scaricato da
|
|
|
- Aureliana Morandi
- 7 anni fa
- Visualizzazioni
Transcript
1

2
3 ACIL CoA DEIDROGENASI
4 La membrana mitocondriale non è permeabile al NADH citosolico. Speciali sistemi navetta (shuttle) trasportano gli equivalenti riducenti dal NADH citosolico all interno dei mitocondri
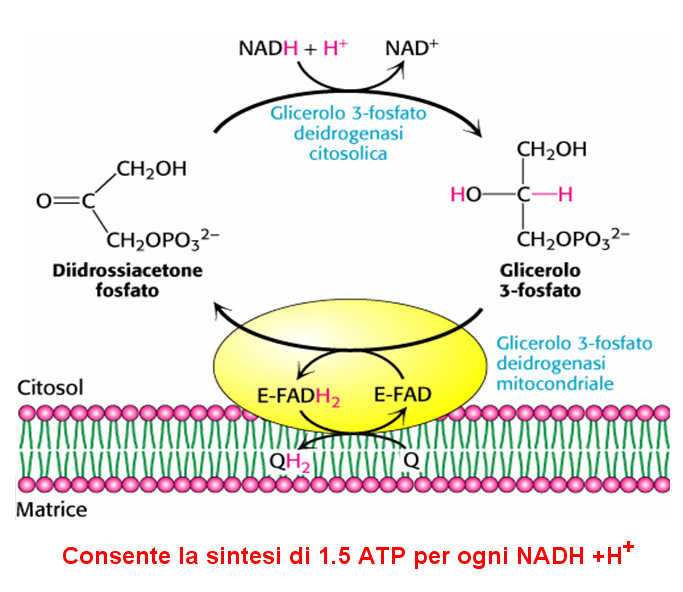
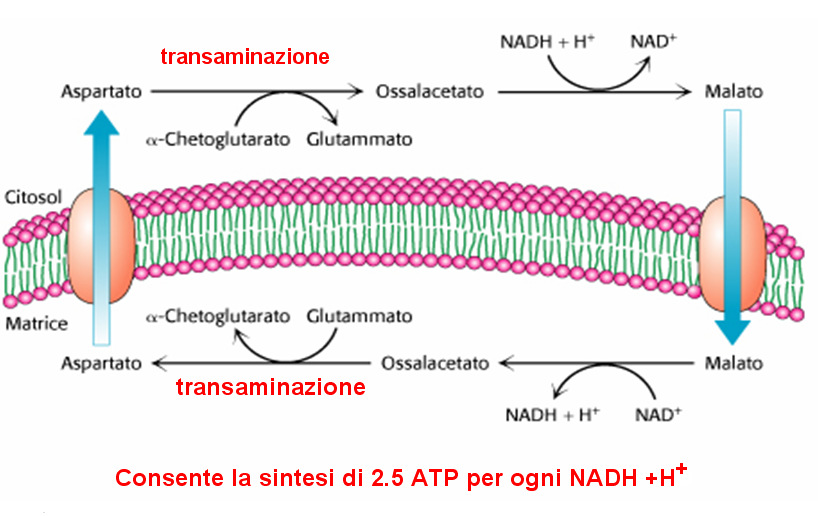
5 I principali sistemi navetta sono: Shuttle glicerolo fosfato (muscoli, tessuto nervoso) Shuttle malato-aspartato (fegato, reni cuore)
6
7
8 RESA IN ATP PER MOLECOLA DI Glc OSSIDATA Glicolisi Glc Glc-6P -1 ATP Frut-6P Frut1,6P -1 ATP 2 NADH (shuttle) ATP Fosforilazione a livello del substrato + 2 ATP Dec ossidativa piruvato 2 NADH + 5 ATP Ciclo di Krebs 6NADH + 15 ATP 2FAH2 + 3 ATP 2GTP + 2 ATP 30 o 32 ATP
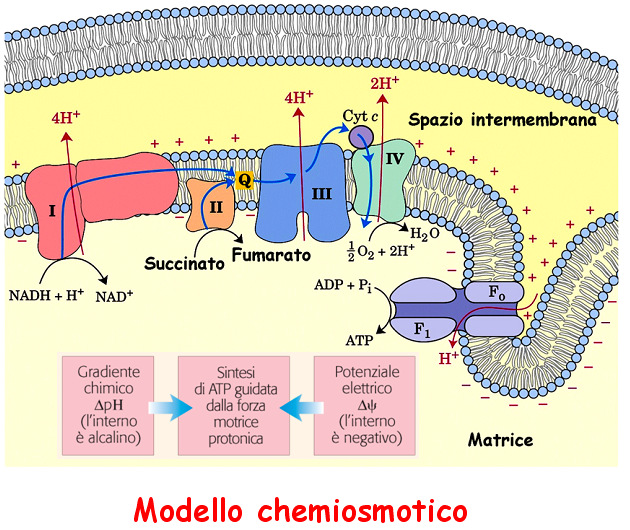
9 Il flusso degli e - è accompagnato da una traslocazione di H + che produce un gradiente chimico ( ph) e un gradiente elettrico ( ψ ψ). Si crea così un gradiente elettrochimico che genera la forza motrice protonica che fornisce l'energia per la sintesi di ATP quando gli H + ritornano nella o matrice attraverso una canale protonico, F, 1 associato con il complesso F dell ATP sintasi

10 Mitocondrio di Neurospora in un campione congelato in assenza di fissativi o coloranti che possano alterare le caratteristiche delle strutture interne. Wadsworth Center. Creste tubulari si connettono alla membrana interna attraverso passaggi stretti che possono limitare la velocità di equilibrio di H+ fra il lume delle creste e lo spazio intermembrana. Ci sono evidenze che i protoni estratti dalla matrice si disperdono lungo la superficie anionica delle membrane e solo lentamente si equilibrano con il grosso della fase circostante, massimizzando il reale gradiente di H+.
11
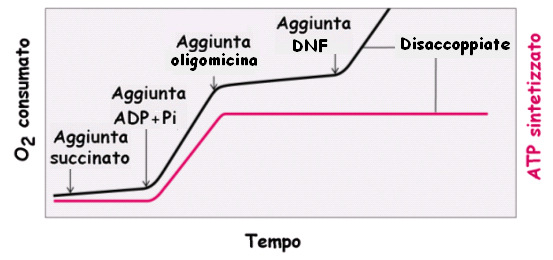
12
13 Il dinitrofenolo (DNF) è un disaccoppiante della FO Il DNF possiede un protone dissociabile ed è molto idrofobico: agisce trasportando H + attraverso la membr. mitoc. int. e dissipando il gradiente protonico

14 I mitocondri manipolati in modo da generare una differenza di ph e di carica tra le due superfici della membrana interna sintetizzano ATP in assenza di un substrato ossidabile: la forza motrice protonica è sufficiente a favorire la sintesi di ATP scaricato da
15 L ATP sintasi, detta anche complesso V, è formata da due componenti distinte: 1 F, una proteina periferica della membrana (il primo fattore identificato) o F, una proteina integrale della membrana (o sta ad indicare che questa è la porzione che conferisce al complesso la sensibilità all oligomicina) scaricato da
16 In laboratorio, piccole vescicole della membr. mitoc. 1 int. mancanti di F sono in grado di catalizzare il trasferimento degli e - ma non possono produrre ATP Quando F 1 purificato viene aggiunto alle vescicole esso si riassocia ad F o e può catalizzare la sintesi di ATP 1 La porzione F isolata catalizza l idrolisi (non la sintesi) 1
17 L ATP sintasi, detta anche complesso V, è formata da due componenti distinte: 1 F, una proteina periferica della membrana o F, una proteina integrale della membrana scaricato da

18 1 F ha una forma a pomello ed è costituita da un alternanza di subunità α e β disposte come gli spicchi di un arancia. La subunità γ attraversa il centro della struttura sferoidale costituendo l asta del pomello.
19 Più precisamente, γ contiene un dominio che attraversa F 1 ed un altro dominio che è associato ad una delle tre subunità β È proprio l associazione di γ con le subunità β che costringe ciascuna di esse ad assumere conformazioni differenti e con diverse affinità per i nucleotidi
20 1 F consiste in 9 subunità di 5 tipi diversi, con la 3 3 composizione α β γ δε γ ed ε costituiscono una sorta di gamba e piede

21 o Il canale protonico F composto da 3 tipi di è 2 10 subunità: ab c 2

22 o F Vista laterale Vista frontale Ciascuna delle dieci subunità c (organizzate in 2 cerchi concentrici) sono formate da due eliche transmembrana Le eliche N-terminali sono situate all interno; quelle C-terminali all esterno scaricato da
 Subunità dell anello c 0 F contiene l anello del canale protonico 1 di 10-14 subunità c La subunità F ha 5 tipi di catena polipetidica 3 Subunità (α3, β, γ, β δ, subunit ε), e presenta")
23 Subunità γ La subunità a si lega all anello esterno La colonna esteriore ha 1 subunità a 2 subunità b, e la subunità δ Subunità α ATP sintasi Il rotore è l anello is c & γε Il rimanente è stazionario (statore) Subunità dell anello c 0 F contiene l anello del canale protonico 1 di subunità c La subunità F ha 5 tipi di catena polipetidica 3 Subunità (α3, β, γ, β δ, subunit ε), e presenta attività ATPasica α& β sono membri della famiglia P-loop
24 Meccanismo di sintesi dell ATP i Un atomo di ossigeno dell ADP attacca il fosforo del P per formare un intermedio pentavalente che poi forma ATP e rilascio una molecola di H 2 O

25 Osservazione della rotazione dell ATP sintasi scaricato da marcato con fluoresceina e legato alla subunità γ La rotazione osservata mediante un microscopio a fluorescenza dopo idrolisi di ATP 3 L esamero α3β fissato su una superficie

26 Componenti dell unita di conduzione dei protoni scaricato da 2 α helices span the membrane Structure not yet directly observed
27 Subunit c Asp 61 in the middle of the second helix

28 Subunit a Proton enters Proton exits

29 Proton motion across the membrane drives rotation scaricato da Proton enters cytosolic half-channel, neutralizes Asp Proton enters matrix, resetting system Asp enters matrix half-channel

30 Proton path through the membrane Cytosolic half-channel Matrix half-channel
31 o La reazione sulla superficie di F F ha un G vicino allo 0, con una K eq di circa 1: si tratta, cioè, di una reazione reversibile F isolata catalizza invece l idrolisi, non la sintesi, 1 di ATP (tanto da venir chiamata F ATPasi) con un 0
32 L ATP sintasi rende l'atp stabile quanto i suoi prodotti di idrolisi ADP e Pi, legando molto saldamente l'atp e fornendo in questo modo l'energia necessaria per controbilanciare quella richiesta per la sintesi dell ATP stesso
33 F o F 1 lega l'atp con un affinità 7 10 più elevata di quella con cui lega l ADP Questa differenza di energia di legame è sufficiente a spostare l'equilibrio verso la formazione del prodotto ATP
34 Nella reazione catalizzata dall'atp sintasi la principale barriera energetica non è la sintesi di ATP ma il suo rilascio dall'enzima L'energia libera richiesta per il rilascio dell ATP è fornita dalla forza motrice protonica
35 Per la continua sintesi di ATP, l'enzima deve effettuare un ciclo tra una forma che lega saldamente ATP e una forma che lo rilascia Studi chimici e cristallografici sull ATP sintasi hanno rivelato le basi strutturali di questa alternanza di funzionalità
36 F 1 possiede tre siti di legame per nucleotidi adenilici, uno per ogni coppia αβ, con conformazioni non equivalenti I tre siti attivi di F 1 catalizzano a turno la sintesi di ATP (catalisi rotazionale)
37 Le tre conformazioni sono: - β-atp che lega saldamente l ATP - β-adp che lega debolmente l ATP - β -vuota che lega l ATP ancora più debolmente La forza motrice protonica provoca la rotazione del cilindro c e di γ (freccia verde) ed il contatto di questa con ciascuna coppia αβ
38 Ciò produce una modificazione conformazionale cooperativa: - il sito β-atp è convertito nella conformazione β-vuota e rilascia l'atp β -ADP è convertito in β -ATP, che provoca la condensazione di ADP e Pi per formare ATP - β -vuoto diventa β -ADP che lega debolmente ADP e Pi provenienti dal solvente L'ATP non può essere rilasciato da un sito finché ADP e Pi non sono legati all'altro scaricato da
39 Il passaggio di H + attraverso la porzione F o provoca la rotazione del cilindro costituito dalle subunità c e dalla subunità y ad esso attaccata. Ogni rotazione di 120 pone in contatto y con una diversa subunità β e la costringe ad assumere la conformazione β-vuota.
40 Le tre subunità β interagiscono tra di loro in modo tale che quando una assume la conformazione β-vuota, la subunità presente su di un lato deve assumere la forma β-adp mentre quella sull'altro lato acquista la conformazione β-atp
41 Per ogni rotazione completa di γ, ogni subunità β compie un ciclo attraverso le tre possibili conformazioni e vengono sintetizzate e rilasciate dalla superficie dell enzima tre molecole di ATP
42 Per molti anni i libri di testo hanno riportato un rapporto P/O = 3 quando il donatore di e - era il NADH un rapporto P/O = 2 quando il donatore di e - era il succinato L'introduzione della teoria chemiosmotica consentì di ipotizzare che il rapporto P/O non fosse necessariamente un numero intero.
43 + Si è stabilito che il numero di H - pompati fuori per coppia di e trasportati è di 10 per il NADH e di 6 per il succinato. + Si è concordato che il numero di H richiesti per la sintesi di una molecola di ATP è di 4, di cui uno è usato per trasportare Pi, ATP e ADP attraverso la membr. mitoc.
44 Il rapporto P/O è 2.5 (10:4) quando il donatore di e - è il NADH Il rapporto P/O è 1.5 (6:4) quando il donatore di e - è il succinato
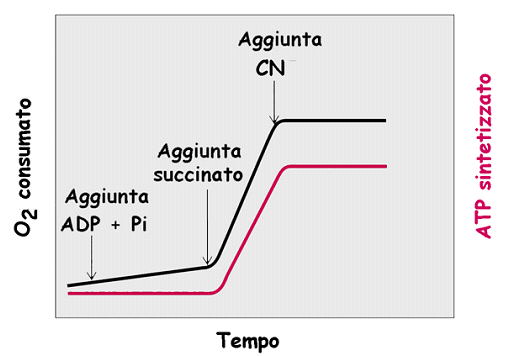
45 La funzione dell'atp sintasi è strettamente integrata con quella di due traslocasi: - t. dei nucleotidi adenilici (antiporto) - t. del fosfato (simporto) Nel loro insieme le tre proteine costituiscono un complesso, l ATP sintasoma

46
47 La traslocasi dei nucleotidi adenilici lega una molecola 3- di ADP nello spazio intermembrana e la trasporta nella matrice, in antiporto con 4- una molecola di ATP Questo trasporto di 4 cariche negative all'esterno e 3 all'interno è favorito dal fatto che la matrice è
48 La fosfato traslocasi catalizza il simporto nella matrice di uno ione H 2 PO 4 - e di uno ione H +. Anche questo processo è favorito dal gradiente + protonico: un H passa dal lato p al lato n della membrana interna, consumando parte dell'energia prodotta dal trasferimento degli e -.
49 - 2 Per ogni coppia di e vengono prodotte che arriva all O 1.5 molecole di ATP nello shuttle del glicerolo 3P 2.5 molecole di ATP nello shuttle Mal-Asp

50
51 L'ossidazione completa di una molecola di Glc produce 30/32 molecole di ATP, mentre la glicolisi anaerobia ne produce solo 2 La via ossidativa aerobia accoppiata alla FO è dunque la fonte primaria dell ATP prodotto dal catabolismo
52 La velocità di ossidazione delle sostanze nutrienti è finemente regolata in modo tale da produrre una quantità di ATP adeguata alle necessità energetiche della cellula
53 La velocità della respirazione (consumo di O 2 ) nei mitocondri è altamente controllata e strettamente dipendente dalla concentrazione dell ADP (controllo dell'accettore della respirazione) Avendo a disposizione più ADP per la FO ossidativa, la velocità della respirazione aumenta, determinando la rigenerazione dell'atp
54
55 Nei neonati e negli animali in letargo vi è un particolare tipo di tessuto adiposo, chiamato grasso bruno (per la per la presenza di numerosi mitocondri) Questi mitocondri possiedono nella loro membrana interna una proteina speciale, la termogenina, chiamata anche proteina disaccoppiante scaricato da
56 Questa proteina costituisce una via di ritorno o 1 per gli H + che non attraversano F F per effetto di questo cortocircuito, l'energia delle ossidazioni non viene utilizzata per produrre ATP ma viene dissipata come calore necessario a mantenere il corpo a temperatura costante

57
58 Quando una cellula è ischemica (privata di O 2 ), come avviene durante un attacco di cuore, il - + trasferimento di e all O 2 e il pompaggio di H si interrompono, e la forza motrice protonica collassa immediatamente In queste condizioni, l ATP sintasi potrebbe agire in modo opposto, idrolizzando l'atp per pompare gli H + nella direzione inversa e causando una disastrosa riduzione dei livelli di ATP scaricato da
59 Ciò viene impedito da una piccola (84 aa) proteina inibitrice, IF 1, che lega simultaneamente due molecole di ATP sintasi, inibendo la loro attività ATPasica L'inibitore è attivo solo nella sua forma dimerica, che si produce a valori di ph inferiori a 6,5. In una cellula privata di O 2, la glicolisi anaerobia fa abbassare il ph, favorendo la dimerizzazione di IF 1 Quando viene ripristinato il metabolismo aerobio, il ph sale, il dimero IF 1 si destabilizza e l'inibizione dell'atp sintasi viene rimossa. scaricato da
60 SVANTAGGI DELLA F.O. Specie reattive dell ossigeno (ROS). 2 2 O + e- O Superossidodismutasi.
FADH ADP + Pi ATP...29 AG ' = -30,5 kj/mol...29 Resa di ATP per l ossidazione completa del glucosio...31
 FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
FOSFORILAZIONE OSSIDATIVA FOSFORILAZIONE OSSIDATIVA...1 Glicolisi e ciclo di Krebs...3 il NADH il NADPH e il FADH2 sono traportatori solubili di elettroni...4 LA FOSFORILAZIONE OSSIDATIVA...5 Il mitocondrio...5
L ossidazione completa del glucosio da parte dell O 2. può essere suddivisa in due semi-reazioni
 L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
L ossidazione completa del glucosio da parte dell O 2 C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O può essere suddivisa in due semi-reazioni C 6 H 12 O 6 + 6 O 2 6 CO 2 + 24 H + + 24 e - (si ossidano gli atomi
matrice Sp. Interm. Il gambo (γε) cilindro C10 ruota con esso H + H+ H + H + H + H H + H H + H + H + H +
 cilindro C10 ruota con esso H + H+ H + H + H + H H + H H + H + H + H +") I protoni si muovono secondo gradiente attraverso un canale d ingresso creato dalla subunità a e raggiungono il cilindro c 10. Quando una subunità c lega 1 un altro è rilasciato nella matrice lasciando
I protoni si muovono secondo gradiente attraverso un canale d ingresso creato dalla subunità a e raggiungono il cilindro c 10. Quando una subunità c lega 1 un altro è rilasciato nella matrice lasciando
Anatomia biochimica di un mitocondrio
 Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Anatomia biochimica di un mitocondrio Le involuzioni(creste)aumentano considerevolmente l area della superficie della membrana interna. La membrana interna di un singolo mitocondrio può avere anche più
Fosforilazione ossidativa
 Chimica Biologica A.A. 2010-2011 Fosforilazione ossidativa Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Teoria Chemiosmotica ΔG = 2.3 RT (ph (matrice) -ph (spazio
Chimica Biologica A.A. 2010-2011 Fosforilazione ossidativa Marco Nardini Dipartimento di Scienze Biomolecolari e Biotecnologie Università di Milano Teoria Chemiosmotica ΔG = 2.3 RT (ph (matrice) -ph (spazio
Come le cellule traggono energia dal cibo: produzione di ATP
 Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
Come le cellule traggono energia dal cibo: produzione di ATP L energia è contenuta nei legami chimici delle molecole nutritive; la cellula estrae questa energia e la conserva nell ATP: respirazione cellulare
CHIMICA BIOLOGICA. Seconda Università degli Studi di Napoli. DiSTABiF. Corso di Laurea in Scienze Biologiche. Insegnamento di. Anno Accademico
 Seconda Università degli Studi di Napoli DiSTABiF Prof. Antimo Di Maro Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Anno Accademico 2015-16 Lezione 15 Catena di trasporto degli
Seconda Università degli Studi di Napoli DiSTABiF Prof. Antimo Di Maro Corso di Laurea in Scienze Biologiche Insegnamento di CHIMICA BIOLOGICA Anno Accademico 2015-16 Lezione 15 Catena di trasporto degli
Prof. Maria Nicola GADALETA FOSFORILAZIONE OSSIDATIVA 4
 Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
Prof. Maria Nicola GADALETA E-mail: m.n.gadaleta@biologia.uniba.it Facoltà di Scienze Biotecnologiche Corso di Laurea in Biotecnologie Sanitarie e Farmaceutiche Biochimica e Biotecnologie Biochimiche FOSFORILAZIONE
fornire energia chimica in vettori attivati ATP e NADH e NADPH e FADH.
 Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
Cibo ed Energia Il glucosio e altre molecole nutritive per esempio i grassi vengono demolite attraverso un ossidazione graduale controllata per fornire energia chimica in una forma utilizzabile dalla cellula,
COME VIENE METABOLIZZATA QUESTA MASSICCIA QUANTITA DI ENERGIA? Trasformazione di energia potenziale di riduzione. energia libera di fosforilazione
 Trasformazione di energia potenziale di riduzione in energia libera di fosforilazione Un uomo adulto produce in media abbastanza energia da sintetizzare ogni giorno una quantità di ATP pari al suo peso
Trasformazione di energia potenziale di riduzione in energia libera di fosforilazione Un uomo adulto produce in media abbastanza energia da sintetizzare ogni giorno una quantità di ATP pari al suo peso
NADH FADH 2 (trasportatori ridotti di elettroni) Catena respiratoria (trasferimento degli
 Catena respiratoria (trasferimento degli") NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
NADH FADH 2 (trasportatori ridotti di elettroni) Fase 3 Trasferimento degli elettroni e fosforilazione ossidativa Catena respiratoria (trasferimento degli elettroni) Nicotinamide adenin dinucleotide (NAD
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA
 I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
I MITOCONDRI: LE CENTRALI ENERGETICHE DELLA CELLULA MITOCONDRIO OSSERVATO AL SEM Localizzazione dei mitocondri SONO IN GENERE PIÙ ABBONDANTI DELLE ZONE DELLA CELLULA CHE RICHIEDONO MAGGIORI QUANTITA DI
Fosforilazione ossidativa
 Fosforilazione ossidativa H 2 2H + + 2e - NADH+H + NAD + + 2H + +2e - FADH 2 FAD + 2H + + 2e - ½O 2 + 2H + + 2e - H 2 O + Calore ½ O 2 + 2H + + 2e - H 2 O + ATP 1 La fosforilazione ossidativa avviene nella
Fosforilazione ossidativa H 2 2H + + 2e - NADH+H + NAD + + 2H + +2e - FADH 2 FAD + 2H + + 2e - ½O 2 + 2H + + 2e - H 2 O + Calore ½ O 2 + 2H + + 2e - H 2 O + ATP 1 La fosforilazione ossidativa avviene nella
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa
 e Fosforilazione ossidativa") Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
Catena di trasporto degli elettroni (catena respiratoria) e Fosforilazione ossidativa NADH e FADH2 (accettori universali di e-) formati nella glicolisi e nel ciclo di Krebs (e nell ossidazione degli acidi
Il ciclo di Krebs e la fosforilazione ossidativa
 Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
Il ciclo di Krebs e la fosforilazione ossidativa La respirazione cellulare Sono i processi molecolari in cui è coinvolto il consumo di O 2 e la formazione di CO 2 e H 2 O da parte della cellula. E suddivisa
Cap.19. Ciclo di Krebs. o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico
 Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
Cap.19 Ciclo di Krebs o Ciclo degli acidi Tricarbossilici o Ciclo dell acido Citrico Acetil-CoA OSSIDAZIONE DEL GLUCOSIO C 6 H 12 O 6 (glucosio) + 6O 2.. 6CO 2 + 6H 2 O I tappa: GLICOLISI 2 + 2 H 2 O II
METABOLISMO CELLULARE
 METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
METABOLISMO CELLULARE Struttura dell ATP (Adenosintrifosfato) Adenina (base azotata), Ribosio (zucchero) e un gruppo fosforico ATP Il legame covalente tra i gruppi fosforici si spezza facilmente liberando
Cap.21 CATABOLISMO DEI LIPIDI
 Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
Cap.21 CATABOLISMO DEI LIPIDI Glucosio= forma di energia immediatamente disponibile LIPIDI= Deposito energetico utilizzabile da tutti i tessuti, tranne cervello e globuli rossi Rispetto ai carboidrati,
FOSFORILAZIONE OSSIDATIVA
 FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
FOSFORILAZIONE OSSIDATIVA Lo scopo ultimo dei processi metabolici ossidativi è quello di canalizzare l energia contenuta nello scheletro carbonioso di zuccheri, ac. grassi e amminoacidi nella sintesi di
Mitocondri. -sono visibili al MO (Ø 0,5 µ e lunghezza da 1 a 6 µ) -assenti nei batteri e presenti in tutte le cellule eucariotiche
 -assenti nei batteri e presenti in tutte le cellule eucariotiche") Mitocondri sono organelli deputati alla produzione dell energia, capaci di ossidare i prodotti dell assorbimento intestinale degradandoli e di convertire l energia liberata in ATP processo chiamato respirazione
Mitocondri sono organelli deputati alla produzione dell energia, capaci di ossidare i prodotti dell assorbimento intestinale degradandoli e di convertire l energia liberata in ATP processo chiamato respirazione
AGISCONO NEL SISTEMA NAVETTA 2 ISOFORME DELL ENZIMA: UNA FORMA MITOCONDRIALE ( MEMBRANA MIT. INTERNA) FAD-dipendente
 FAD-dipendente") METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
METABOLISMO AEROBIO I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione mitocondriale attiva), possono trasferire elettroni all interno del mitocondrio attraverso i
I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia
 I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia Edizione 2016-2017 Metabolismo: equilibrio e omeostasi dei processi cellulari Lezione: Le principali vie metaboliche, il
I Lincei per una nuova didattica nella Scuola: una rete nazionale Polo di Brescia Edizione 2016-2017 Metabolismo: equilibrio e omeostasi dei processi cellulari Lezione: Le principali vie metaboliche, il
Capitolo 6 La respirazione cellulare
 Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
Capitolo 6 La respirazione cellulare Introduzione alla respirazione cellulare 6.1 La respirazione polmonare rifornisce le nostre cellule di ossigeno ed elimina diossido di carbonio La respirazione polmonare
il valore 3 o 5 dipende dal meccanismo usato per trasportare gli elettroni del NADH dal citosol alla matrice mitocondriale
 = 976 kj/mole il valore 3 o 5 dipende dal meccanismo usato per trasportare gli elettroni del NADH dal citosol alla matrice mitocondriale Energia contenuta nel glucosio = 2840 kj/mole Recupero = 32 x 30,5
= 976 kj/mole il valore 3 o 5 dipende dal meccanismo usato per trasportare gli elettroni del NADH dal citosol alla matrice mitocondriale Energia contenuta nel glucosio = 2840 kj/mole Recupero = 32 x 30,5
Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la
 Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
Metabolismo Attività cellulare altamente coordinata svolta da sistemi multienzimatici, con i seguenti scopi: ottenere energia chimica dall ambiente attraverso la degradazione di nutrienti operare la sintesi
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Complesso della Piruvato deidrogenasi
COMBUSTIONE = OSSIDAZIONE SOSTANZA ORGANICA. Legno + O2 -> CO2 + H2O + calore
 Respirazione COMBUSTIONE = OSSIDAZIONE SOSTANZA ORGANICA Legno + O2 -> CO2 + H2O + calore O2 ---> CO2 + H2O + calore Cumultive C-CO 2 (% of initial C) Weight remaining (%) Decomposition as mass loss 100
Respirazione COMBUSTIONE = OSSIDAZIONE SOSTANZA ORGANICA Legno + O2 -> CO2 + H2O + calore O2 ---> CO2 + H2O + calore Cumultive C-CO 2 (% of initial C) Weight remaining (%) Decomposition as mass loss 100
Trasporto degli elettroni e fosforilazione ossidativa
 Trasporto degli elettroni e fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobici Tutte le tappe enzimatiche della degradazione
Trasporto degli elettroni e fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobici Tutte le tappe enzimatiche della degradazione
CICLO DI KREBS (o DELL ACIDO CITRICO)
") CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
CICLO DI KREBS (o DELL ACIDO CITRICO) È un processo ossidativo che ha un ruolo centrale nel metabolismo energetico delle cellule eucariotiche. Avviene nella matrice mitocondriale. È alimentato soprattutto
Seminario. Ciclo di Krebs 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A
 Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
Seminario PASSO PER PASSO 1. La citrato sintasi forma citrato a partire dall ossaloacetato e dall acetilcoenzima A Il ciclo dell acido citrico inizia con la reazione di condensazione di un unità con 4
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) La gluconeogenesi utilizza il piruvato e altri composti a
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia.
 La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
La glicolisi non è l unica via catabolica in grado di produrre energia. Se il suo prodotto viene ulteriormente ossidato si ottiene molta più energia. Il ciclo dell acido citrico è una via metabolica centrale
Il metabolismo cellulare
 Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
Il metabolismo cellulare 1 Il metabolismo cellulare CATABOLISMO Processo esoergonico (per produrre energia) Nutriliti NAD FAD NADP e - +H + ANABOLISMO Processo endoergonico Biopolimeri (x es. proteine)
Digestione e assorbimento dei lipidi. β-ossidazione degli acidi grassi
 Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
Digestione e assorbimento dei lipidi β-ossidazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500
INTRODUZIONE AL METABOLISMO. dal gr. metabolè = trasformazione
 INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
INTRODUZIONE AL METABOLISMO dal gr. metabolè = trasformazione IL Metabolismo Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in una cellula o in un organismo. E costituito da
BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA
 Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
Cap.15 BIOENERGETICA IL METABOLISMO RISULTA DALL INSIEME DELLE REAZIONI CHIMICHE CHE PERMETTONO AI SISTEMI VIVENTI DI UTILIZZARE ENERGIA E MATERIA I Legge della Termodinamica: Il contenuto energetico dell
METABOLISMO DEL GLICOGENO
 METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
METABOLISMO DEL GLICOGENO GLICOGENO: Polisaccaride di riserva del regno animale Organi principali: FEGATO MUSCOLO SCHELETRICO Glicogeno Glucosio Glucosio-6P Via del pentoso fosfato Glucosio Fruttosio-6P
2 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA
 INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
INCONTRO: LA PRODUZIONE DI ENERGIA NELLA CELLULA 1 INTRODUZIONE 1 L energia chimica Esistono diversi tipi di energia e una tra queste è l energia chimica: un tipo di energia che possiedono tutte le molecole
scaricato da www.sunhope.it
 CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
CICLO DEGLI ACIDI TRICARBOSSILICI o CICLO DELL ACIDO CITRICO o CICLO DI KREBS scaricato da www.sunhope.it IL CICLO DI KREBS È LA VIA OSSIDATIVA FINALE DEL CATABOLISMO DI GLUCIDI LIPIDI PROTEINE IL CICLO
INTRODUZIONE AL METABOLISMO
 INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
INTRODUZIONE AL METABOLISMO METABOLISMO Il metabolismo (dal greco metabolé = trasformazione) è il complesso delle reazioni chimiche coordinate ed integrate che avvengono in una cellula. Il metabolismo
Il metabolismo cellulare
 Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Il metabolismo cellulare L obesità è problema sanitario e sociale Tessuto adiposo bianco e bruno È tutta una questione di energia: Se si assumono più molecole energetiche di quelle che ci servono per costruire
Utilizzo del glucosio: la glicolisi
 Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Utilizzo del glucosio: la glicolisi GLUCOSIO Sistema rapido, reversibile, GLICOLISI avviene anche in assenza di ossigeno. Produce poca energia OSSIDAZIONE PIRUVATO Fermentazione LATTATO ACETATO CICLO DI
Respirazione cellulare
 Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
Respirazione cellulare L equazione generale della respirazione C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 36 ATP 1 molec zucchero 6 molec ossigeno 6 molec anidride carbonica 6 molec acqua + = + + 36 molecole
(2 x) (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi
 (2 x) (2 x) Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi. Gliceraldeide 3-fosfato deidrogenasi") Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Il NADH prodotto in questa reazione DEVE essere ri-ossidato affinché la glicolisi non si fermi (2 x) (2 x) Gliceraldeide 3-fosfato Fosfato inorganico Gliceraldeide 3-fosfato deidrogenasi Reazione di ossidoriduzione:
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO)
") RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
RESPIRAZIONE CELLULARE (METABOLISMO DEL GLUCOSIO) LA GLICOLISI: dopo il ciclo di Calvin, che avviene nelle cellule autotrofe delle piante, il glucosio prodotto va a demolirsi per produrre energia nelle
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI
 METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
METABOLISMO OSSIDATIVO VIE METABOLICHE CONVERGENTI Nelle piante l ossidazione degli acidi grassi avviene nei perossisomi delle foglie e nei gliossisomi dei semi Nelle cellule animali il sito principale
DESTINI DEL PIRUVATO
 DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
DESTINI DEL PIRUVATO LA GLICOLISI RILASCIA SOLO UNA PICCOLA PARTE DELL ENERGIA TOTALE DISPONIBILE NELLA MOLECOLA DI GLUCOSIO Le due molecole di piruvato prodotte dalla glicolisi sono ancora relativamente
Cap.16 GLICOLISI. Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP H 2 O 2 NAD + 2 NADH + 2H +
 Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Cap.16 GLICOLISI Glucosio + 2 ADP + 2 Pi + 2 NAD + 2 Piruvato + 2 ATP + 2 H 2 O+ 2 NADH + 2H + 2 ADP + 2 Pi 2 ATP 2 + 2 H 2 O 2 NAD + 2 NADH + 2H + Via metabolica in 10 tappe (reazioni) La glicolisi può
Metabolismo: Introduzione
 Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Metabolismo: Introduzione METABOLISMO Insieme delle reazioni chimiche coordinate e integrate che hanno luogo in tutte le cellule C A T A B O L I S M O Ossidazioni esoergoniche Alimenti: Carboidrati Lipidi
Energia e metabolismi energetici
 Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
Energia e metabolismi energetici L energia è la capacità di produrre lavoro Tutti gli organismi hanno bisogno di energia per vivere. L energia è definita come la capacità di effettuare un lavoro L energia
Lezione 7. Il metabolismo terminale e la produzione di ATP
 Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
Lezione 7 Il metabolismo terminale e la produzione di ATP 1 Il metabolismo terminale Catabolismo Lipidi Amminoacidi Carboidrati convergono sul metabolismo terminale, e producono una stessa molecola: l
catabolismo anabolismo
 Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
Il metabolismo è la somma di tutte le trasformazioni chimiche che avvengono in un organismo o in una cellula Il catabolismo è l insieme delle reazioni che portano alla scissione di molecole complesse (ricche
BETA OSSIDAZIONE DEGLI ACIDI GRASSI
 Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
Mitocondri SEMINARIO BETA OSSIDAZIONE DEGLI ACIDI GRASSI http://oregonstate.edu/instruct/bb350/textmaterials/21/slide08.jpg 1 Acidi grassi [1] Sono le principali fonti di energia per alcuni tessuti (es.
MODULO 6 FOSFORILAZIONE OSSIDATIVA REAZIONI DI OSSIDORIDUZIONE
 MODULO 6 FOSFORILAZIONE OSSIDATIVA REAZIONI DI OSSIDORIDUZIONE Introduzione I sistemi viventi possono essere distinti, in base al consumo d ossigeno, in organismi aerobi ed organismi anaerobi. I primi
MODULO 6 FOSFORILAZIONE OSSIDATIVA REAZIONI DI OSSIDORIDUZIONE Introduzione I sistemi viventi possono essere distinti, in base al consumo d ossigeno, in organismi aerobi ed organismi anaerobi. I primi
CATENA DI TRASPORTO DEGLI ELETTRONI
 2 Parte, Mitocondri CATENA DI TRASPORTO DEGLI ELETTRONI http://www.personal.kent.edu/~cearley/pchem/krebs/etc.png http://lh3.ggpht.com/ TXGk 7CtnfE/Us6AfOto9fI/AAAAAAAAA38/TCEjo91 _B8/citric_acid_cycle%25255B12%25255D.jpg?imgmax=800
2 Parte, Mitocondri CATENA DI TRASPORTO DEGLI ELETTRONI http://www.personal.kent.edu/~cearley/pchem/krebs/etc.png http://lh3.ggpht.com/ TXGk 7CtnfE/Us6AfOto9fI/AAAAAAAAA38/TCEjo91 _B8/citric_acid_cycle%25255B12%25255D.jpg?imgmax=800
6 H 2. con G=-686 kcal/mole di H 12 O 6 O + 6 CO O 2. glucosio La respirazione avviene in tre stadi principali; ognuno di questi
 La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
La respirazione La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O 2 Il
scaricato da 1
 scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
scaricato da www.sunhope.it 1 La decarbossilazione ossidativa del piruvato rappresenta il legame tra glicolisi e ciclo di Krebs Questa reazione, come tutto il ciclo di Krebs, avviene nella matrice mitocondriale
ENERGIA LIBERA DI GIBBS (G)
") METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
METABOLISMO: Descrive tutte le numerose reazioni con cui le molecole biologiche sono sintetizzate e degradate, e che permettono di ricavare, accumulare e utilizzare energia Ogni reazione metabolica comporta
Proprietà di permeabilità della membrana
 Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
Proprietà di permeabilità della membrana Il Trasporto attraverso le membrane La fase lipidica delle membrane le rende impermeabili alla maggior parte degli ioni e delle sostanze polari. Questi composti
Il trasporto del glucosio
 Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Il trasporto del glucosio Il glucosio non può diffondere direttamente nelle cellule Esso entra nelle cellule mediante due meccanismi di trasporto: a) diffusione facilitata Na + indipendente b) co-trasporto
Ossidazione del glucosio
 Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
Ossidazione del glucosio Vie metaboliche all uso del glucosio Glicolisi e Fermentazioni 1 Reazione di ossidazione C 6 H 12 O 6 + 6 O 2 => 6 CO 2 + 6 H 2 O + 686 Kcal/mole Il glucosio reagisce direttamente
Glycogen Synthesis Glicogeno sintesi UTP UDP + 2 P i glycogen (n) + glucose-1-p glycogen (n + 1) Glycogen Phosphorylase P i Glicogeno lisi
 + glucose-1-p glycogen (n + 1) Glycogen Phosphorylase P i Glicogeno lisi") Glycogen Synthesis Glicogeno sintesi UTP UDP + 2 P i glycogen (n) + glucose-1-p glycogen (n + 1) Glycogen Phosphorylase P i Glicogeno lisi Se entrambe le vie fossero attive simultaneamente nella cellula
Glycogen Synthesis Glicogeno sintesi UTP UDP + 2 P i glycogen (n) + glucose-1-p glycogen (n + 1) Glycogen Phosphorylase P i Glicogeno lisi Se entrambe le vie fossero attive simultaneamente nella cellula
Catena Respiratoria e Fosforilazione Ossidativa. glucosio. piruvato H + Ac-CoA O 2. e - NADH. ADP P i H 2 O H + H + ATP NADH
 Catena Respiratoria e Fosforilazione Ossidativa glucosio piruvato H + H + Ac-CoA O 2 H 2 O H + e - NADH ATP ADP P i NADH Catena di Trasporto degli Elettroni glicolisi / ciclo di Krebs / -ox acidi grassi:
Catena Respiratoria e Fosforilazione Ossidativa glucosio piruvato H + H + Ac-CoA O 2 H 2 O H + e - NADH ATP ADP P i NADH Catena di Trasporto degli Elettroni glicolisi / ciclo di Krebs / -ox acidi grassi:
Connessioni. Le cellule cardiache sono unite tra loro da parti più ispessite del sarcolemma dette: Gap - junction
 Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
Muscolo cardiaco Le fibrocellule cardiache sono simili alle fibrocellule muscolari di tipo I (fibre rosse), ma si differenziano, infatti: Sono più piccole Non sono polinucleate Minor massa fibrillare (50%
REGOLAZIONE DEL CICLO TCA
 REGOLAZIONE DEL CICLO TCA Respirazione cellulare Fase 1 1) Catabolismo di acidi grassi 2) Glucosio 3) Amminoacidi Acetil-CoA Fase2 Ciclo dell acido citrico: ossidazione dell acetil CoA con produzione di
REGOLAZIONE DEL CICLO TCA Respirazione cellulare Fase 1 1) Catabolismo di acidi grassi 2) Glucosio 3) Amminoacidi Acetil-CoA Fase2 Ciclo dell acido citrico: ossidazione dell acetil CoA con produzione di
MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE
 BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
BIOENERGETICA MANTENIMENTO DELLA STRUTTURA CRESCITA SVILUPPO RIPRODUZIONE GLI ORGANISMI VIVENTI POSSONO UTILIZZARE SOLO DUE FORME DI ENERGIA: LUMINOSA (radiazioni di determinate lunghezze d onda) (ORGANISMI
Biosintesi dei carboidrati
 Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Biosintesi dei carboidrati Gluconeogenesi: sintesi di glucosio da precursori non saccaridici La riserva di glucosio dell organismo (glucosio + glicogeno) è sufficiente per circa 1 giorno. La via gluconeogenetica
Degradazione degli acidi grassi
 Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
Degradazione degli acidi grassi I grassi della dieta sono assorbiti nell intestino tenue Il diametro della particella dei chilomicroni varia da circa 100 a circa 500 nm. Mobilizzazione dei triacilgliceroli
da: Nelson & Cox (IV Ed.)
") da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
da: Nelson & Cox (IV Ed.) da: Nelson & Cox (IV Ed.) Il prodotto finale dopo 7 cicli è il palmitil ACP. Il processo di allungamento si ferma a C 16 e l acido palmitico viene liberato dalla molecola di ACP
Corso di Biochimica. Destino catabolico del piruvato. Prof. Paolo Cirri. Dipartimento Scienze Biomediche Sperimentali e Cliniche Università di Firenze
 Corso di Biochimica Prof. Paolo Cirri Dipartimento Scienze Biomediche Sperimentali e Cliniche Università di Firenze Orario ricevimento: martedì 14,30-16,30 Diapositive lezioni: http://bioserver2.sbsc.unifi.it/
Corso di Biochimica Prof. Paolo Cirri Dipartimento Scienze Biomediche Sperimentali e Cliniche Università di Firenze Orario ricevimento: martedì 14,30-16,30 Diapositive lezioni: http://bioserver2.sbsc.unifi.it/
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico
 Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Il ciclo degli acidi tricarbossilici o ciclo dell acido citrico Dalla glicolisi alla fosforilazione ossidativa NAD + e NADP + Sono coinvolti nelle reazioni di trasferimento dell idruro H: -. Tutte le reazioni
Laboratorio Fitness & Wellness
 Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
Laboratorio Fitness & Wellness Anno Accademico 2015/2016 Ewan Thomas, MSc Principi di Bioenergetica e Biomeccanica Metabolismo Catabolismo Processi di degradazione che permettono di liberare energia Anabolismo
Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale
 Stechiometria della reazione finale") Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
Ciclo di Krebs 1 Complesso della piruvato deidrogenasi (1) Stechiometria della reazione finale 2 Complesso della piruvato deidrogenasi (2) FAD Diidrolipoil deidrogenasi Piruvato decarbossilasi E 3 Diidrolipoil
ENZIMI. Durante la reazione l enzima può essere temporaneamente modificato ma alla fine del processo ritorna nel suo stato originario, un enzima viene
 ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari, e lavorano in condizioni blande di temperatura e ph (sono in grado di aumentare la velocità delle
ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari, e lavorano in condizioni blande di temperatura e ph (sono in grado di aumentare la velocità delle
La membrana e il trasporto di membrana
 Clostridium botulinum. Batterio strettamente anaerobio che produce una tossina termostabile che è estremamente neurotossica per l uomo. Può proliferare nelle conserve vegetali e nelle conserve sott olio.
Clostridium botulinum. Batterio strettamente anaerobio che produce una tossina termostabile che è estremamente neurotossica per l uomo. Può proliferare nelle conserve vegetali e nelle conserve sott olio.
Metabolismo fermentativo
 Metabolismo fermentativo Anaerobio (non richiede ossigeno) Energeticamente poco efficiente (se confrontata alla respirazione) La catena di trasferimento degli elettroni è molto limitata: donatori ed accettori
Metabolismo fermentativo Anaerobio (non richiede ossigeno) Energeticamente poco efficiente (se confrontata alla respirazione) La catena di trasferimento degli elettroni è molto limitata: donatori ed accettori
IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI
 GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
GLUCONEOGENESI IL GLUCOSIO PUO ESSERE SINTETIZZATO DA PRECURSORI NON GLUCIDICI Quando la quantità di glucosio fornito dagli alimenti o dalle riserve è insufficiente, i diversi organismi sintetizzano glucosio
IL METABOLISMO. Dal carbonio agli OGM Capitolo 2
 IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
IL METABOLISMO Dal carbonio agli OGM Capitolo 2 IL METABOLISMO L'insieme delle trasformazioni chimiche ed energetiche che si verificano nelle cellule di un organismo vivente e ne garantiscono la conservazione,
Fosforilazione a livello del substrato
 10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
10 piruvato chinasi Fosfoenolpiruvato Rib Adenina Piruvato ADP Rib ATP Adenina Fosforilazione a livello del substrato 3 Fosfofruttochinasi-1 (PFK-1) Fruttosio 6-fosfato Fruttosio 1,6-bisfosfato 1 esochinasi
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI
 METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
METABOLISMO del GLUCOSIO: GLUCONEOGENESI e VIA DEI PENTOSI Glicolisi: Glucosio Glucosio Piruvato Piruvato Gluconeogenesi La Glicolisi (dal greco glykys dolce e lysis scissione) E` la via metabolica che
TRANSAMINASI (AMINOTRANSFERASI) agiscono con un meccanismo ping-pong. sono presenti sia nel citosol che nei mitocondri
 agiscono con un meccanismo ping-pong. sono presenti sia nel citosol che nei mitocondri") TRANSAMINASI (AMINOTRANSFERASI) sono PLP-dipendenti agiscono con un meccanismo ping-pong catalizzano reazioni reversibili sono presenti sia nel citosol che nei mitocondri Seguiamo le fasi di una reazione
TRANSAMINASI (AMINOTRANSFERASI) sono PLP-dipendenti agiscono con un meccanismo ping-pong catalizzano reazioni reversibili sono presenti sia nel citosol che nei mitocondri Seguiamo le fasi di una reazione
Fosforilazione ossidativa
 Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Fosforilazione ossidativa La fosforilazione ossidativa rappresenta il culmine del metabolismo energetico negli organismi aerobi E costituita da due fenomeni strettamente accoppiati: 1. I coenzimi ridotti
Tessuto Adiposo - metabolismo degli acidi grassi
 Tessuto Adiposo - metabolismo degli acidi grassi Insieme di numerose cellule, ADIPOCITI: AG + Glicerolo TAG 1. Funzione meccanica: protezione contro gli urti e sostegno degli organi interni 2. Funzione
Tessuto Adiposo - metabolismo degli acidi grassi Insieme di numerose cellule, ADIPOCITI: AG + Glicerolo TAG 1. Funzione meccanica: protezione contro gli urti e sostegno degli organi interni 2. Funzione
L energia negli esseri viventi: fotosintesi e respirazione viste dal punto di vista chimico
 Dipartimento di Scienze Chimiche e Geologiche L energia negli esseri viventi: fotosintesi e respirazione viste dal punto di vista chimico Gianantonio Battistuzzi La fotosintesi e la respirazione aerobica
Dipartimento di Scienze Chimiche e Geologiche L energia negli esseri viventi: fotosintesi e respirazione viste dal punto di vista chimico Gianantonio Battistuzzi La fotosintesi e la respirazione aerobica
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)
 Glucosio. GLUCONEOGENESI (sintesi da precursori non glucidici)") DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
DEGRADAZIONE di polisaccaridi (glicogeno epatico, amido o glicogeno dalla dieta) Glucosio GLUCONEOGENESI (sintesi da precursori non glucidici) 1 2 Il glucosio viene trasportato all interno della cellula
FISIOLOGIA dell ALLENAMENTO. dai processi fisiologici alle metodiche di allenamento
 FISIOLOGIA dell ALLENAMENTO dai processi fisiologici alle metodiche di allenamento Introduzione alla fisiologia Metabolismo cellulare Il sistema cardiovascolare: funzione cardiaca vasi, flusso, pressione
FISIOLOGIA dell ALLENAMENTO dai processi fisiologici alle metodiche di allenamento Introduzione alla fisiologia Metabolismo cellulare Il sistema cardiovascolare: funzione cardiaca vasi, flusso, pressione
Le reazioni esoergoniche e quelle endoergoniche del metabolismo cellulare sono legate dalla molecola di ATP.
 1 METABOLISMO CELLULARE Definizione Metabolismo cellulare insieme delle reazioni che producono e necessitano energia nel nostro corpo. Nel metabolismo si distinguono due fasi: Fase catabolica Insieme delle
1 METABOLISMO CELLULARE Definizione Metabolismo cellulare insieme delle reazioni che producono e necessitano energia nel nostro corpo. Nel metabolismo si distinguono due fasi: Fase catabolica Insieme delle
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di
 IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
IL TESSUTO ADIPOSO La funzione del tessuto adiposo, che è costituito da adipociti, è quella di garantire all organismo animale una riserva di energia, prontamente disponibile quando l organismo ne ha bisogno.
Helena Curtis N. Sue Barnes
 Helena Curtis N. Sue Barnes LA FOTOSINTESI INDICE Organismi autotrofi ed eterotrofi Confronto tra fotosintesi e respirazione Reazioni endoergoniche ed esoergoniche Reazioni accoppiate Fase luce-dipendente
Helena Curtis N. Sue Barnes LA FOTOSINTESI INDICE Organismi autotrofi ed eterotrofi Confronto tra fotosintesi e respirazione Reazioni endoergoniche ed esoergoniche Reazioni accoppiate Fase luce-dipendente
Il metabolismo microbico
 Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
Corso di Microbiologia Generale. A.A. 2015-2016 Il metabolismo microbico Dott.ssa Annalisa Serio Il metabolismo Insieme di reazioni chimiche che avvengono all interno di un organismo vivente: Le reazioni
Le modalità di trasporto attraverso la membrana plasmatica
 Le modalità di trasporto attraverso la membrana plasmatica Esperimento sulla diffusione di diversi tipi di composti attraverso un doppio strato lipidico sintetico Le membrane biologiche sono semipermeabili
Le modalità di trasporto attraverso la membrana plasmatica Esperimento sulla diffusione di diversi tipi di composti attraverso un doppio strato lipidico sintetico Le membrane biologiche sono semipermeabili
Lattato. Glicolisi. Piruvato
 Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
Lattato Glicolisi Piruvato REAZIONI MITOCONDRIALI DEL PIRUVATO + Piruvato Deidrogenasi + CO 2 + H 2 O ATP ADP + Pi Piruvato Carbossilasi + 2 H + ossalacetato La Piruvato Carbossilasi è una proteina Mitocondriale
Struttura del Mitocondrio Due membrane
 Mitocondri Mitocondri Mitocondri Organello con doppia membrana 0.5-1.0 micron di larghezza, fino a 10 micron di lunghezza Presente in tutte le cellule Principale centrale energetica della cellula Energia
Mitocondri Mitocondri Mitocondri Organello con doppia membrana 0.5-1.0 micron di larghezza, fino a 10 micron di lunghezza Presente in tutte le cellule Principale centrale energetica della cellula Energia
ERITROCITI STRUTTURA E METABOLISMO
 ERITROCITI STRUTTURA E METABOLISMO Il globulo rosso maturo ha una struttura molto semplice perché sfornito di nucleo e degli organuli citoplasmatici Dott.ssa M. COPPOLA Dott.ssa C. CRISCUOLO LA MEMBRANA
ERITROCITI STRUTTURA E METABOLISMO Il globulo rosso maturo ha una struttura molto semplice perché sfornito di nucleo e degli organuli citoplasmatici Dott.ssa M. COPPOLA Dott.ssa C. CRISCUOLO LA MEMBRANA
IL METABOLISMO ENERGETICO
 IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
IL METABOLISMO ENERGETICO IL METABOLISMO L insieme delle reazioni chimiche che riforniscono la cellula e l organismo di energia e materia SI DIVIDE IN: CATABOLISMO produce ENERGIA METABOLISMO ENERGETICO
NADH e NADPH. ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente)
 della cellula NADH, NADPH = 2 moneta (potere riducente)") NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
NADH e NADPH ATP = 1 moneta (energetica) della cellula NADH, NADPH = 2 moneta (potere riducente) NADH e NADPH non sono funzionalmente intercambiabili. [NAD + ]/[NADH] ~ 1000 favorisce ossidazione [NADP
Principi di Biochimica
 Principi di Biochimica Augusto Innocenti Biologo Nutrizionista Perfezionamento in Biochimica e Biologia Molecolare Phd in Neurobiologia e Neurofisiologia Materia: Atomi e Molecole La materie è costituita
Principi di Biochimica Augusto Innocenti Biologo Nutrizionista Perfezionamento in Biochimica e Biologia Molecolare Phd in Neurobiologia e Neurofisiologia Materia: Atomi e Molecole La materie è costituita
Meccanismi di trasporto dei nutrienti
 CH 2 O CH 2 O Meccanismi di trasporto dei nutrienti Le molecole di nutrienti devono oltrepassare la barriera cellulare costituita sia dalla parete batterica che dalla membrana cellulare che rimane il principale
CH 2 O CH 2 O Meccanismi di trasporto dei nutrienti Le molecole di nutrienti devono oltrepassare la barriera cellulare costituita sia dalla parete batterica che dalla membrana cellulare che rimane il principale