Note sui messagg eri intracellulari (revisione 2005, distribuzione 2005) 1. Introduzione
|
|
|
- Alfonsina Turco
- 8 anni fa
- Visualizzazioni
Transcript
1 Note sui messagg eri intracellulari (revisione 2005, distribuzione 2005) 1. Introduzione Com é noto, nella larga maggioranza dei casi, il passaggio attraverso la membrana plasmatica dell infor mazione associata a molecole extracellular i, avviene tramite recettori di membrana che permettono di attraversare la membrana al segnale extracellulare, ma non alla molecola che lo veicola. Infatti, la risposta dei recettori di membrana all occupazione dei loro siti recettoriali si sostanzia in un cambiamento confor mazionale che induce le opportune modifiche dei parametr i intracellular i in modo diretto o indiretto. I recettor i possono infatti attivare una funzione effettr ice associata alla stessa molecola recettoriale, e si definiscono come di tipo 1; quando invece i recettori non hanno funzioni effettr ici dirette (detti di tipo 2), é necessar io che l infor mazione sia trasmessa agli effettor i interessati. In questo caso, il segnale chimico por tato dalle molecole extracellular i, e convertito in cambiamento confor mazionale del recettore, viene riconvertito in un ulteriore segnale chimico riconoscibile dai meccanismi intracellular i che debbono essere modificati. Questo riconoscimento avviene tramite una caratter istica cascata di reazioni che induce l aumento delle concentrazioni intracellular i di opportune molecole o ioni, i messaggeri intracellular i o secondi messaggeri. Pur non costituendo la totalità dei recettori di tipo 2, i recettori che utilizzano secondi messagger i sono tipicamente quelli accoppiati alle proteine G, cui si farà prevalentemente rifer imento. 2. Considerazioni generali 2.1. Schema comune Tr a l occupazione del sito recettoriale e l alterazione del comportamento cellulare esistono serie di proteine intracellular i che for mano una catena di trasmissione. La massima parte di queste proteine appartiene a due classi: proteine che vengono fosfor ilate da specifiche proteina chinasi e proteine che hanno un sito di legame per i nucleotidi guaninici e che scambiano GTP con GTP. In entrambi i casi viene aggiunto alla proteina un atomo di fosforo: tuttavia, il legame é covalente nel primo caso ma non nel secondo. In entrambi i casi, queste proteine causano tipicamente la fosfor ilazione di altre proteine, dando luogo a cascate di reazioni in genere estremamente complessa anche per la presenza di sistemi multipli interagenti. La successione degli eventi connessi con l attivazione di un effettore intracellulare ad opera dei secondi messaggeri può essere schematizzata come segue: 1. il segnale extracellulare giunge ad un sito recettoriale capace di riceverlo, inducendo un cambiamento confor mazionale della molecola del recettore, attivandolo. 2. Nella sua confor mazione attiva, il recettore interagisce con una proteina G; 3. L interazione con il recettore induce nella proteina G un primo cambiamento confor mazionale, che la mette in grado di interagire con un enzima (l enzima intermedio) facendolo passare nella sua confor mazione attiva. 4. L enzima intermedio attivato catalizza la trasfor mazione di un precursore nello specifico secondo messaggero; 5. Il secondo messaggero diffonde nel citoplasma fino a legarsi ad un effettore intermedio dando inizio ad una cascata di reazioni; 6. L ultimo effettore intermedio agisce sull effettore finale: ad esempio, una proteina chinasi fosfor ila l effettore finale citoplasmatico, por tandolo così nella sua confor mazione attiva. Va anche osservato che molto spesso i segnali esterni devono alterare l espressione di specifici geni; i segnali devono quindi essere trasmessi al nucleo, che può essere considerato l effettore finale. Come ripor tato oltre, questo schema può coincidere con i casi specifici in modo più o meno completo: nella fattispecie, gli eventi intermedi possono essere diversi da quelli schematizzati sopra: il secondo messaggero può attivare enzimi diversi da una proteina chinasi, operando modifiche regolative degli effettor i finali differenti dalla fosfor ilazione. In casi specifici funzionalmente rilevanti descritti più oltre, gli effettor i finali possono essere attivati direttamente dalle proteine G, oppure dai secondi messaggeri, come avviene nel caso dei canali ionici controllati dai nucleotidi ciclici. In questo caso, l occupazione di un recettore metabotròpo produce un effetto ionico anzichè metabolico. Inoltre, ilca 2+ intracellulare può essere mobilitato dalle sue riser ve non mitocondriali tramite specifici canali a controllo di ligando, i meglio noti dei quali sono -1-

2 recettor i per l IP 3 eicosì detti recettori per la rianodina. In fine, alcuni recettori con attività chinasica attivano direttamente l enzima intermedio, ossia senza l intervento di una proteina G Considerazioni funzionali L attivazione dei secondi messaggeri compor ta una serie di eventi metabolici che necessariamente inducono un ritardo più o meno rilevante tra l interazione del ligando con il sito recettoriale e l attivazione dell effettore finale. Al tempo stesso, propr io l esistenza di reazioni intermedie comporta anche un amplificazione in genere molto cospicua del segnale originale. Ciò significa che l occupazione di un singolo sito recettoriale extracellulare può attivare un gran numero, spesso nell ordine delle migliaia, di reazioni intracellular i. Pertanto, i secondi messaggeri trasmettono normalmente segnali in cui un ritardo nella trasmissione non é un fattore limitante, ma per i quali l amplificazione risultante costituisce un effetto positivo. Per fare qualche esempio, i recettor i nicotinici dell acetilcolina ad azione rapida sono di tipo 1 (recettori-canale), mentre quelli muscar inici, che mediano effetti metabolici a più lungo termine, agiscono tramite secondi messaggeri. Al contrar io, i recettor i adrenergici, le cui azioni sono sempre di tipo metabolico (e.g. degradazione del glicogeno, degradazione dei lipidi, secrezione e riassorbimento di liquidi...) agiscono solo tramite secondi messagger i. Va però notato come la trasduzione del segnale avvenga tramite secondi messaggeri anche nel caso dei segnali visivi e di quelli olfattor i. Si può supporre che, in questi casi, il vantaggio funzionale der ivante dall amplificazione del segnale sia maggiore dello svantaggio causato dal ritardo nella trasmissione. Questo é relativamente ridotto perchè la catena metabolica é particolar mente breve. Tuttavia, nel caso dei fotorecettor i, la lunghezza del tempo di trasduzione del segnale si traduce in mancanza di nitidezza nella percezione degli oggetti in movimento veloce e nella persistenza delle immagini retiniche Valenza funzionale Paragonato al grandissimo numero di molecole infor mazionali extracellular i conosciute, il numero di secondi messagger i noti é molto limitato: infatti, all esterno della cellula i segnali sono veicolati da moltissime differenti molecole (e il numero di quelle note continua ad aumentare); al contrar io, la trasmissione dell infor mazione intracellulare é affidata a solo poche classi di molecole. In più, le vie dei secondi messagger i sono estesamente incrociate, di modo che l attivazione di una comporta in genere anche l attivazione di altre. Tale organizzazione funzionale é sorprendente solo in parte. Infatti, le cellule degli organismi animali tendono ad essere for temente specializzate; inoltre, le funzioni cellulari attivabili dall ester no dipendono sostanzialmente dalla specificità dei recettori presenti nella membrana. In questi ter mini, i segnali esterni attivano, o inibiscono, le poche funzioni proprie della cellula, anzichè selezionarne una specifica entro un gran numero di funzioni potenzialmente attivabili. Infatti, ogni singola classe di cellule é dotata di un numero limitato di differenti tipi di recettori e la selezione tra i segnali extracellular i che giungono alla cellula viene appunto effettuata al livello di questi. Ciò vuol dire che vengono recepiti da ogni cellula solo quei segnali che sono portati da molecole infor mazionali per i quali la cellula stessa esprime recettor i. Quindi, le funzioni che vengono attivate sono funzione sia del tipo di recettori presenti che del tipo di risposta che la specifica cellula é programmata a for nire. Ad esempio, l acetilcolina stimola la contrazione del muscolo scheletrico legandosi ai recettori nicotinici della placca motrice, ma riduce le caratter istiche di contrazione del cuore legandosi ai recettori muscar inici delle cellule muscolar i cardiache. Com é noto, i primi sono canali cationici con azione depolarizzante; i secondi sono recettori di tipo 2 che attivano (tramite proteine G) i canali potassici di tipo M, la cui apertura deter mina iper polarizzazione. Sempre l acetilcolina legandosi a recettori muscar inici stimola la secrezione di cellule secernenti. In questo caso la differenza nella risposta cellulare é data dal tipo di risposta per la quale la cellula é programmata: le cellule muscolar i rispondono contraendosi, le cellule secernenti, appunto secernendo. Per tanto, i segnali extracellular i che giungono ai recettori accoppiati alle proteine G vengono in genere integrati in una risposta cellulare che non é di solito molto var ia. Nei termini qui sintetizzati, al limite un solo secondo messaggero (o due, uno eccitator io ed uno inibitorio) potrebbe essere sufficiente a coprire le necessità della cellula. Com é ovvio, la realtà é assai più complessa; tuttavia, la rappresentazione for nita da questo schema concettuale sembra in ragionevole accordo con la maggior parte dei dati noti. Va infine notato che la mancata identificazione del secondo messaggero relativo ad alcuni recettori fa -2-

3 supporre che possano esistere vie di trasmissione intracellulare ancora sconosciute. Nulla tuttavia induce oggi a pensare che si possa assistere in futuro ad una cospicua proliferazione del numero dei secondi messagger i, così come é avvenuto ed ancora in par te avviene nel caso delle molecole infor mazionali extracellular i Classi Sono oggi conosciute var ie vie di trasmissione intracellulare che utilizzano differenti secondi messaggeri, come descritto più oltre. Questi sono: 1. gli inositolo fosfati; questa via, che utilizza come messaggero finale gli ioni Ca 2+, agisce tramite i due messaggeri inter medi inositolo(1,4,5)-tr ifosfato (IP 3 ) e inositolo (1,3,4,5)-tetrafosfato (IP 4 ), polari, nonchè il diacilglicerolo (DAG), apolare. 2.L AMP ciclico (camp), anche esso spesso coinvolto nella liberazione di ioni Ca 2+ ;3.Ilguanosina monofosfato ciclico (cgmp) che ha funzioni di secondo messaggero, sia autonome che in relazione alla liberazione di ioni Ca 2+. Hanno funzioni di secondi messaggeri anche (4) l acido arachidonico e alcuni suoi metaboliti, prodotti da fosfolipasi attivate da proteine G che utilizzano come substrato var i fosfolipidi e (5) l ossido nitrico (o anidride nitrica, NO) (e probabilmente anche l ossido di carbonio, CO) che agiscono come secondi messaggeri e come trasmettitor i, sopratutto nell ambito del sistema nervoso centrale. Infine (6) gli ioni Ca 2+ giocano un ruolo fondamentale nella trasmissione intracellulare delle infor mazioni, sia in modo autonomo che in qualità di messaggeri finali di eventi intracellular i ("secondi secondi messaggeri"), come descritto sotto. 3. Ioni Ca 2+ In quanto ioni, le concentrazioni intracellular i degli ioni Ca 2+ non possono ovviamente essere modificate per sintesi e demolizione, com é il caso degli altri secondi messaggeri. In loro vece, vengono attivati var i meccanismi di traspor to verso e dal citoplasma: in generale, le concentrazioni calciche citoplasmatiche vengono tenute molto più basse di quelle extracellular i (circa 1x10-7 M) dall azione di numerose proteine di traspor to che operano in direzione extracitoplasmatica, mentre aumentano a seguito dell apertura di canali selettivi che permettono il passaggio seguendo il ripido gradiente elettrochimico di questi ioni. Per tanto, questi meccanismi traspor tano gli ioni Ca 2+ attraverso la membrana plasmatica, dentro e fuori lacellula, e attraverso le membrane interne da e verso organelli intracellular i (sopratutto reticolo sarcoplasmatico, calcisomi e mitocondri). A seguito dell apertura dei canali calcici, le concentrazioni intracellular i di Ca 2+ possono aumentare anche di 100 volte, for nendo il segnale finale che induce funzioni cellulari come la contrazione muscolare, la secrezione, la liberazione di neurotrasmettitor i. Infine, moltissimi degli effetti intracellular i degli ioni Ca 2+ sono mediati dalla fosfor ilazione di proteine, catalizzata da una famiglia di proteina chinasi calcio-calmodulina dipendenti (chinasi CaM). Queste chinasi sono attive solo in presenza di oppor tune concentrazioni calciche: infatti, quando le concentrazioni calciche citoplasmatiche aumentano, gli ioni Ca 2+ si possono legare alla calmodulina (vedi sotto); a siti calcici occupati, questa cambia confor mazione e diventa in grado li legarsi alla chinasi M che, a sua volta, cambia confor mazione assumendo la confor mazione nella quale il sito catalitico é attivo. Data l importanza e l ubiquità degli ioni Ca 2+ imeccanismi di traspor to coinvolti sono moltissimi, ed una loro descr izione é del tutto al di fuori dei fini presenti. In termini molto generici, rapidi aumenti delle concentrazioni citoplasmatiche di ioni Ca 2+ sono in gran par te deter minati dall aper tura dei canali calcici della membrana cellulare e degli organelli di deposito calcico, mentre la riduzione della concentrazione calcica citoplasmatica avviene mediante meccanismi di traspor to attivo diretto o indiretto. Ingenerale, gli ioni Ca 2+ entrano nella cellula attraverso la membrana plasmatica tramite canali a controllo di potenziale, entrano nel citoplasma da organelli intracellular i attraverso canali sia a controllo di potenziale che a controllo di ligando e vengono rimossi dal citoplasma da traspor tatori multipli. Spesso il traspor to iniziale é operato da traspor tatori abassa affinità ed alta capacità di traspor to; quando le concentrazioni si sono ridotte tanto che i traspor tatori a bassa affinità non legano più una quntità significativa di ioni, entrano in gioco altri traspor tatori, caratter izzati da elevata affinità ma da bassa capacità di traspor to. In questo modo, il recupero delle concentrazioni basali può avvenire con la rapidità richiesta dalle esigenze metaboliche e le concentrazioni calciche citoplasmatiche possono essere mantenute ai bassi livelli necessari. -3-

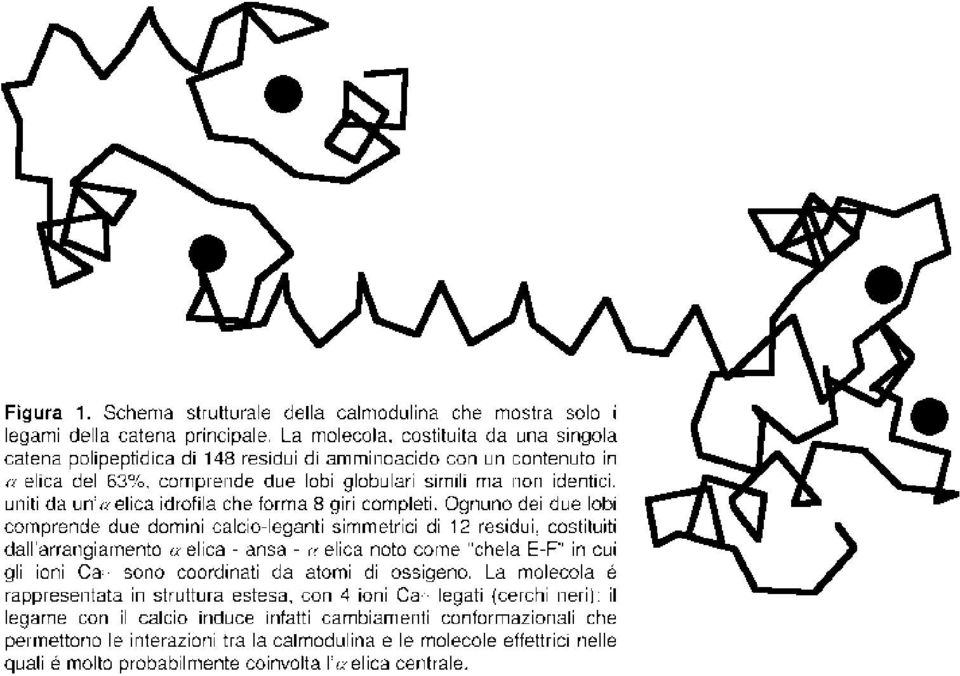
4 3.1. Meccanismi d azione Gli ioni Ca 2+ esercitano la loro azione legandosi a specifiche proteine endocellulari che presentano in genere domini calcio-leganti di struttura caratter istica, noti sotto il nome di "chele E-F", quali l ubiquitaria calmodulina, o la sua for ma specifica del muscolo scheletrico, latroponina C. Lacalmodulina, che é largamente presente in tutte le cellule animali (costituisce, inmedia, circa l 1% della massa proteica totale), si trova sia libera che come subunità regolatrice di alcuni enzimi, ad esempio della chinasi A attivata dall camp, odella fosfor ilasi-chinasi che attiva laglicogeno sintetasi nel muscolo scheletrico. Lacalmodulina (Figura 1)éuna proteina for mata da due domini calcio-leganti strutturalmente simili ma dotati di differente affinità per gli ioni Ca 2+,ognuno dei quali può legare due ioni (4 in totale). Questi domini sono separati da un α elica idrofila piuttosto lunga che, molto verosimilmente, écoinvolta nelle interazioni con almeno alcune delle proteine effettr ici calmodulina-dipendenti. L occupazione dei siti Ca 2+ -leganti induce nella molecola cambiamenti confor mazionali, presumibilmente multipli, a seguito dei quali la calmodulina può interagire con gli effettor i. Inoltre, dato che come detto sopra laconcentrazione intracellulare basale degli ioni Ca 2+ viene mantenuta a livelli molto bassi, le molecole di calmodulina presenti nel citoplasma dovrebbero essere in grado di assorbire le var iazioni delle concentrazioni di ioni Ca 2+ conseguenti all apertura dei canali calcici (sia della membrana plasmatica che intracellular i), contr ibuendo a mantenere costanti le concentrazioni citoplasmatiche di questi ioni. In effetti, eccetto casi specifici come il muscolo scheletrico, lamassima parte degli effetti metabolici degli ioni Ca 2+ émediata da proteine chinasi calcio/calmodulina-dipendenti (chinasi CaM) che sono attivate dal legame della calmodulina, che si stabilisce solo quando i siti calcio-leganti della calmodulina sono occupati. Chinasi CaM a specificità ristretta sono, adesempio, lachinasi delle catene leggere della miosina che regola la contrazione del muscolo liscio o la chinasi CaM II, un enzima a bassa specificità particolar mente concentrato nelle sinapsi. Nel caso di quest ultima, nelle sinapsi dei neuroni catecolamminergici il flusso di Ca 2+ entrante indotto dalla depolarizzazione (che apre canali calcici a controllo di potenziale) non solo interagisce con le proteine motrici inducendo lo spostamento delle vescicole verso la membrana presinaptica ma tramite appunto la chinasi CaM II fosfor ila, attivandola, la tirosina idrossilasi così aumentando la sintesi delle catecolammine. Vaancora osser vato che le chinasi CaM II nel suo stato attivato é in grado di autofosfor ilarsi: in queste condizioni rimane attiva anche in assenza di ioni Ca 2+ fino a quando non é defosfor ilata dalle fosfatasi citoplasmatiche, for nendo così un amplificazione dei segnali calcici Traspor to attraverso la membrana plasmatica Come detto, iltraspor to passivo diioni Ca 2+ attraverso la membrana plasmatica é garantito dall apertura di canali ionici. Tra i canali a controllo di ligando hanno un ruolo particolar mente impor tante nel rilascio di ioni Ca 2+ dai depositi intracellular i non mitocondriali i recettori per l IP 3 ed i recettori per la rianodina. Specificamente, nei tessuti eccitabili, in cui il potenziale di membrana va incontro a var iazioni par ticolarmente rapide, molto calcio entra nella cellula dall esterno tramite i canali a controllo di potenziale presenti nella membrana plasmatica; nelle cellule di altri tessuti il ruolo principale dovrebbe invece essere giocato dai canali a controllo di ligando. Va però osservato che in quasi tutte le cellule esiste uno sbilanciamento di potenziale transmembrana con interno negativoche facilita l ingresso di questi ioni Traspor to in direzione extracitoplasmatica Dopo l ingresso di ioni Ca 2+,lenor mali concentrazioni calciche vengono ripr istinate dall azione di opportune proteine vettr ici. E nota l esistenza di meccanismi che traspor tano il calcio dal citoplasma in direzione extracellulare, edialtr i che lo traspor tano verso specifici organelli. Dei traspor tatori degli ioni Ca 2+ in direzione extracellulare, leatpasi Ca 2+ dipendenti operano a relativamente bassa capacità di traspor to ma con alta affinità, ossia sono in grado di effettuare il traspor to in presenza di basse concentrazioni di ioni Ca 2+ ; queste sono presenti in tutte le cellule dell organismo. In molte cellule, le ATPasi sono affiancate da scambiator i Na + -Ca 2+,che sono dotati di elevata capacità di traspor to ma hanno una relativamente ridotta affinità per gli ioni calcio: ciò vuol dire che iniziano ad operare quando le concentrazioni calciche intracellular i sono relativamente elevate (circa un ordine di grandezza sopra leconcentrazioni basali). Come accennato, si -4-

5 suppone che le ATPasi operino una regolazione fine degli ioni Ca 2+,eche gli scambiatori Na + -Ca 2+ entr ino in gioco per compensare massicci ingressi di ioni -Ca 2+ nella cellula. Va poi osservato che esistono in molti organi e tessuti specifici traspor tatori calcici, e specifiche isofor me di questi, cui si può attribuire la funzione di adeguare la regolazione del Ca 2+ intracellulare alle esigenze dello specifico tessuto. Inogni caso, per la loro rilevanza funzionale i meccanismi di traspor to degli ioni Ca 2+ sono dotati di un elevato grado di ridondanza, risultando quindi particolar mente complessi Traspor to verso gli organelli Un altra ser ie di meccanismi opera traspor tando gli ioni Ca 2+ dal citoplasma agli organelli intracellular i, anche in questo caso con la doppia funzione di mantenere l integrità dei depositi calcici e di ripr istinare i bassi livelli citoplasmatici di questi ioni che si riscontrano in condizioni di riposo. Queste ATPasi sono estremamente abbondanti nel muscolo scheletrico, dove possono rappresentare una frazione rilevante della massa proteica totale: é infatti nota una famiglia di ATPasi Ca 2+ -dipendenti del sistema sarcoplasmatico, che comprende almeno cinque membri che differ iscono tra loro per distribuzione tissutale, tur nover ed affinità per il calcio. Anche i mitocondri rappresentano un importante deposito in questo caso a lungo termine di ioni Ca 2+. A causa dell elevata differenza di potenziale (circa -140 mv) esistente tra l inter no dei mitocondri ed il citoplasma, in questi organelli sono attivi meccanismi di traspor to specifici, che sfruttano questa differenza di potenziale per attivare un traspor to attivo indiretto che opera dal citoplasma verso l interno dei mitocondri, traspor tando uno ione Ca 2+ contro due elettroni. In più, l elevata differenza di potenziale riduce for temente la mobilità verso l esterno degli ioni Ca 2+ :probabilmente in ragione di ciò, e anche del fatto che il calcio nei mitocondr i é in buona parte immobilizzato, il traspor to dai mitocondri al citoplasma non avviene tramite canali, ma per mezzo di ulter iore meccanismo di traspor to attivo indiretto, consistente in uno scambiatore Na + -Ca 2+ elettr icamente neutro, che scambia uno ione Ca 2+ contro due ioni Na AMP ciclico Il primo scoperto ed ancor oggi il meglio conosciuto dei secondi messaggeri é l adenosina monofosfato ciclico (camp). La via che si serve di questo secondo messaggero é spesso utilizzata da recettori distinti, eccitator i ed inibitori, co-presenti nella stessa cellula, che mediano eventi cellulari con significato funzionale opposto tramite differenti proteine G (G s eg i.adesempio, negli adipociti l occupazione dei recettori β adrenergici per quattro diversi mediatori (adrenalina, ACTH, glucagone, TSH) aumenta la velocità di degradazione dei lipidi, mentre nelle stesse cellule l attivazione dei recettori α 2 adrenergici stimola una sequenza di eventi che si traduce in una diminuzione della velocità di degradazione dei lipidi. L camp è sintetizzato a partire dall ATP da un enzima di membrana, l adenil ciclasi (o adenilato ciclasi), i cui siti catalitici (si pensa due) sono affacciati alla faccia citoplasmatica della membrana. A seguito dell attivazione di quest enzima, le concentrazioni intracellular i di camp aumentano in pochi secondi di circa cinque volte a par tire dal valore di riposo (intorno a 1 x 10-7 M). Eccetto che nel caso dei neuroni olfattor i, descr itto oltre, tutti gli effetti del camp sono mediati da una specifica proteina chinasi, la chinasi A. Questa é una proteina tetramer ica, con due subunità catalitiche e due subunità regolatrici). Quando due molecole di camp si legano ad ognuna delle due subunità regolatrici della chinasi A (quattro in totale), l occupazione di questi siti compor ta un cambiamento confor mazionale tale da indebolire il legame tra lecomponenti regolatrici e quelle catalitiche; in conseguenza, le subunità regolatrici si staccano da quelle catalitiche; l ulteriore cambiamento confor mazionale indotto dalla sostituzione delle interazioni proteina - proteina con quelle proteina - solvente attiva poi i siti enzimaticamente attivi delle subunità catalitiche. La successione degli eventi relativi al camp può essere schematizzata come segue: 1. l occupazione dei siti recettor iali dei recettori stimolator i o inibitor i compor ta un cambiamento confor mazionale della molecola del recettore che (2) conduce all attivazione della proteina G, come descritto altrove; 3. la subunità α della proteina G interagisce con l adenilciclasi; 4. l interazione con la proteina G modifica la confor mazione dell enzima, stabilizzandolo nella sua confor mazione attiva, in cui é in grado di catalizzare la conversione dell ATP citoplasmatico in camp. Questo diffonde nel citoplasma fino ad incontrare le chinasi A; 5. Le chinasi A si attivano come descritto sopra; 6. le subunità catalitiche fosfor ilano le specifiche proteine effettr ici; 7. il cambiamento confor mazionale indotto dalla fosfor ilazione mette gli effettor i in grado di esprimere la funzione -5-

6 cellulare che deve essere modificata. Per ripor tare un esempio particolar mente semplice, l azione dell adrenalina sul muscolo scheletrico é mediata da recettori β adrenergici, l occupazione dei siti recettoriali dei quali induce aumento delle concentrazioni di camp e attivazione della chinasi A. Per ripor tare un esempio particolar mente semplice e ben noto, nel tessuto muscolare la chinasi A ha come substrato due distinti enzimi: il primo é la fosfor ilasi chinasi che, quando attivata dalla fosfor ilazione da parte della chinasi A, fosfor ila a sua volta la glicogeno fosfor ilasi, enzima che catalizza il rilascio dal glicogeno di glucosio 1-fosfato, che può così essere avviato alla glicolisi; il secondo enzima fosfor ilato dalla chinasi A è la glicogeno sintetasi; l attività di questo enzima é inibita nello stato confor mazionale che viene assunto a seguito della fosfor ilazione. Il risultato di queste interazioni é un aumento della quantità di glucosio 1-fosfato disponibile per la contrazione muscolare. Molto spesso l aumento delle concentrazioni di camp induce un aumento delle concentrazione citoplasmatiche di ioni Ca 2+,che possono provenire sia da depositi intracellular i che da sede extracellulare. L interazione del camp con gli ioni Ca 2+ avviene con var ie modalità: nel caso più semplice, ilcamp stimola direttamente l apertura dicanali calcici a controllo di nucleotidi ciclici; in questo caso il camp agisce come un ligando intracellulare per i canali calcici il cui sito recettoriale é intracellulare. Alter nativamente, la chinasi A può fosfor ilare traspor tatori calcici, modificandone l attività, oppure fosfor ilare canali calcici con un effetto regolator io positivo sull aper tura. In tutti questi i casi, il risultato é un aumento citoplasmatico di ioni Ca 2+,che si legano alle specifiche proteine calcio-leganti: saranno poi queste ultime ad indurre la funzione cellulare finale, come avviene ad esempio nel caso delle catene leggere della clatrina che partecipano alla formazione di vescicole di secrezione. Inoltre, gli enzimi che degradano il camp possono essere regolati dalla calmodulina. Ancora, enzimi regolati indipendentemente dagli ioni Ca 2+ edal camp possono interagire fra loro, oavere effetti interagenti (in senso positivoonegativo) sulle stesse molecole bersaglio. Esistono poi specifici meccanismi di disattivazione: infatti, avviene piuttosto raramente nella cellula che un effetto metabolico venga lasciato decadere spontaneamente. Nel caso specifico, tre principali sequenze di eventi mettono termine alla trasmissione del segnale intracellulare: 1. L adenilciclasi, in assenza di attivazione da parte della proteina G, riassume la sua confor mazione non attiva, decadendo nel suo stato non catalitico. Questa inattivazione é funzione della dinamica degli scambi energetici tra la molecola di enzima e l ambiente esterno e, come tale, é molto veloce; 2. Le concentrazioni intracellular i di camp vengono ripor tate ai livelli basali da due processi fondamentali: idrolisi del camp a AMP ad opera delle fosfodiesterasi presenti nella cellula e traspor to del camp in direzione extracellulare. La diminuzione delle concentrazioni di secondo messaggero così indotta determina la disattivazione dell effettore attivato dal camp. 3.Infine, gli effettor i che erano stati fosfor ilati dalle chinasi A devono essere defosfor ilati ad opera di una delle proteina fosfatasi citoplasmatiche (essenzialmente le proteina fosfatasi I, IIA e IIB). 5. Fosfoinositidi Come accennato, la via dei fosfoinositidi condivide con quella del camp i primi due eventi: 1. l occupazione del sito recettoriale determina l attivazione della proteina G nota come G q ;2.questa attiva unenzima, in questo caso una fosfodiesterasi, e precisamente la fosfoinositidasi C o fosfolipasi C-β. Questa é specifica per il fosfatidilinositolo (4,5)-difosfato (PIP 2 ), che viene scisso in inositolo (1,4,5)-trifosfato (IP 3 )ediacilglicerolo. Inoltre, sembra molto probabile il ruolo di un altro prodotto del metabolismo del PIP 2,precisamente l inositolo (1,3,4,5)-tetrafosfato (IP 4 ) che deriva dalla fosfor ilazione dell IP 3. Al livello immediatamente successivo, questa via si biforca: il diacilglicerolo pochissimo polare rimane nella membrana, diffonde nello strato lipidico e va ad attivare un enzima di membrana (in genere la chinasi C o PCK) che, essendo polare, può diffondere nel citoplasma e fosfor ila proteine bersaglio che sono diverse in diverse cellule. Ad esempio, ripor tando in modo estremamente sintetico solo una delle molte vie note, la chinasi C può fosfor ilare (attivandola) una seconda chinasi (MAP-chinasi), che a sua volta fosfor ila ed attiva una proteina regolatrice dei geni. L IP 3 (e probabilmente anche l IP 4 ), polari, diffondono invece nel citoplasma e stimolano il rilascio di ioni Ca 2+ dai depositi intracellular i. Almeno una parte del rilascio di ioni Ca 2+ dagli organelli nei quali questi sono sequestrati avviene tramite canali calcici a controllo di ligando, irecettor i per l IP 3 localizzati nella membrana degli organelli di deposito per il calcio. Aloro volta gli ioni Ca 2+ agiscono come messaggeri finali per stimolare le proteine effettr ici, come descritto nel caso del camp. Malgrado che il ruolo dell IP 4 sia ancora poco chiaro, invar i tessuti é stato notato sinergismo di azione tra IP 3 eip 4.Ciò vuol dire che almeno in -6-

7 cer ti casi la presenza di entrambi i fosfoinositidi sembra necessar ia per la mobilitazione degli ioni Ca 2+.In effetti, mentre l IP 3 éingrado di agire in modo autonomo, l IP 4,almeno apparentemente, richiede la presenza di IP 3.Vainfine notato che sono stati identificati inositoli più altamente fosfor ilati (IP 5 eip 6 ), la cui funzione é ancora da chiar ire. In fine, la relativa impor tanza di questi meccanismi in differenti tessuti é verosimilmente diverso. Ad esempio, in cellule eccitabili la regolazione della concentrazione di ioni calcio avviene tramite i canali calcici a controllo di potenziale localizzati nella membrana plasmatica e i recettori per la rianodina localizzati nella membrana di organelli, come il reticolo sarcoplasmatico del tessuto muscolare. Questi meccanismi non operano nel caso delle cellule non eccitabili, nelle quali la gran par te del calcio dovrebbe essere mobilitata tramite i recettori per l IP 3 enei quali il ruolo dei fosfoinositidi dovrebbe quindi essere comparativamente più rilevante che nelle cellule eccitabili. Anche in questo caso, esistono ovviamente meccanismi per mettere termine alla risposta cellulare: in primo luogo l IP 3 édefosfor ilato piuttosto velocemente da una specifica fosfatasi; inoltre, gli ioni Ca 2+ che sono entrati in risposta all elevarsi delle concentrazioni di IP 3 sono ripor tati in direzione periplasmatica ad opera dei meccanismi descritti sopra. 6. GMP ciclico Da un punto di vista chimico, il cgmp é molto simile al camp, dal quale differ isce solo perchè l adenosina é sostituita da una guanina. L associazione con le funzioni recettoriali di membrana della guanilato ciclasi che catalizza la conversione del GTP in cgmp é apparentemente meno ubiquitaria di quella del camp, ed stata dimostrata solo in pochi casi. A quanto é noto, come il camp anche il cgmp, agisce attivando una o più proteina chinasi (specificamente, chinasi G). Vi sono anche consistenti indicazioni del fatto che la sintesi di cgmp sia connessa con l attivazione della via dei fosfoinositidi e con quella dell acido arachidonico. In più, anche in questo caso e anche se in modo non sempre chiaro,sono chiamati in causa gli ioni Ca 2+. I recettor i con attività guanilato ciclasica sono in grado di catalizzare la sintesi di cgmp a partire dal GTP. Questi recettori sono attivati dai peptidi (o fattor i) atr iali natr iuretr ici (ANP o ANF, una famiglia di peptidi che hanno proprietà vasodilatator ie efavor iscono l escrezione renale di Na + e, non ostante il nome, anche di K + ). Infatti, i recettori per questi peptidi, che sono rilasciati dal tessuto atriale in condizioni di elevata pressione ar teriosa, sono localizzati nel parenchima renale e nei distretti vascolar i per ifer ici. Si tratta di recettori ad attività enzimatica (appunto, guanilato ciclasica) caratter izzati dalla struttura classico di questo tipo di recettor i (una singola catena polipeptidica con singolo passaggio transmembrana) che, al contrar io di quanto avviene nel caso della massima parte dei recettori con attività enzimatica, non esercitano i loro effetti sul genoma, ma su effettor i cellular i. Infatti, il GMP ciclico la cui sintesi é catalizata dall occupazione dei siti recettor iali con le modalità consuete per questi recettori (dimer izzazione, fosfor ilazione reciproca e conseguente attivazione del sito catalitico), tramite una specifica chinasi (chinasi G), interagisce con i traspor tatori di membrana e con la muscolatura vascolare, rispettivamente aumentandone l attività e causandone il rilassamento, con effetto ipotensivo a livello dell intero organismo. Inoltre, il cgmp é coinvolto nel meccanismo della visione, come descritto al Acido arachidonico Vi sono numerose evidenze a proposito dell attivazione, dipendente dal GTP, di un altra via di comunicazione, quella relativa all acido arachidonico e di suoi metaboliti; questi composti sono attivi tanto come secondi messaggeri insede intracellulare quanto come trasmettitor i in sede extracellulare. L acido arachidonico costituisce infatti il principale precursore dal quale sono sintetizzati gli eicosanoidi (prostglandine, prostacicline, tromboxani e leucotrieni). Var ie vie metaboliche sono in grado di produrre acido arachidonico e suoi derivati, spesso ma apparentemente non sempre sotto controllo di specifici recettor i etramite proteine G. Quelli che seguono sono alcuni esempi noti: i. Var ie fosfolipasi A 2 sono in grado di liberare acido arachidonico partendo dalla gran par te dei fosfolipidi di membrana (sopratutto fosfatidilser ina, fosfatidilcolina e fosfatidiletanolammina). Infatti, il digliceride dall esterificazione del quale der ivano i fosfolipidi che fanno parte delle membrane é molto spesso proprio l acido arachidonico, che può venire quindi liberato a partire da questi composti. ii. Anche le fosfolipasi C possono liberare acido -7-

8 arachidonico partendo sia da fosfoinositidi che dalla fosfatidilcolina. Questa reazione genera un arachidonglicer ide dal quale l acido arachidonico può essere liberato per azione di una lipasi. iii. Ancora, diglicer idi possono essere fosfor ilati da una specifica chinasi for nendo acido fosfatidico. L acido arachidonico viene quindi prodotto da una fosfolipasi A 2,come visto al punto i. Come avviene nel caso di tutte le vie di secondi messaggeri conosciuti, sono note var ie possibilità di interazione tra l acido arachidonico e suoi metaboliti e altri secondi messaggeri. Ad esempio, la lisofosfatidilcolina stimola la camp fosfodiesterasi; l acido arachidonico stimola la fosfoinositasi C, con conseguente produzione di IP 3 ed IP 4 ;var i metaboliti dell acido arachidonico aumentano i livelli citosolici di Ca 2+,indipendentemente dal loro effetto sulla produzione di inositolofosfati. 8. Gas: ossido nitrico e ossido di carbonio Entrambe queste molecole mediano sia segnali extracellular i locali (l emivita di entrambe le molecole é molto breve, nell ordine dei secondi) come anche segnali intratracellular i. Infatti, questi gas hanno una sorte assai diversa da quella di altri mediator i: una volta sintetizzati, diffondono liberamente sia dentro che fuori delle cellule di origine, fino ad incontrare i loro "recettori" che, nel caso dell ossido di azoto, sono costituiti dall atomo di ferro del sito attivo della guanilato ciclasi, stimolando così la sintesi di questo secondo messaggero. Data la sua capacità di diffondere rapidamente e la sua prevalente localizzazione nel sistema ner voso centrale, l ossido di azoto agisce sopratutto come neurotrasmettitore. Anche a causa delle loro peculiar i caratter istiche (dato che diffondono molto rapidamente, sono di difficile localizzazione), a tutt oggi la massima parte delle infor mazioni su queste sostanze si hanno relativamente alla localizzazione anatomica dell enzima che catalizza la sintesi dell ossido nitrico, la NO sintetasi, mentre ancora più scarse sono le infor mazioni relative all ossido di carbonio. Nel sistema nervoso centrale e nei vasi la NO sintetasi richiede la presenza di calmodulina: per conseguenza, la sintesi di NO é stimolata dagli agonisti glutamminergici (ad esempio, irecettor i per l NMDH), che permettono l ingresso di ioni Ca 2+.Questi occupano i siti di legame della calmodulina che in queste condizioni può agire come cofattore per l NO sintetasi. 9. Trasduzione dei segnali olfattori e visivi La trasduzione dei segnali olfattor i, come di quelli visivi, dipendono dall esistenza di canali ionici regolati da nucleotidi ciclici che agiscono non come secondi messaggeri, ma come ligandi intracellular i per canali a controllo di ligando il cui sito recettoriale é intracellulare. In entrambi i casi l occupazione di recettori metabotròpi ha quindi un effetto finale ionotròpo. L esistenza di canali analoghi é stata anche descritta nel tessuto cardiaco e renale, dove laloro funzione é tutt oraignota Olfatto Le cellule sensibili agli odori sono costituite da neuroni perifer ici bipolar i localizzati nella mucosa olfattiva, un area specializzata della mucosa nasale. Questi neuroni sono dotati di fascetti di cilia modificate che si estendono verso la cavità nasale, nella membrana delle quali sono localizzati specifici recettori accoppiati alle proteine G. Le sostanze traspor tate dall ar ia inspirata vanno in soluzione nel muco che ricopre l epitelio, legandosi ad una proteina vettr ice che le traspor ta fino alla membrana delle cilia. Le sostanze in grado di legarsi ad uno (o più d uno) dei var i recettor i interessati ne vanno ad occupare il sito recettoriale, come qualsiasi altro ligando. Con le modalità consuete per questa classe di recettori, l occupazione del sito recettor iale stabilizza un cambiamento confor mazionale che induce la dissociazione di una specifica proteina G, nota come G olf.lacui subunità α della G olf si lega all adenilcoclasi, attivandola ed inducendo un aumento delle concentrazioni citoplasmatiche di camp. Questo interagisce direttamente con canali cationici campdipendenti (vedi "Note sui recettori", 2.4) il cui sito recettoriale é intracellulare ed il cui canale fisico, come di norma, é chiuso in assenza di ligando. Il legame con l camp stabilizza la confor mazione in cui il canale fisico é aperto, aumentando la conduttanza di membrana essenzialmente per gli ioni Na +.Ciò induce una depolar izzazione che si propaga in direzione centripeta verso il bulbo olfattivo che costituisce la prima stazione delle vie olfattive. Come di norma, il segnale si mantiene fino a quando le concentrazioni -8-

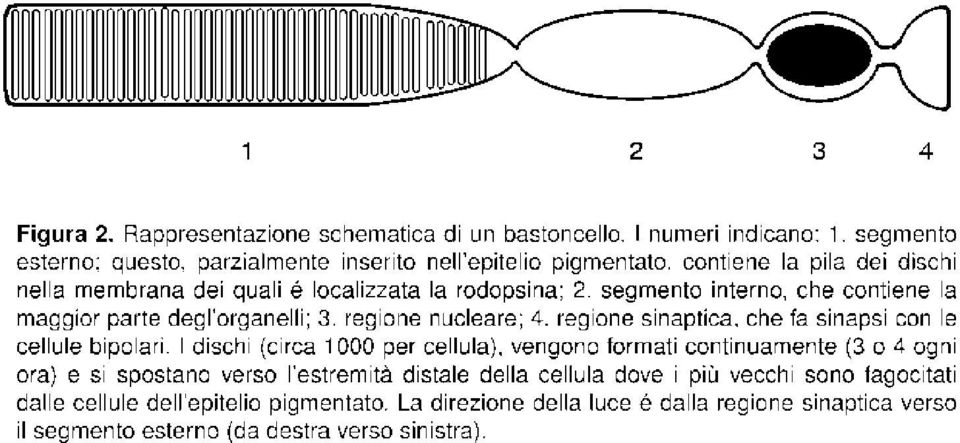
9 extracellular i di ligando rimangono sufficientemente elevate, mentre in sede intracellulare le concentrazioni di camp sono ripor tate ai valor i basali da specifiche fosfodiesterasi. Va anche osservato che esistono altri recettor i olfattivi, meno ben conosciuti, che agiscono tramite la via dell inositolo trifosfato; in questo caso, il segnale in uscita é generato dall apertura dicanali calcici IP 3 -dipendenti. Ancora, non ostante che gli odori fondamentali descritti siano solo sei, si suppone che siano interessati var ie centinaia di recettori distinti; ciò dovrebbe comportare che, per rispondere al grandissimo numero di sostanze riconoscibili, la specificità di questi recettori sia molto bassa, mentre il riconoscimento dello specifico odore con ogni probabilità avviene integrando in sede centrale i segnali provenienti dalle var ie cellule in grado di rispondere alla stessa molecola Visione Canali ionici con sito recettoriale intracellulare regolati da un nucleotide ciclico (cgmp) e controllati da recettor i accoppiati alle proteine G sono coinvolti anche nel meccanismo della visione. Nelle cellule fotorecettr ici (anche in questo caso neuroni perifer ici) il segnale elettrico efferente consiste in un iper polarizzazione, non una depolarizzazione, delle cellule recettrici, ed il meccanismo é quindi parzialmente diverso da quello descritto sopra. In condizioni basali (cioé di assenza di luce), le cellule fotorecettr ici sono iperpolar izzate. Infatti, nella membrana plasmatica del segmento esterno (Figura 2), esistono canali cationici a controllo di ligando relativamente aspecifici (brevemente descritti in "Note sui recettor i", 2.4) il cui sito recettoriale (intracellulare) lega il cgmp. Dato il rappor to tra le concentrazioni basali di cgmp e la costante di dissociazione per il cgmp dei siti recettoriali dei canali, questi sono in gran par te occupati ed il canale fisico é aperto. Quindi, in assenza di luce i canali lasciano passare una corrente (prevalentemente sodica) detta corrente di buio, le cellule sono depolarizzate (a circa -40 mv) ed esiste un rilascio continuo di mediatore sinaptico verso le cellule bipolari. La trasduzione di un segnale luminoso ha inizio con l arrivo di un fotone al complesso recettoriale, costituito da un recettore accoppiato alle proteine G, la rodopsina, e da una molecola, il retinale, legata covalentemente al sito recettoriale della rodopsina. In condizioni basali il retinale si trova in configurazione cis. L energia relativa all arr ivo di un fotone é in grado di indurre l isomerizzazione del retinale (dalle for ma cis a quella trans) evento che, a sua volta, induce nella rodopsina il cambiamento confor mazionale adatto a pilotare la dissociazione della specifica proteina G (trasducina o G t ). La subunità α della trasducina può quindi interagire con una cgmp-fosfodiesterasi presente nei dischi. Quest enzima é costituito da tre subunità la più piccola delle quali (la γ) é regolatr ice. L interazione della subunità α della trasducina con la cgmp-fosfodiesterasi ne determina un cambiamento confor mazionale a seguito del quale la subunità regolatrice si stacca, attivando la funzione enzimatica. Dato che questa consiste nel catalizzare l idrolisi del cgmp, la concentrazione intracellulare di questa molecola si abbassa bruscamente. Al diminuire delle concentrazioni di ligando, le probabilità che i siti recettoriali dei canali ionici possano essere occupati diminuiscono e i canali tendono quindi a trovarsi prevalentemente nella confor mazione corr ispondente al canale fisico chiuso, facendo cessare la corrente di buio. Sopratutto per l esistenza di canali potassici cronicamente aperti, la membrana si iperpolar izza, la liberazione di mediatore chimico verso le cellule bipolari si riduce e questo segnale, oppor tunamente integrato, viene trasmesso in sede centrale. Lo stato di apertura dei canali ha effetto regolativo anche sulla sintesi del cgmp: questo ( e infatti sintetizzato da una guanilato ciclasi Ca 2+ -dipendente. Data la poca specificità dei canali cationici del segmento esterno, quando questi sono aperti ossia in condizioni di buio permettono l ingresso anche di ioni Ca 2+.Questi si legano ad una proteina calcio-legante (la recoverina) che, atipicamente, éinattiva quando isuoi siti calcio-leganti sono occupati. La recoverina é invece attiva abassi livelli calcici (ossia in condizioni di segnale luminoso), condizioni sotto le quali si può legare alla guanilato ciclasi, stimolando la sintesi di cgmp. Questo meccanismo permette alla cellula recettrice sia di rigenerare rapidamente la corrente di buio dopo uno stimolo breve che di adattarsi alla luce, diminuendo la risposta in condizioni di illuminazione continua. -9-

10
11
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
I RECETTORI ACCOPPIATI A PROTEINE G
 I RECETTORI ACCOPPIATI A PROTEINE G I recettori accoppiati a proteine G sono costituiti da un unica subunità formata da un filamento di aminoacidi che attraversa 7 volte la membrana plasmatica Il sito
I RECETTORI ACCOPPIATI A PROTEINE G I recettori accoppiati a proteine G sono costituiti da un unica subunità formata da un filamento di aminoacidi che attraversa 7 volte la membrana plasmatica Il sito
Segnalazione cellulare e trasduzione del segnale. Comunicazione fra le cellule
 Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce
Segnalazione cellulare e trasduzione del segnale Comunicazione fra le cellule Le cellule comunicano e interagiscono tra loro tramite il fenomeno della segnalazione cellulare Una cellula segnalatrice produce
in Terminazione Neurone Dendriti Soma Fessura sinaptica sinaptica Nucleo dendrite Segmento iniziale Sinapsi inibitoria Segmento mielinico Assone
 Le funzioni del sistema nervoso si basano sull attività dei neuroni che consiste nel generare, trasmettere ed elaborare informazioni nervose, che dipendono da modificazioni del potenziale di membrana,
Le funzioni del sistema nervoso si basano sull attività dei neuroni che consiste nel generare, trasmettere ed elaborare informazioni nervose, che dipendono da modificazioni del potenziale di membrana,
Farmaci del sistema nervoso autonomo
 Farmaci del sistema nervoso autonomo Farmaci colinergici Esercitano i loro effetti farmacologici sul sistema nervoso parasimpatico che utilizza Acetilcolina come mediatore chimico L acetilcolina (Ach)
Farmaci del sistema nervoso autonomo Farmaci colinergici Esercitano i loro effetti farmacologici sul sistema nervoso parasimpatico che utilizza Acetilcolina come mediatore chimico L acetilcolina (Ach)
scaricato da www.sunhope.it
 Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Recettori a tirosina chinasi I recettori a tirosina chinasi presentano vari domini Una regione di legame (extracellulare) Una regione transmembrana Una coda intracellulare con numerose tirosine scaricato
Comunicazione chimica
 Comunicazione chimica La comunicazione tra le cellule Modalità di comunicazione cellulare: Mediante messaggi elettrici Mediante messaggi chimici In ambedue le modalità il messaggio, sia esso una variazione
Comunicazione chimica La comunicazione tra le cellule Modalità di comunicazione cellulare: Mediante messaggi elettrici Mediante messaggi chimici In ambedue le modalità il messaggio, sia esso una variazione
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido
 Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
Il mantenimento dell omeostasi cellulare dipende dai sistemi che permettono lo scambio di molecole tra citoplasma e liquido extracellulare e dalla loro regolazione. Membrana cellulare Ogni cellula presenta
 TRASDUZIONE DEL SEGNALE LE PRINCIPALI MOLECOLE CHE PROVVEDONO ALLO SCAMBIO DI INFORMAZIONI TRA LE CELLULE SONO FATTORI SOLUBILI QUALI NEUROTRASMETTITORI, ORMONI, FATTORI DI CRESCITA, CITOCHINE. UN CASO
TRASDUZIONE DEL SEGNALE LE PRINCIPALI MOLECOLE CHE PROVVEDONO ALLO SCAMBIO DI INFORMAZIONI TRA LE CELLULE SONO FATTORI SOLUBILI QUALI NEUROTRASMETTITORI, ORMONI, FATTORI DI CRESCITA, CITOCHINE. UN CASO
Proteine integrali di membrana legate sul versante esterno a gruppi di carboidrati. Formati da diverse subunità che circoscrivono un poro acquoso che
 Canali ionici Proteine integrali di membrana legate sul versante esterno a gruppi di carboidrati. Formati da diverse subunità che circoscrivono un poro acquoso che permette il passaggio selettivo di ioni.
Canali ionici Proteine integrali di membrana legate sul versante esterno a gruppi di carboidrati. Formati da diverse subunità che circoscrivono un poro acquoso che permette il passaggio selettivo di ioni.
Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati
 Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati della membrana. Il potenziale di membrana (negativo
Potenziale di membrana: Differenza di potenziale elettrico a cavallo della membrana cellulare dovuta ad una diversa distribuzione ionica ai due lati della membrana. Il potenziale di membrana (negativo
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA
 CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
CONTROLLO ORMONALE DEL METABOLISMO GLUCIDICO DA PARTE DI GLUCAGONE, ADRENALINA E INSULINA QUESTI ORMONI REGOLANO IL FLUSSO DEI METABOLITI NELLA GLICOLISI, NELLA GLICOGENO-SINTESI, NELLA GLIGENO-LISI E
La membrana cellulare racchiude il protoplasma, la sostanza vivente che costituisce la cellula, e lo separa dall ambiente esterno, extracellulare.
 Membrana Plasmatica La membrana cellulare racchiude il protoplasma, la sostanza vivente che costituisce la cellula, e lo separa dall ambiente esterno, extracellulare. La sua funzione principale è quella
Membrana Plasmatica La membrana cellulare racchiude il protoplasma, la sostanza vivente che costituisce la cellula, e lo separa dall ambiente esterno, extracellulare. La sua funzione principale è quella
Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
, si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.") Membrana cellulare Proteine inserite nella membrana cellulare: Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
Membrana cellulare Proteine inserite nella membrana cellulare: Integrali (transmembrana), si estendono attraverso l intera membrana. Classificate in famiglie a seconda del numero di segmenti transmembrana.
 forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
forme diabetiche Diabete di tipo I o magro o giovanile (IDDM): carenza primaria di insulina, più frequente nei giovani e nei bambini. Il deficit insulinico consegue (per predisposizione ereditaria o per
04/04/14. Fondamenti di biochimica Terza edizione. Le ghiandole principali del sistema endocrino. Capitolo 13 La segnalazione biochimica
 Fondamenti di biochimica Terza edizione Donald Voet Judith G. Voet Charlotte W. Pratt La segnalazione biochimica Copyright 2013 Zanichelli editore S.p.A. Gli ormoni Conce& chiave 13.1 Gli ormoni endocrini
Fondamenti di biochimica Terza edizione Donald Voet Judith G. Voet Charlotte W. Pratt La segnalazione biochimica Copyright 2013 Zanichelli editore S.p.A. Gli ormoni Conce& chiave 13.1 Gli ormoni endocrini
MACROSTRUTTURA DEL MUSCOLO STRIATO SCHELETRICO
 FUNZIONE DEL MUSCOLO SCHELETRICO Ultrastruttura della cellula muscolare. Miofilamenti. Meccanismo della contrazione. Trasmissione neuromuscolare. Scossa semplice e tetano muscolare. Unità motoria. PROPRIETA
FUNZIONE DEL MUSCOLO SCHELETRICO Ultrastruttura della cellula muscolare. Miofilamenti. Meccanismo della contrazione. Trasmissione neuromuscolare. Scossa semplice e tetano muscolare. Unità motoria. PROPRIETA
TRASDUZIONE DEL SEGNALE
 TRASDUZIONE DEL SEGNALE TRASDUZIONE DEL SEGNALE Specificità (specificità riconoscimento) Amplificazione e diversificazione della risposta (cascata enzimatica) Integrazione tra segnali Spegnimento del segnale
TRASDUZIONE DEL SEGNALE TRASDUZIONE DEL SEGNALE Specificità (specificità riconoscimento) Amplificazione e diversificazione della risposta (cascata enzimatica) Integrazione tra segnali Spegnimento del segnale
2. Fisiologia Cellulare Diffusione, Trasporto, Osmosi
 2. Fisiologia Cellulare Diffusione, Trasporto, Osmosi Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Diffusione semplice e mediata da
2. Fisiologia Cellulare Diffusione, Trasporto, Osmosi Prof. Carlo Capelli Fisiologia Laurea in Scienze delle attività motorie e sportive Università di Verona Obiettivi Diffusione semplice e mediata da
Comunicazione intercellulare. Negli organismi pluricellulari le cellule comunicano tramite un ampia varietà di segnali extracellulari
 Comunicazione intercellulare Negli organismi pluricellulari le cellule comunicano tramite un ampia varietà di segnali extracellulari PRINCIPI GENERALI DELLA COMUNICAZIONE TRA CELLULE TRAMITE MOLECOLE SEGNALE
Comunicazione intercellulare Negli organismi pluricellulari le cellule comunicano tramite un ampia varietà di segnali extracellulari PRINCIPI GENERALI DELLA COMUNICAZIONE TRA CELLULE TRAMITE MOLECOLE SEGNALE
Meccanismi molecolari di trasduzione del segnale. Le vie di trasduzione del segnale sono molto specifiche ed estremamente sensibili.
 Meccanismi molecolari di trasduzione del segnale. Le vie di trasduzione del segnale sono molto specifiche ed estremamente sensibili. La sensibilita delle vie di trasduzione dipende da 3 fattori: -l affinita
Meccanismi molecolari di trasduzione del segnale. Le vie di trasduzione del segnale sono molto specifiche ed estremamente sensibili. La sensibilita delle vie di trasduzione dipende da 3 fattori: -l affinita
La pompa Na + /Glucosio: simporto
 MFN0366-A1 (I. Perroteau) - trasportatori e canali La pompa Na + /Glucosio: simporto Il trasportatore oscilla fra due stati alternativi (A e B); nello stato A la proteina è aperta nello spazio extracellulare,
MFN0366-A1 (I. Perroteau) - trasportatori e canali La pompa Na + /Glucosio: simporto Il trasportatore oscilla fra due stati alternativi (A e B); nello stato A la proteina è aperta nello spazio extracellulare,
Regolazione del metabolismo del glucosio
 Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
Regolazione del metabolismo del glucosio Regolazione coordinata di glicolisi e gluconeogenesi Glicolisi e gluconeogenesi sono regolate in modo reciproco La regolazione è a livello dei punti di deviazione
www.fisiokinesiterapia.biz
 www.fisiokinesiterapia.biz FARMACODINAMICA La farmacodinamica può essere definita come lo studio degli effetti biochimici e fisiologici dei farmaci e dei loro meccanismi d azione. Gli obiettivi dell analisi
www.fisiokinesiterapia.biz FARMACODINAMICA La farmacodinamica può essere definita come lo studio degli effetti biochimici e fisiologici dei farmaci e dei loro meccanismi d azione. Gli obiettivi dell analisi
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Helena Curtis N. Sue Barnes LA RESPIRAZIONE CELLULARE INDICE Demolizione del glucosio Glicolisi Fermentazione lattica Fermentazione alcolica Respirazione cellulare Ciclo di Krebs Catena di trasporto degli
Energia nelle reazioni chimiche. Lezioni d'autore di Giorgio Benedetti
 Energia nelle reazioni chimiche Lezioni d'autore di Giorgio Benedetti VIDEO Introduzione (I) L energia chimica è dovuta al particolare arrangiamento degli atomi nei composti chimici e le varie forme di
Energia nelle reazioni chimiche Lezioni d'autore di Giorgio Benedetti VIDEO Introduzione (I) L energia chimica è dovuta al particolare arrangiamento degli atomi nei composti chimici e le varie forme di
Biosintesi non ribosomiale di metaboliti peptidici bioattivi
 Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Biosintesi non ribosomiale di metaboliti peptidici bioattivi Principali bersagli degli antibiotici Gli antibiotici derivano per la maggior parte da composti naturali Strutture di alcuni peptidi bioattivi
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2015-16
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2015-16 BIOFISICA DELLE MEMBRANE Negli organismi pluricellulari, a parte i tessuti tegumentari e gli epiteli assorbenti
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I A.A. 2015-16 BIOFISICA DELLE MEMBRANE Negli organismi pluricellulari, a parte i tessuti tegumentari e gli epiteli assorbenti
Tipi di neurotrasmettitori
 Tipi di neurotrasmettitori Neurotrasmettitori classici: molecole a basso peso molecolare, di varia natura: acetilcolina amine biogene (dopamina, adrenalina, noradrenalina) istamina aminoacidi (GABA, glicina,
Tipi di neurotrasmettitori Neurotrasmettitori classici: molecole a basso peso molecolare, di varia natura: acetilcolina amine biogene (dopamina, adrenalina, noradrenalina) istamina aminoacidi (GABA, glicina,
La trasduzione del segnale intracellulare
 La trasduzione del segnale intracellulare Chemical signaling mechanisms Amplification in signal transduction pathways Un aspetto importante è il controllo temporale del comportamento cellulare: il trasferimento
La trasduzione del segnale intracellulare Chemical signaling mechanisms Amplification in signal transduction pathways Un aspetto importante è il controllo temporale del comportamento cellulare: il trasferimento
MECCANISMI DI COMUNICAZIONE CELLULARE
 MECCANISMI DI COMUNICAZIONE CELLULARE La COMUNICAZIONE CELLULARE è cruciale negli organismi multicellulari - serve a coordinare/organizzare le attività lo sviluppo dei vari organi/tessuti -Lo scambio di
MECCANISMI DI COMUNICAZIONE CELLULARE La COMUNICAZIONE CELLULARE è cruciale negli organismi multicellulari - serve a coordinare/organizzare le attività lo sviluppo dei vari organi/tessuti -Lo scambio di
Classificazione dei Sensori. (raccolta di lucidi)
") Classificazione dei Sensori (raccolta di lucidi) 1 Le grandezze fisiche da rilevare nei processi industriali possono essere di varia natura; generalmente queste quantità sono difficili da trasmettere e
Classificazione dei Sensori (raccolta di lucidi) 1 Le grandezze fisiche da rilevare nei processi industriali possono essere di varia natura; generalmente queste quantità sono difficili da trasmettere e
Il metabolismo dell RNA. Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
 Il metabolismo dell RNA I vari tipi di RNA Il filamento di DNA che dirige la sintesi dello mrna è chiamato filamento stampo o filamento antisenso. L altro filamento che ha sequenza identica a quella dello
Il metabolismo dell RNA I vari tipi di RNA Il filamento di DNA che dirige la sintesi dello mrna è chiamato filamento stampo o filamento antisenso. L altro filamento che ha sequenza identica a quella dello
Farmacodinamica II. Mariapia Vairetti. Dipartimento di Medicina Interna e Terapia Sezione di Farmacologia e Tossicologia Cellulare e Molecolare
 Farmacodinamica II Mariapia Vairetti Dipartimento di Medicina Interna e Terapia Sezione di Farmacologia e Tossicologia Cellulare e Molecolare Processo di riconoscimento fra farmaco e recettore TIPI DI
Farmacodinamica II Mariapia Vairetti Dipartimento di Medicina Interna e Terapia Sezione di Farmacologia e Tossicologia Cellulare e Molecolare Processo di riconoscimento fra farmaco e recettore TIPI DI
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) Metabolismo del glucosio La gluconeogenesi epatica è regolata
Recettori di superficie
 Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
unità C2. Le trasformazioni energetiche nelle cellule
 unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
unità 2. Le trasformazioni energetiche nelle cellule Il trasporto nelle cellule avviene senza consumo di energia con consumo di energia trasporto passivo trasporto attivo attraverso il doppio strato fosfolipidico
Cellula nervosa tipo. Polarizzazione dinamica. Specificità delle connessioni. Fessura sinaptica Terminazione sinaptica. dendri te Segmento iniziale
 Le funzioni del sistema nervoso, anche le più complesse, si fondano sulle proprietà funzionali dei neuroni. L attività specifica dei neuroni consiste nel generare, trasmettere ed elaborare informazioni.
Le funzioni del sistema nervoso, anche le più complesse, si fondano sulle proprietà funzionali dei neuroni. L attività specifica dei neuroni consiste nel generare, trasmettere ed elaborare informazioni.
Prof. Maria Nicola GADALETA DISPENSA N. 19
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Tecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Tecnologie Biochimiche DISPENSA
Giunzione neuromuscolare
 Il calcio provoca un rimaneggiamento di membrana e la sua conseguente fusione. Questo compito è svolto dalle proteine SNARE che si agganciano tra la membrana interna pre-sinaprica e quella della vescicola.
Il calcio provoca un rimaneggiamento di membrana e la sua conseguente fusione. Questo compito è svolto dalle proteine SNARE che si agganciano tra la membrana interna pre-sinaprica e quella della vescicola.
SISTEMI ENERGETICI. L ATP privato di uno dei suoi 3 radicali fosforici diventa ADP (adenosindifosfato).
.") SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
SISTEMI ENERGETICI LE FONTI ENERGETICHE MUSCOLARI I movimenti sono resi possibili, dal punto di vista energetico, grazie alla trasformazione, da parte dei muscoli, dell energia chimica ( trasformazione
Bioingegneria Elettronica I
 Bioingegneria Elettronica I Cenni alla fisiologia delle cellule e dei sistemi biologici A. Bonfiglio La cellula struttura generale La cellula Struttura generale della cellula Composizione dei liquidi intracellulare
Bioingegneria Elettronica I Cenni alla fisiologia delle cellule e dei sistemi biologici A. Bonfiglio La cellula struttura generale La cellula Struttura generale della cellula Composizione dei liquidi intracellulare
Βeta ossidazione degli acidi grassi BETA OSSIDAZIONE DEGLI ACIDI GRASSI 13/12/2013 SEMINARIO. Acidi grassi (2)
") Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
Acidi grassi (1) Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI Sono le principali fonti di energia per alcuni tessuti (es. muscolo cardiaco). Si genera molto più energia dall ossidazione degli
LA CORRENTE ELETTRICA
 L CORRENTE ELETTRIC H P h Prima che si raggiunga l equilibrio c è un intervallo di tempo dove il livello del fluido non è uguale. Il verso del movimento del fluido va dal vaso a livello maggiore () verso
L CORRENTE ELETTRIC H P h Prima che si raggiunga l equilibrio c è un intervallo di tempo dove il livello del fluido non è uguale. Il verso del movimento del fluido va dal vaso a livello maggiore () verso
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery
 Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
Corso di Laurea in Farmacia Insegnamento di BIOCHIMICA Angela Chambery Lezione 26 La gluconeogenesi Concetti chiave: Il fegato e il rene possono sintetizzare glucosio da lattato, piruvato e amminoacidi.
Metalli in medicina. L utilizzo dei metalli in medicina ha radici ben antiche. Il ferro ed il
 Metalli in medicina L utilizzo dei metalli in medicina ha radici ben antiche. Il ferro ed il rame, per esempio, erano utilizzati nella Grecia antica. Già da secoli il Hg 2+ era utilizzato nel trattamento
Metalli in medicina L utilizzo dei metalli in medicina ha radici ben antiche. Il ferro ed il rame, per esempio, erano utilizzati nella Grecia antica. Già da secoli il Hg 2+ era utilizzato nel trattamento
Legami chimici. Covalente. Legami deboli
 Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
Legami chimici Covalente Legami deboli Legame fosfodiesterico Legami deboli Legami idrogeno Interazioni idrofobiche Attrazioni di Van der Waals Legami ionici Studio delle macromolecole Lipidi
BIOSEGNALAZIONE. La ricezione e la trasmissione delle informazioni extracellulari Parte II
 BIOSEGNALAZIONE La ricezione e la trasmissione delle informazioni extracellulari Parte II Tipi generali di trasduttori di segnali RECETTORI CON ATTIVITA TIROSIN CHINASICA (RTK) INS: insulin VEGF: vascular
BIOSEGNALAZIONE La ricezione e la trasmissione delle informazioni extracellulari Parte II Tipi generali di trasduttori di segnali RECETTORI CON ATTIVITA TIROSIN CHINASICA (RTK) INS: insulin VEGF: vascular
I SISTEMI NEUROTRASMETTITORI
 I SISTEMI NEUROTRASMETTITORI PERCHÉ UNA SOSTANZA POSSA DIRSI NEUROTRASMETTITORE (NT) 1.1 La molecola deve essere localizzata nel neurone presinaptico IMMUNOCITOCHIMICA 1.2 La molecola deve essere sintetizzata
I SISTEMI NEUROTRASMETTITORI PERCHÉ UNA SOSTANZA POSSA DIRSI NEUROTRASMETTITORE (NT) 1.1 La molecola deve essere localizzata nel neurone presinaptico IMMUNOCITOCHIMICA 1.2 La molecola deve essere sintetizzata
Dal DNA all RNA. La trascrizione nei procarioti e negli eucarioti
 Dal DNA all RNA La trascrizione nei procarioti e negli eucarioti DOGMA CENTRALE DELLA BIOLOGIA MOLECOLARE Gene Regione di DNA che porta l informazione (= che CODIFICA) per una catena polipeptidica o per
Dal DNA all RNA La trascrizione nei procarioti e negli eucarioti DOGMA CENTRALE DELLA BIOLOGIA MOLECOLARE Gene Regione di DNA che porta l informazione (= che CODIFICA) per una catena polipeptidica o per
Propagazione e trasmissione di segnali nervosi
 Propagazione e trasmissione di segnali nervosi La comunicazione tra le cellule Modalità di comunicazione cellulare: Mediante messaggi elettrici Mediante messaggi chimici In ambedue le modalità il messaggio,
Propagazione e trasmissione di segnali nervosi La comunicazione tra le cellule Modalità di comunicazione cellulare: Mediante messaggi elettrici Mediante messaggi chimici In ambedue le modalità il messaggio,
Amplificatori Audio di Potenza
 Amplificatori Audio di Potenza Un amplificatore, semplificando al massimo, può essere visto come un oggetto in grado di aumentare il livello di un segnale. Ha quindi, generalmente, due porte: un ingresso
Amplificatori Audio di Potenza Un amplificatore, semplificando al massimo, può essere visto come un oggetto in grado di aumentare il livello di un segnale. Ha quindi, generalmente, due porte: un ingresso
DNA - RNA. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi.
 DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
DNA - RNA Le unità fondamentali costituenti il DNA e l RNA sono i Nucleotidi. Nucleotide = Gruppo Fosforico + Zucchero Pentoso + Base Azotata. Esistono 4 basi azotate per il DNA e 4 per RNA Differenze
OBESITA. 11 Aprile 2013
 OBESITA 11 Aprile 2013 L obesità è una patologia cronica caratterizzata dall eccessivo accumulo di tessuto adiposo nell organismo ed è causata da fattori ereditari e non ereditari che determinano un introito
OBESITA 11 Aprile 2013 L obesità è una patologia cronica caratterizzata dall eccessivo accumulo di tessuto adiposo nell organismo ed è causata da fattori ereditari e non ereditari che determinano un introito
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE
 COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
COMUNICAZIONE CELLULARE TRASDUZIONE DEL SEGNALE recettori (proteine recettoriali) vie di segnalazione intracellulare (molecole che elaborano il segnale e lo distribuiscono ai bersagli appropriati) recettori
Il trasporto di membrana e le proprietà elettriche delle membrane. www.fisiokinesiterapia.biz
 Il trasporto di membrana e le proprietà elettriche delle membrane www.fisiokinesiterapia.biz Membrana cellulare barriera al passaggio della maggior parte delle molecole polari Permette alla cellula di
Il trasporto di membrana e le proprietà elettriche delle membrane www.fisiokinesiterapia.biz Membrana cellulare barriera al passaggio della maggior parte delle molecole polari Permette alla cellula di
Metabolismo degli aminoacidi
 Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Metabolismo degli aminoacidi Quando gli aminoacidi provenienti dalla digestione gastrica e intestinale, non vengono utilizzati per la costruzione di nuove proteine vengono demoliti Transaminazione Per
Prof. Maria Nicola GADALETA
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche DISPENSA
Le Biomolecole I parte. Lezioni d'autore di Giorgio Benedetti
 Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
Le Biomolecole I parte Lezioni d'autore di Giorgio Benedetti LE BIOMOLECOLE Le biomolecole, presenti in tutti gli esseri viventi, sono molecole composte principalmente da carbonio, idrogeno, azoto e ossigeno.
1) FIBRE ROSSE A CONTRAZIONE LENTA (Tipo I) 2) FIBRE BIANCHE INTERMEDIE (Tipo IIa) 3) FIBRE BIANCHE A CONTRAZIONE RAPIDA (Tipo IIb)
 FIBRE ROSSE A CONTRAZIONE LENTA (Tipo I) 2) FIBRE BIANCHE INTERMEDIE (Tipo IIa) 3) FIBRE BIANCHE A CONTRAZIONE RAPIDA (Tipo IIb)") LE FIBRE MUSCOLARI La fibra muscolare è considerata l' unità funzionale del muscolo scheletrico o, più semplicemente, una delle tante cellule che lo compongono. Ogni muscolo è infatti formato da un certo
LE FIBRE MUSCOLARI La fibra muscolare è considerata l' unità funzionale del muscolo scheletrico o, più semplicemente, una delle tante cellule che lo compongono. Ogni muscolo è infatti formato da un certo
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare
 Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
CORRENTE E TENSIONE ELETTRICA LA CORRENTE ELETTRICA
 CORRENTE E TENSIONE ELETTRICA La conoscenza delle grandezze elettriche fondamentali (corrente e tensione) è indispensabile per definire lo stato di un circuito elettrico. LA CORRENTE ELETTRICA DEFINIZIONE:
CORRENTE E TENSIONE ELETTRICA La conoscenza delle grandezze elettriche fondamentali (corrente e tensione) è indispensabile per definire lo stato di un circuito elettrico. LA CORRENTE ELETTRICA DEFINIZIONE:
Il metabolismo del glicogeno e la gluconeogenesi. Copyright 2013 Zanichelli editore S.p.A.
 Il metabolismo del glicogeno e la gluconeogenesi Copyright 2013 Zanichelli editore S.p.A. Capitolo 16 La demolizione del glicogeno Concetti chiave 16.1 Il glicogeno, la forma di immagazzinamento del glucosio,
Il metabolismo del glicogeno e la gluconeogenesi Copyright 2013 Zanichelli editore S.p.A. Capitolo 16 La demolizione del glicogeno Concetti chiave 16.1 Il glicogeno, la forma di immagazzinamento del glucosio,
Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica.
 Concanavalina A Emoglobina subunità Trioso fosfato isomerasi Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica. 1 La conformazione è
Concanavalina A Emoglobina subunità Trioso fosfato isomerasi Una proteina qualsiasi assume costantemente un unica conformazione ben definita, cui è legata la sua azione biologica. 1 La conformazione è
LA REAZIONE ALL'AMBIENTE: L'ATTIVITÀ MUSCOLARE TUTTI I MOVIMENTI NEL CORPO E DEL CORPO SONO LA CONSEGUENZA DI CONTRAZIONI E RILASCIAMENTI MUSCOLARI
 LA REAZIONE ALL'AMBIENTE: L'ATTIVITÀ MUSCOLARE TUTTI I MOVIMENTI NEL CORPO E DEL CORPO SONO LA CONSEGUENZA DI CONTRAZIONI E RILASCIAMENTI MUSCOLARI CLASSIFICAZIONI DEI MUSCOLI: STRIATO SCHELETRICO STRATO
LA REAZIONE ALL'AMBIENTE: L'ATTIVITÀ MUSCOLARE TUTTI I MOVIMENTI NEL CORPO E DEL CORPO SONO LA CONSEGUENZA DI CONTRAZIONI E RILASCIAMENTI MUSCOLARI CLASSIFICAZIONI DEI MUSCOLI: STRIATO SCHELETRICO STRATO
CELLULE EUCARIOTICHE
 CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
CELLULE EUCARIOTICHE Le cellule eucariotiche sono di maggiori dimensioni, rispetto a quelle procariotiche (almeno 10 volte più grandi) Oltre a: membrana plasmatica, citoplasma, DNA e ribosomi (comuni a
Appunti sulla Macchina di Turing. Macchina di Turing
 Macchina di Turing Una macchina di Turing è costituita dai seguenti elementi (vedi fig. 1): a) una unità di memoria, detta memoria esterna, consistente in un nastro illimitato in entrambi i sensi e suddiviso
Macchina di Turing Una macchina di Turing è costituita dai seguenti elementi (vedi fig. 1): a) una unità di memoria, detta memoria esterna, consistente in un nastro illimitato in entrambi i sensi e suddiviso
La candela accesa. Descrizione generale. Obiettivi. Sequenza didattica e metodo di lavoro. Esperimenti sulla crescita delle piante
 Esperimenti sulla crescita delle piante unità didattica 1 La candela accesa Durata 60 minuti Materiali per ciascun gruppo - 1 candela - 1 vaso di vetro - 1 cronometro - 1 cannuccia - fiammiferi - 1 pezzo
Esperimenti sulla crescita delle piante unità didattica 1 La candela accesa Durata 60 minuti Materiali per ciascun gruppo - 1 candela - 1 vaso di vetro - 1 cronometro - 1 cannuccia - fiammiferi - 1 pezzo
TEST BIOLOGIA 1 ANNO ABEI Da inviare a connesso@alice.it entro e non oltre il 6 novembre 2015
 1) I batteri sono organismi: a- bicellulari b- monocellulari c- pluricellulari 2) I virus: a- possono riprodursi solo nell acqua b- possono riprodursi solo sulla superficie di una cellula c- possono riprodursi
1) I batteri sono organismi: a- bicellulari b- monocellulari c- pluricellulari 2) I virus: a- possono riprodursi solo nell acqua b- possono riprodursi solo sulla superficie di una cellula c- possono riprodursi
INTERVENTO DI CLAUDIA RICCARDI PLASMAPROMETEO - Dipartimento di Fisica Università degli Studi di Milano - Bicocca
 INTERVENTO DI CLAUDIA RICCARDI PLASMAPROMETEO - Dipartimento di Fisica Università degli Studi di Milano - Bicocca La ricerca come strumento per lo sviluppo aziendale: sinergia tra università e industria
INTERVENTO DI CLAUDIA RICCARDI PLASMAPROMETEO - Dipartimento di Fisica Università degli Studi di Milano - Bicocca La ricerca come strumento per lo sviluppo aziendale: sinergia tra università e industria
Il glicogeno (riserva di glucosio) è immagazzinato nel fegato e nei muscoli (con finalità diverse )
 è immagazzinato nel fegato e nei muscoli (con finalità diverse )") Glicogeno Glicogeno...1 Glicogenolisi...3 Glicogenosintesi...7 Regolazione ormonale della glicogenolisi e delle glicogenosintesi...13 Il recettore del glucagone e quello dell adrenalina sono 2 GPCR (G-Protein
Glicogeno Glicogeno...1 Glicogenolisi...3 Glicogenosintesi...7 Regolazione ormonale della glicogenolisi e delle glicogenosintesi...13 Il recettore del glucagone e quello dell adrenalina sono 2 GPCR (G-Protein
I documenti di www.mistermanager.it. Gli ingredienti per l allenamento per la corsa LE RIPETUTE
 I documenti di www.mistermanager.it Gli ingredienti per l allenamento per la corsa LE RIPETUTE Le Ripetute sono una delle forme di allenamento che caratterizzano i corridori più evoluti, in quanto partono
I documenti di www.mistermanager.it Gli ingredienti per l allenamento per la corsa LE RIPETUTE Le Ripetute sono una delle forme di allenamento che caratterizzano i corridori più evoluti, in quanto partono
Siamo così arrivati all aritmetica modulare, ma anche a individuare alcuni aspetti di come funziona l aritmetica del calcolatore come vedremo.
 DALLE PESATE ALL ARITMETICA FINITA IN BASE 2 Si è trovato, partendo da un problema concreto, che con la base 2, utilizzando alcune potenze della base, operando con solo addizioni, posso ottenere tutti
DALLE PESATE ALL ARITMETICA FINITA IN BASE 2 Si è trovato, partendo da un problema concreto, che con la base 2, utilizzando alcune potenze della base, operando con solo addizioni, posso ottenere tutti
Descrivere la struttura e la funzione di un fuso neuromuscolare. Che cosa si intende per coattivazione alfa-gamma? Differenziare fra riflesso
 Descrivere la struttura e la funzione di un fuso neuromuscolare. Che cosa si intende per coattivazione alfa-gamma? Differenziare fra riflesso miotatico fasico e riflesso miotatico tonico. Inibizione dell
Descrivere la struttura e la funzione di un fuso neuromuscolare. Che cosa si intende per coattivazione alfa-gamma? Differenziare fra riflesso miotatico fasico e riflesso miotatico tonico. Inibizione dell
TRASDUZIONE DEL SEGNALE CELLULARE
 TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
Dispositivi di rete. Ripetitori. Hub
 Ripetitori Dispositivi di rete I ripetitori aumentano la distanza che può essere ragginta dai dispositivi Ethernet per trasmettere dati l'uno rispetto all'altro. Le distanze coperte dai cavi sono limitate
Ripetitori Dispositivi di rete I ripetitori aumentano la distanza che può essere ragginta dai dispositivi Ethernet per trasmettere dati l'uno rispetto all'altro. Le distanze coperte dai cavi sono limitate
V= R*I. LEGGE DI OHM Dopo aver illustrato le principali grandezze elettriche è necessario analizzare i legami che vi sono tra di loro.
 LEGGE DI OHM Dopo aver illustrato le principali grandezze elettriche è necessario analizzare i legami che vi sono tra di loro. PREMESSA: Anche intuitivamente dovrebbe a questo punto essere ormai chiaro
LEGGE DI OHM Dopo aver illustrato le principali grandezze elettriche è necessario analizzare i legami che vi sono tra di loro. PREMESSA: Anche intuitivamente dovrebbe a questo punto essere ormai chiaro
Meccanismi di trasporto attivo e passivo 1
 Meccanismi di trasporto attivo e passivo 1 Il trasporto di sostanze attraverso la membrana cellulare può avvenire con la partecipazione attiva della membrana: in questo caso si parla di trasporto attivo
Meccanismi di trasporto attivo e passivo 1 Il trasporto di sostanze attraverso la membrana cellulare può avvenire con la partecipazione attiva della membrana: in questo caso si parla di trasporto attivo
Il muscolo liscio vascolare
 Il muscolo liscio vascolare La cellula muscolare liscia Giorgio Gabella, professor of histology University College, London La cellula muscolare liscia Eccitabilita : potenziale d azione o potenziali graduati
Il muscolo liscio vascolare La cellula muscolare liscia Giorgio Gabella, professor of histology University College, London La cellula muscolare liscia Eccitabilita : potenziale d azione o potenziali graduati
Regole della mano destra.
 Regole della mano destra. Macchina in continua con una spira e collettore. Macchina in continua con due spire e collettore. Macchina in continua: schematizzazione di indotto. Macchina in continua. Schematizzazione
Regole della mano destra. Macchina in continua con una spira e collettore. Macchina in continua con due spire e collettore. Macchina in continua: schematizzazione di indotto. Macchina in continua. Schematizzazione
Indice. pagina 2 di 10
 LEZIONE PROGETTAZIONE ORGANIZZATIVA DOTT.SSA ROSAMARIA D AMORE Indice PROGETTAZIONE ORGANIZZATIVA---------------------------------------------------------------------------------------- 3 LA STRUTTURA
LEZIONE PROGETTAZIONE ORGANIZZATIVA DOTT.SSA ROSAMARIA D AMORE Indice PROGETTAZIONE ORGANIZZATIVA---------------------------------------------------------------------------------------- 3 LA STRUTTURA
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
dieta vengono convertiti in composti dei corpi chetonici.
 Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Metabolismo degli aminoacidi Metabolismo degli aminoacidi Gli aminoacidi introdotti in eccesso con la dieta vengono convertiti in composti precursori del glucosio, degli acidi grassi e dei corpi chetonici.
Capitolo 2. Operazione di limite
 Capitolo 2 Operazione di ite In questo capitolo vogliamo occuparci dell operazione di ite, strumento indispensabile per scoprire molte proprietà delle funzioni. D ora in avanti riguarderemo i domini A
Capitolo 2 Operazione di ite In questo capitolo vogliamo occuparci dell operazione di ite, strumento indispensabile per scoprire molte proprietà delle funzioni. D ora in avanti riguarderemo i domini A
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I
 Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I BIOFISICA DELLE MEMBRANE BIOFISICA DELLE MEMBRANE Le funzioni biologiche di tutti gli organismi viventi si svolgono mediante
Corso di Laurea Magistrale in Medicina e Chirurgia Biofisica e Fisiologia I BIOFISICA DELLE MEMBRANE BIOFISICA DELLE MEMBRANE Le funzioni biologiche di tutti gli organismi viventi si svolgono mediante
Indice generale. OOA Analisi Orientata agli Oggetti. Introduzione. Analisi
 Indice generale OOA Analisi Orientata agli Oggetti Introduzione Analisi Metodi d' analisi Analisi funzionale Analisi del flusso dei dati Analisi delle informazioni Analisi Orientata agli Oggetti (OOA)
Indice generale OOA Analisi Orientata agli Oggetti Introduzione Analisi Metodi d' analisi Analisi funzionale Analisi del flusso dei dati Analisi delle informazioni Analisi Orientata agli Oggetti (OOA)
PROTEINE. sono COMPOSTI ORGANICI QUATERNARI
 PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
PROTEINE sono COMPOSTI ORGANICI QUATERNARI Unione di elementi chimici diversi Il composto chimico principale è il C (carbonio) Sono quattro gli elementi chimici principali che formano le proteine : C (carbonio),
BIOLOGIA GENERALE 22-24 ottobre 2007
 Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
Biologia generale Massolo Alessandro massolo@unifi.it; Tel. 347-9403330 BIOLOGIA GENERALE 22-24 ottobre 2007 Facoltà di Psicologia Tecniche di Psicologia Generale e Sperimentale Alessandro Massolo Dip.
CONDIZIONI GENERALI DI LAVORO PRESSO GLI STABILIMENTI AGUSTAWESTLAND ITALIA
 CONDIZIONI GENERALI DI LAVORO PRESSO GLI STABILIMENTI AGUSTAWESTLAND ITALIA 1. Nelle presenti Condizioni Generali, le parole elencate qui di seguito saranno da intendersi con i significati qui descritti:
CONDIZIONI GENERALI DI LAVORO PRESSO GLI STABILIMENTI AGUSTAWESTLAND ITALIA 1. Nelle presenti Condizioni Generali, le parole elencate qui di seguito saranno da intendersi con i significati qui descritti:
Il flusso dell informazione genetica. DNA -->RNA-->Proteine
 Il flusso dell informazione genetica DNA -->RNA-->Proteine Abbiamo visto i principali esperimenti che hanno dimostrato che il DNA è la molecola depositaria dell informazione genetica nella maggior parte
Il flusso dell informazione genetica DNA -->RNA-->Proteine Abbiamo visto i principali esperimenti che hanno dimostrato che il DNA è la molecola depositaria dell informazione genetica nella maggior parte
EMISSIONE E ASSORBIMENTO DI LUCE DA PARTE DELLA MATERIA
 EMISSIONE E ASSORBIMENTO DI LUCE DA PARTE DELLA MATERIA Poiché la luce è energia trasportata da oscillazioni del campo elettrico (fotoni) e la materia è fatta di particelle elettricamente cariche (atomi
EMISSIONE E ASSORBIMENTO DI LUCE DA PARTE DELLA MATERIA Poiché la luce è energia trasportata da oscillazioni del campo elettrico (fotoni) e la materia è fatta di particelle elettricamente cariche (atomi
DOMANDE A SCELTA MULTIPLA
 DOMANDE A SCELTA MULTIPLA Nome e cognome Alle domande rispondere con V (vero) o F (falso) nell apposita casella riservato alla giuria 1-I neuroni: Sono costituiti da corpo cellulare, assone e dendriti
DOMANDE A SCELTA MULTIPLA Nome e cognome Alle domande rispondere con V (vero) o F (falso) nell apposita casella riservato alla giuria 1-I neuroni: Sono costituiti da corpo cellulare, assone e dendriti
Laboratorio di Tecniche Microscopiche AA 2007-2008 Lezione 12 Marzo 2008 Ore 15-16
 Laboratorio di Tecniche Microscopiche AA 2007-2008 Lezione 12 Marzo 2008 Ore 15-16 L'immunoistochimica e' una tecnica ampiamente utilizzata per l'identificazione e la localizzazione di costituenti cellulari
Laboratorio di Tecniche Microscopiche AA 2007-2008 Lezione 12 Marzo 2008 Ore 15-16 L'immunoistochimica e' una tecnica ampiamente utilizzata per l'identificazione e la localizzazione di costituenti cellulari
6.1. Risultati simulazioni termovalorizzatore Osmannoro2000
 pag. 217 6. Risultati Di seguito si riportano i risultati relativi alle diverse simulazioni di diffusione atmosferica degli inquinanti effettuate. In particolare sono riportati i risultati sotto forma
pag. 217 6. Risultati Di seguito si riportano i risultati relativi alle diverse simulazioni di diffusione atmosferica degli inquinanti effettuate. In particolare sono riportati i risultati sotto forma
Funzioni della membrana plasmatica
 TRASPORTO M.P. -Omeostasi cellulare Funzioni della membrana plasmatica Mantenimento della conc. intracell. di ioni e molecole entro valori corretti Scambi interno - esterno Non c è consumo E Consumo E
TRASPORTO M.P. -Omeostasi cellulare Funzioni della membrana plasmatica Mantenimento della conc. intracell. di ioni e molecole entro valori corretti Scambi interno - esterno Non c è consumo E Consumo E
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Utilizzo ECG
 a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Utilizzo ECG 27/4/2006 Cuore come dipolo elettrico Il cuore considerato come un generatore elettrico complesso, in cui sono presenti
a.a. 2005/2006 Laurea Specialistica in Fisica Corso di Fisica Medica 1 Utilizzo ECG 27/4/2006 Cuore come dipolo elettrico Il cuore considerato come un generatore elettrico complesso, in cui sono presenti
UTILIZZATORI A VALLE: COME RENDERE NOTI GLI USI AI FORNITORI
 UTILIZZATORI A VALLE: COME RENDERE NOTI GLI USI AI FORNITORI Un utilizzatore a valle di sostanze chimiche dovrebbe informare i propri fornitori riguardo al suo utilizzo delle sostanze (come tali o all
UTILIZZATORI A VALLE: COME RENDERE NOTI GLI USI AI FORNITORI Un utilizzatore a valle di sostanze chimiche dovrebbe informare i propri fornitori riguardo al suo utilizzo delle sostanze (come tali o all
Modulo 11 biosegnalazione
 Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
«Macromolecole» Lipidi
 LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
LIPIDI (1) «Macromolecole» Lipidi Biotecnologie Miscellanea di molecole biologiche che condividono la proprietà di non essere solubili in acqua. Molecole idrofobiche. Es: Grassi Oli (grasso liquido a temperatura
COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA.2015-16
 COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA.2015-16 OMEOSTASI Fisiologia Umana La fisiologia è la scienza che studia i meccanismi di funzionamento degli organismi viventi. E una scienza integrata che
COMUNICAZIONE TRA CELLULA ED AMBIENTE FGE AA.2015-16 OMEOSTASI Fisiologia Umana La fisiologia è la scienza che studia i meccanismi di funzionamento degli organismi viventi. E una scienza integrata che