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Piruvato: prodotto finale della glicolisi Può andare incontro a vari destini: -in condizioni anaerobie: il piruvato viene ridotto a lattato (fermentazione lattica) o ad etanolo (fermentazione alcolica)
Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la
 Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
Il ciclo di Krebs e la fosforilazione ossidativa
 Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
L ossidazione completa del glucosio da parte dell O 2. può essere suddivisa in due semi-reazioni
 L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa
 e Fosforilazione ossidativa") Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
6 H 2. con G=-686 kcal/mole di H 12 O 6 O + 6 CO O 2. glucosio La respirazione avviene in tre stadi principali; ognuno di questi
 La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
FADH ADP + Pi ATP...29 AG ' = -30,5 kj/mol...29 Resa di ATP per l ossidazione completa del glucosio...31
 FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
scaricato da www.sunhope.it
 CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
scaricato da 1
 scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
Come le cellule traggono energia dal cibo: produzione di ATP
 Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale
 Stechiometria della reazione finale") Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
Principali vie di utilizzo del Glucosio
 Principali vie di utilizzo del Glucosio DESTINO DEL PIRUVATO Il piruvato, può seguire due vie. La via fermentativa La via della respirazione cellulare In presenza di ossigeno, il piruvato, viene ossidato
Principali vie di utilizzo del Glucosio DESTINO DEL PIRUVATO Il piruvato, può seguire due vie. La via fermentativa La via della respirazione cellulare In presenza di ossigeno, il piruvato, viene ossidato
INTRODUZIONE AL METABOLISMO
 INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
Capitolo 6 La respirazione cellulare
 Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
FOSFORILAZIONE OSSIDATIVA
 FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
GLUCOSIO. Vena porta DIGESTIONE DEI DISACCARIDI. Enzimi parete intestinale
 DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
DIGESTIONE DELL AMIDO AMIDO Amilasi salivare DESTRINE Amilasi pancreatica MALTOSIO Enzimi parete intestinale GLUCOSIO Vena porta DIGESTIONE DEI DISACCARIDI DISACCARIDI Enzimi parete intestinale MONOSACCARIDI
Regolazione del Ciclo di Krebs 8 reazioni
 Regolazione del Ciclo di Krebs 8 reazioni Stechiometria del Ciclo Acetil-CoA + 3NAD + + FAD + GDP + Pi + 2H 2 O 2 CO 2 + HS-CoA + 3NADH + 2H + + FADH2 + GTP (GTP ATP) 1 NADH 2,5 ATP 1 FADH 2 1,5 ATP Per
Regolazione del Ciclo di Krebs 8 reazioni Stechiometria del Ciclo Acetil-CoA + 3NAD + + FAD + GDP + Pi + 2H 2 O 2 CO 2 + HS-CoA + 3NADH + 2H + + FADH2 + GTP (GTP ATP) 1 NADH 2,5 ATP 1 FADH 2 1,5 ATP Per
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
Introduzione allo studio del metabolismo Parte I
 Introduzione allo studio del metabolismo Parte I FOTOTROFI Le piante ed alcuni tipi di batteri ricavano l energia libera dal Sole mediante la fotosintesi, un processo in cui l energia luminosa è convertita
Introduzione allo studio del metabolismo Parte I FOTOTROFI Le piante ed alcuni tipi di batteri ricavano l energia libera dal Sole mediante la fotosintesi, un processo in cui l energia luminosa è convertita
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
NADH FADH 2 (trasportatori ridotti di elettroni) Catena respiratoria (trasferimento degli
 Catena respiratoria (trasferimento degli") NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
BIOCHIMICA e BIOTECNOLOGIE degli ALIMENTI
 Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2016-17 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Seconda Università degli Studi di Napoli DiSTABiF Anno Accademico 2016-17 Corso di Laurea Magistrale in SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA Insegnamento di BIOCHIMICA e BIOTECNOLOGIE degli
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore)
") Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
Attenzione: Lunedì 15 la lezione sarà dalle 14 alle 16 (al posto di L. Migliore) La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato
INTRODUZIONE AL METABOLISMO. dal gr. metabolè = trasformazione
 INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
I processi metabolici cellulari
 I processi metabolici cellulari 1 Metabolismo E l insieme delle reazioni chimiche che avvengono in una cellula o, più in generale, in qualsiasi organismo. Le sostanze coinvolte in tali reazioni sono dette
I processi metabolici cellulari 1 Metabolismo E l insieme delle reazioni chimiche che avvengono in una cellula o, più in generale, in qualsiasi organismo. Le sostanze coinvolte in tali reazioni sono dette
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA
 I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
catabolismo anabolismo
 Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
DESTINI METABOLICI DEL PIRUVATO
 DETINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DETINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
Anatomia biochimica di un mitocondrio
 Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Prof. Maria Nicola GADALETA DISPENSA N. 23
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA FOSFORILAZIONE OSSIDATIVA 4
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
Biosintesi dei carboidrati
 Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Utilizzo del glucosio: la glicolisi
 Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA
 Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre
 Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre molecole, anche chimicamente simili al substrato. Saturabilità:
Enzimi = catalizzatori di una reazione biochimica Caratteristiche: Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre molecole, anche chimicamente simili al substrato. Saturabilità:
Ciclo dell acido citrico
 Ciclo dell acido citrico Il metabolismo ossidativo dei carburanti metabolici I gruppi acetili derivati da carboidrati, amminoacidi e acidi grassi entrano nel ciclo dell acido citrico dove vengono ossidati
Ciclo dell acido citrico Il metabolismo ossidativo dei carburanti metabolici I gruppi acetili derivati da carboidrati, amminoacidi e acidi grassi entrano nel ciclo dell acido citrico dove vengono ossidati
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico
 Seconda Università degli Studi di Napoli DiSTABiF Prof. Antimo Di Maro Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Anno Accademico 2015-16 Lezione 15 Catena di trasporto degli
Seconda Università degli Studi di Napoli DiSTABiF Prof. Antimo Di Maro Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Anno Accademico 2015-16 Lezione 15 Catena di trasporto degli
Il metabolismo cellulare
 Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Il metabolismo microbico
 Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
ADP + HPO 3, NAD +, NADP +, FAD ATP, NADH, NADPH, FADH 2 ENERGIA CHIMICA
 MACROMOLECOLE CELLULARI Proteine Polisaccaridi Lipidi Acidi nucleici ADP + HPO 3 2-, NAD +, NADP +, FAD Metaboliti complessi RICCHI DI ENERGIA Carboidrati Lipidi Proteine ANABOLISMO (biosintesi) MOLECOLE
MACROMOLECOLE CELLULARI Proteine Polisaccaridi Lipidi Acidi nucleici ADP + HPO 3 2-, NAD +, NADP +, FAD Metaboliti complessi RICCHI DI ENERGIA Carboidrati Lipidi Proteine ANABOLISMO (biosintesi) MOLECOLE
IL METABOLISMO ENERGETICO
 IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
2 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA
 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE
 BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia
 I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia Edizione 2016-2017 Metabolismo: equilibrio e omeostasi dei processi cellulari Lezione: Le principali vie metaboliche, il
I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia Edizione 2016-2017 Metabolismo: equilibrio e omeostasi dei processi cellulari Lezione: Le principali vie metaboliche, il
ANABOLISMO DEI LIPIDI
 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una quota
COME VIENE METABOLIZZATA QUESTA MASSICCIA QUANTITA DI ENERGIA? Trasformazione di energia potenziale di riduzione. energia libera di fosforilazione
 Trasformazione di energia potenziale di riduzione in energia libera di fosforilazione Un uomo adulto produce in media abbastanza energia da sintetizzare ogni giorno una quantità di ATP pari al suo peso
Trasformazione di energia potenziale di riduzione in energia libera di fosforilazione Un uomo adulto produce in media abbastanza energia da sintetizzare ogni giorno una quantità di ATP pari al suo peso
Ciclo dell Acido Citrico
 Chimica Biologica A.A. 2010-2011 Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano -la glicolisi non è l unica via catabolica in grado di produrre energia Anzi, le
Chimica Biologica A.A. 2010-2011 Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano -la glicolisi non è l unica via catabolica in grado di produrre energia Anzi, le
Energia e metabolismi energetici
 Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO
 IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO ( Introduzione Metabolismo energetico Funzione Il metabolismo energetico è un attività coordinata cui partecipano
IL METABOLISMO ENERGETICO BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE GSCATULLO ( Introduzione Metabolismo energetico Funzione Il metabolismo energetico è un attività coordinata cui partecipano
BETA OSSIDAZIONE DEGLI ACIDI GRASSI
 Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
IL CICLO DELL ACIDO CITRICO
 IL CICLO DELL ACIDO CITRICO Il ciclo dell acido citrico rappresenta il processo centrale attraverso il quale vengono catabolizzati tutti i combustibili metabolici Per entrare nel ciclo, lo scheletro carbonioso
IL CICLO DELL ACIDO CITRICO Il ciclo dell acido citrico rappresenta il processo centrale attraverso il quale vengono catabolizzati tutti i combustibili metabolici Per entrare nel ciclo, lo scheletro carbonioso
DESTINI DEL PIRUVATO
 DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
Catabolismo = degradazione di composti per recuperare i loro. Anabolismo = Le vie anaboliche e cataboliche sono in relazione fra loro
 Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI
 METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
Il metabolismo cellulare
 Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Catabolismo = degradazione di composti per recuperare i loro. Anabolismo = Le vie anaboliche e cataboliche sono in relazione fra loro
 Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
Mitocondri. -sono visibili al MO (Ø 0,5 µ e lunghezza da 1 a 6 µ) -assenti nei batteri e presenti in tutte le cellule eucariotiche
 -assenti nei batteri e presenti in tutte le cellule eucariotiche") Mitocondri sono organelli deputati alla produzione dell energia, capaci di ossidare i prodotti dell assorbimento intestinale degradandoli e di convertire l energia liberata in ATP processo chiamato respirazione
Mitocondri sono organelli deputati alla produzione dell energia, capaci di ossidare i prodotti dell assorbimento intestinale degradandoli e di convertire l energia liberata in ATP processo chiamato respirazione
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Cap.16 GLICOLISI. Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP H 2 O 2 NAD + 2 NADH + 2H +
 Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
La respirazione cellulare
 La respirazione cellulare Lo zucchero ("alimento") prodotto nella fase oscura della fotosintesi viene ossidato nel mitocondrio e qui si libera energia metabolica sotto forma di ATP. ANABOLISMO e CATABOLISMO
La respirazione cellulare Lo zucchero ("alimento") prodotto nella fase oscura della fotosintesi viene ossidato nel mitocondrio e qui si libera energia metabolica sotto forma di ATP. ANABOLISMO e CATABOLISMO
Catabolismo = degradazione di composti per recuperare i loro. Anabolismo = Le vie anaboliche e cataboliche sono in relazione fra loro
 Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
AGISCONO NEL SISTEMA NAVETTA 2 ISOFORME DELL ENZIMA: UNA FORMA MITOCONDRIALE ( MEMBRANA MIT. INTERNA) FAD-dipendente
 FAD-dipendente") METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
Fonte diretta di Energia è l ATP.
 Una cellula compie tre tipi di lavoro: -Lavoro meccanico: movimenti muscolari.. -Lavoro di trasporto: trasporto contro gradiente -Lavoro chimico: assemblaggio di polimeri da monomeri Fonte diretta di Energia
Una cellula compie tre tipi di lavoro: -Lavoro meccanico: movimenti muscolari.. -Lavoro di trasporto: trasporto contro gradiente -Lavoro chimico: assemblaggio di polimeri da monomeri Fonte diretta di Energia
metano alcool metilico formaldeide acido formico CO 2
 -4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
-4-2 0 +2 +4 metano alcool metilico formaldeide acido formico CO 2 + ridotto + ossidato + ossidato Glicolisi 2 Glucosio Piruvato Glicolisi Glucosio + 2NAD + 2 piruvato + 2NADH + 2H + ΔG = -146 kj/mol 2ADP
Fosforilazione a livello del substrato
 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
Degradazione degli acidi grassi
 Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 27 Panoramica del ciclo dell acido citrico e sintesi di acetil-coa Concetti chiave: Il ciclo dell acido citrico èun processo
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 27 Panoramica del ciclo dell acido citrico e sintesi di acetil-coa Concetti chiave: Il ciclo dell acido citrico èun processo
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
ENERGIA LIBERA DI GIBBS (G)
") METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
Tipi di trasportatori di elettroni nella catena respiratoria
 Tipi di trasportatori di elettroni nella catena respiratoria 1. Flavoproteine 2. Proteine Ferro Zolfo 3. Citocromi 4. Citocromi contenenti Rame 5. Chinone: Coenzima Q All eccezione del coenzima Q, tutti
Tipi di trasportatori di elettroni nella catena respiratoria 1. Flavoproteine 2. Proteine Ferro Zolfo 3. Citocromi 4. Citocromi contenenti Rame 5. Chinone: Coenzima Q All eccezione del coenzima Q, tutti
FOSFORILAZIONE OSSIDATIVA
 FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
I processi mediante i quali le molecole biologiche vengono scisse e risintetizzate formano una rete di reazioni enzimatiche, complessa e finemente
 I processi mediante i quali le molecole biologiche vengono scisse e risintetizzate formano una rete di reazioni enzimatiche, complessa e finemente regolata, detta metabolismo dell organismo. Questa rete
I processi mediante i quali le molecole biologiche vengono scisse e risintetizzate formano una rete di reazioni enzimatiche, complessa e finemente regolata, detta metabolismo dell organismo. Questa rete
Ossidazione del glucosio
 Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
Il ciclo dell acido citrico
 Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Il ciclo dell acido citrico Il catabolismo di proteine, grassi e carboidrati avviene nelle tre fasi della respirazione cellulare Il piruvato viene ossidato ad acetil-coa e CO 2 La decarbossilazione
Il metabolismo: concetti di base
 Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche Il metabolismo: concetti di base Caratteristiche generali Strategie Trofiche Il metabolismo:
Corso di Laurea Magistrale in Ingegneria Biomedica Complementi di Chimica e Biochimica per le Tecnologie Biomediche Il metabolismo: concetti di base Caratteristiche generali Strategie Trofiche Il metabolismo:
Catabolismo = degradazione di composti per recuperare i loro. Anabolismo = Le vie anaboliche e cataboliche sono in relazione fra loro
 Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
Il METABOLISMO comprende la totalità dei processi attraverso i quali gli esseri viventi ricavano e utilizzano energia secondo i limiti imposti dalle leggi della termodinamica : Catabolismo = degradazione
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO)
") RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
Fosforilazione ossidativa
 Fosforilazione ossidativa H 2 2H + + 2e - NADH+H + NAD + + 2H + +2e - FADH 2 FAD + 2H + + 2e - ½O 2 + 2H + + 2e - H 2 O + Calore ½ O 2 + 2H + + 2e - H 2 O + ATP 1 La fosforilazione ossidativa avviene nella
Fosforilazione ossidativa H 2 2H + + 2e - NADH+H + NAD + + 2H + +2e - FADH 2 FAD + 2H + + 2e - ½O 2 + 2H + + 2e - H 2 O + Calore ½ O 2 + 2H + + 2e - H 2 O + ATP 1 La fosforilazione ossidativa avviene nella
IL METABOLISMO. Dal carbonio agli OGM Capitolo 2
 IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
Appunti di biologia IL CICLO DI KREBS. Giancarlo Dessì Licenza Creative Commons BY-NC-SA
 Appunti di biologia IL IL DI KREBS Giancarlo Dessì http://www.giand.it Licenza reative ommons BY-N-SA BY: Attribuzione - N: Non commerciale - SA: ondividi allo stesso modo IL DI KREBS detto anche IL DELL'AID
Appunti di biologia IL IL DI KREBS Giancarlo Dessì http://www.giand.it Licenza reative ommons BY-N-SA BY: Attribuzione - N: Non commerciale - SA: ondividi allo stesso modo IL DI KREBS detto anche IL DELL'AID
Fosforilazione ossidativa
 Chimica Biologica A.A. 2010-2011 Fosforilazione ossidativa Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Teoria Chemiosmotica ΔG = 2.3 RT (ph (matrice) -ph (spazio
Chimica Biologica A.A. 2010-2011 Fosforilazione ossidativa Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Teoria Chemiosmotica ΔG = 2.3 RT (ph (matrice) -ph (spazio
Cap.21 ANABOLISMO DEI LIPIDI
 Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
Cap.21 ANABOLISMO DEI LIPIDI Sintesi di Acidi Grassi I grassi depositati come riserva nel nostro organismo non provengono soltanto dai grassi alimentari normalmente i grassi alimentari rappresentano una
Funzioni dei nucleotidi
 Funzioni dei nucleotidi monomeri degli acidi nucleici esempi di altre funzioni ATP: moneta energetica GTP: fonte di energia nella sintesi proteica camp: secondo messaggero nella trasduzione del segnale
Funzioni dei nucleotidi monomeri degli acidi nucleici esempi di altre funzioni ATP: moneta energetica GTP: fonte di energia nella sintesi proteica camp: secondo messaggero nella trasduzione del segnale