Lezione 5. Cambiamenti evolutivi nelle sequenze nucleotidiche Distanze
|
|
|
- Flaviana Gianni
- 6 anni fa
- Visualizzazioni
Transcript
1 Lezione 5 Cambiamenti evolutivi nelle sequenze nucleotidiche Distanze
2 Graur and Li ch 3 materiale La lezione 5 ci permetterà di capire 1. come possa evolvere una sequenza di nucleotidi 2. quanto due sequenze nulceotidiche sono diverse fra loro 3. mettere questa differenza in relazione con il tempo trascorso dall antenato comune
3 Sostituzioni nucleotidiche AATGAAAGAA tempo ACTGGAGGAA 10 siti; 3 differenze
4 Sostituzioni nucleotidiche Processo base nell evoluzione molecolare Essenziale per comprendere i meccanismi di evoluzione del DNA I cambiamenti delle sequenze nucleotidiche vengono usati per Stimare il tasso di evoluzione Ricostruire la storia evolutiva degli organismi
5 Sostituzioni nucleotidiche AATGAAAGAA Non direttamente osservabili C T tempo ACTGGAGGAA 10 siti; 3 differenze
6 Sostituzioni nucleotidiche: modelli Se vogliamo davvero capire come evolve una sequenza dobbiamo: Stabilire che cosa ci aspettiamo ad un determinato sito al passare del tempo Stabilire i cambiamenti temporali nella probabilità di avere un determinato nucleotide ad un dato sito, considerando possibili «complicazioni» (sostituzioni multiple etc)
7 Sostituzioni nucleotidiche: modelli Modello: descrizione teorica del modo in cui un processo funziona Parametro: un fattore che definisce il sistema e ne determina il comportamento
8 Sostituzioni nucleotidiche: modelli Quanti parametri? Modelli diversi con un diverso numero di parametri Per studiare la dinamica delle sostituzioni dobbiamo fare delle assunzioni sulla probabilità di cambiamento di un nucleotide con un altro
9 Sostituzioni nucleotidiche: modelli Un processo stocastico markoviano è un processo stocastico nel quale la probabilità di transizione che determina il passaggio ad uno stato di sistema dipende unicamente dallo stato di sistema immediatamente precedente (proprietà di Markov) e non dal come si è giunti a tale stato Le probabilità di transizione p(a A), p(a C), p(a T), p(a G) etc modellano la probabilità di transizione da A ad A, C, T e G rispettivamente. La matrice verrà definita come segue: probabilità di transizione tra gli stati della catena Attenzione! Il termine TRNSIZIONE usato in questo contesto indica il CAMBIAMENTO, il PASSAGGIO da un nucleotide ad un altro. Qui non si intende come PUR PUR o PIR PIR come alternativa alla trasversione
10 Sostituzioni nucleotidiche: JC69 Modello di Jukes e Cantor (1969) Assunzioni: Tutti i cambiamenti sono equiprobabili: modello ad UN parametro: un solo tasso di sostituzione (α) Tutti i nucleotidi sono presenti alla stessa frequenza (25%) Esempio di : 10-9 sostituzioni/sito /anno
11 Sostituzioni nucleotidiche: JC69 Qual è la probabilità che il nucleotide A al tempo 0 sia ancora A al tempo 2? Due possibili percorsi: 1. Il nucleotide è rimasto lo stesso dal tempo 0 al tempo Il nucleotide è cambiato in T, C, o G al tempo 1, ma è tornato ad essere A al tempo 2 JC69 permette di correggere per sostituzioni multiple (multiple hits)
12 Sostituzioni nucleotidiche: K80 o K2P Modello di Kimura (1980): Kimura 2 parametri Assunzioni: In questo modello il tasso con cui avvengono le transizioni (α) può essere diverso da quello con cui avvengono le trasversioni (β): 2 parametri β β β β β β β β Tutti i nucleotidi sono presenti alla stessa frequenza (25%)
13 Sostituzioni nucleotidiche: K80 o K2P Qual è la probabilità che il nucleotide A al tempo 0 sia ancora A al tempo 2? Quattro possibili percorsi:
14 Sostituzioni nucleotidiche: K80 o K2P Modello di Kimura (1980): Kimura 2 parametri Assumiamo di nuovo che al tempo 0 il nucleotide sia A, trasversioni transizioni β β β β β β β β C T G
15 Sostituzioni nucleotidiche: altri modelli Assunzioni: Non tutti i nucleotidi sono presenti alla stessa frequenza!! T C A G T C A G
16 Sostituzioni nucleotidiche: altri modelli T C A G T C A G
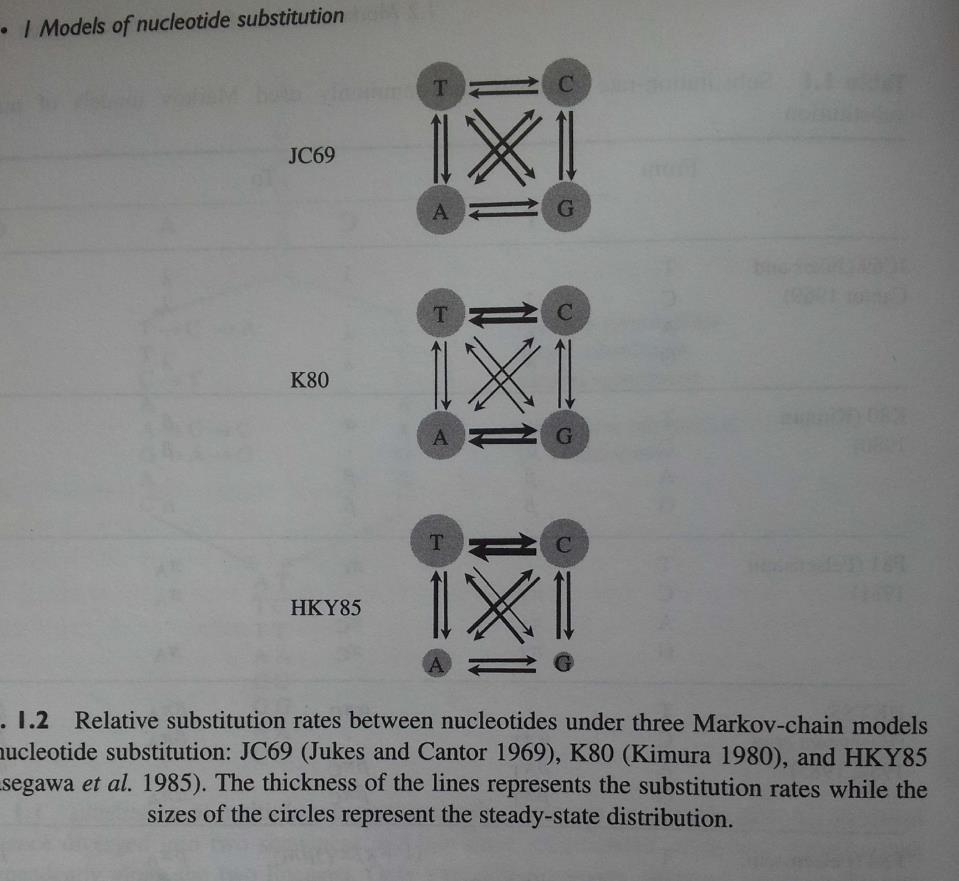
17
18 Sostituzioni nucleotidiche: JC69 Come si può correggere per tutte le possibili sostituzioni multiple contemplando tutti i passaggi nascosti?
19 Sostituzioni nucleotidiche: JC69 Il calcolo della probabilità di transizione considera TUTTI i possibili percorsi evolutivi che potrebbero essere avvenuti i t 1 X= (T, C, A, G) x x x Teorema di Chapman-Kolmogorov: la probabilità di transizione di ogni nucleotide i ad ogni nucleotide j nel tempo t 1 + t 2 è la sommatoria di tutti i possibili stati X ad ogni tempo intermedio t 1 j t 2
20 Distanze fra sequenze Dopo la divergenza fra due sequenze nucleotidiche ognuna inizia ad accumulare sostituzioni Per comparare due sequenze omologhe si stima tempo K = numero di sostituzioni per sito dal momento della divergenza fra due sequenze K

21 Distanze fra sequenze Se il tasso di evoluzione è costante nel tempo la distanza in termini di differenze nucleotidiche tra due sequenze crescerà linearmente con il crescere del tempo di divergenza. AATGAAAGAA ACTGGAGGAA 10 siti; 3 differenze Una semplice misura di distanza è la proporzione dei siti differenti (a volte chiamata distanza p) 10 siti; 3 differenze distanza = 30% = 0.3
22 Sostituzioni Distanze fra nucleotidiche sequenze AATGAAAGAA 10 siti; 3 differenze ACTGGAGGAA distanza = 30% = 0.3 Questa proporzione grezza funziona per sequenze che sono molto vicine evolutivamente. Se è passato molto tempo dalla divergenza, p sottostima il numero di sostituzioni che sono realmente avvenute. Un sito variabile può originarsi attraverso più percorsi e perfino un sito uguale in due sequenze può nascondere retro sostituzioni o sostituzioni parallele. Sostituzioni multiple nascondono alcuni cambiamenti, perciò p non è una funzione diretta del tempo evolutivo. La proporzione grezza p può essere usata solo se p < al 5%
23
24 Distanze fra sequenze Complichiamo lo scenario: correggiamo per multiple hits I modelli di Jukes e Cantor, Kimura, Tamura e Nei etc. possono essere usati oltre che per prevedere l evolversi di una sequenza, anche per valutare la distanza fra due sequenze originatesi da una divergenza
25 Distanze fra sequenze: non coding sites Modello di Jukes e Cantor (1969) K K: numero di sostituzioni per sito dal momento della divergenza p: proporzione osservata di siti differenti tra due sequenze
26 Distanze fra sequenze: non coding sites Modello di Kimura 2 parametri (1980) K: numero di sostituzioni per sito dal momento della divergenza (se P e Q sono uguali si torna all equazione di JC)



27 Distanze fra sequenze: non coding sites Esempio: rrna 12s mtdna Da Yang computational molecular evolution Oxford University Press 2006 Tempo di divergenza: 15.7 Mya (
28 Distanze fra sequenze: non coding sites Esempio: rrna 12s mtdna K2P80 JC69 K N= ( ) + ( ) = 948 p= ( )/948= 90/948= P = transiz = ( )/948 =84/948=0.088 Q= trasv= ( )/948 = 6/948 = JC69 : K = K2P80: K = La differenza è minima Da Yang computational molecular evolution
29 Distanze fra sequenze: non coding sites Aumentiamo la divergenza: N= 948 p= 500/948 = P = transiz = 400/948 = Q= trasv= 100/948 = JC69 : K = 0.91 K2P80: K = 1.55 K2P80 JC69 K La differenza tra le due stime aumenta all aumentare della divergenza Se c è un alto livello di divergenza (diciamo p>5%) e, soprattutto, se ci sono motivi a priori di pensare che il tasso di transizione differisca da quello di trasversione è meglio considerare modelli più complessi di Jukes and Cantor
30 Distanze fra sequenze: coding sites Calcolare il numero di sostituzioni tra due sequenze codificanti proteine è più complesso perché è necessario distinguere tra sostituzioni sinonime e non sinonime
31 Distanze fra sequenze: coding sites K S Seq1 Seq2 Non Sin Ser Thr Glu Met Cys Leu TCA ACT GAG ATG TGT TTA TCG ACA GAG ATA TGT CTA Ser Thr Glu Ile Cys Leu Sin Sin Sin Basta contare? NO: Problemi con il denominatore K A
32 Distanze fra sequenze: coding sites Perché non basta contare? Sinonimo Non sinonimo Per esempio alcuni siti non sono solo sinonimi o solo non sinonimi, dipende da come mutano

33 Distanze fra sequenze: coding sites Seq1 Seq2 Non Sin Ser Thr Glu Met Cys Leu TCA ACT GAG ATG TGT TTA TCG ACA GAG ATA TGT CTA Ser Thr Glu Ile Cys Leu Sin Sin Sin Basta contare? NO: Problemi con il numeratore
34 Distanze fra sequenze: coding sites Problemi col numeratore: Esempio: quando due codoni omologhi differiscono per due o più sostituzioni l ordine delle sostituzioni deve essere conosciuto per classificare il sito come sinonimo o non sinonimo. Esempio: CCC nella sequenza 1 e CAA nella sequenza 2 La classificazione dei siti dipende dall ordine in cui le sostituzioni sono avvenute Percorso I: Percorso II: CCC (Pro) CCA (Pro) CAA (Gln) 1 sinonimo e 1 non sinonimo CCC (Pro) CAC (His) CAA (Gln) 2 non sinonimi
35 Distanze fra sequenze: coding sites Basta contare? NO: possibili soluzioni Metodi di Miyata & Yasunaga (1980) e Nei & Gojobori (1986) Nei and Gojobori calculate average number of synonymous and nonsynonymous sites allowing particular sites to be a portion in each category (can also weight substitution pathway probabilities).
36 Distanze fra sequenze: coding sites Cambiamenti syn M S (o nonsyn M A ) Tot basi syn N S (o nonsyn N A ) Basta contare? NO: possibili soluzioni denominatore 1. Consideriamo una posizione specifica in un codon. Se i è il numero di possibili cambiamenti sinonimi a quel sito allora lo conteremo come i/3 sinonimo e (3 i)/3 non sinonimo. 2. Contiamo il numero di siti sinonimi e non sinonimi in ogni sequenza e calcoliamo la media tra le due sequenze. Il numero medio si siti sinonimi è N S e quello di non sinonimi è N A.
37 Distanze fra sequenze: coding sites Metodi di Miyata & Yasunaga (1980) e Nei & Gojobori (1986) per più di una differenza: considerare i diversi percorsi in che ordine sono avvenute le mutazioni? Differenze syn M S (o nonsyn M A ) Tot basi syn N S (o nonsyn N A ) 3. Classifichiamo le differenze in sinonime e non sinonime per due codon con 1 differenza è semplice Val GTC GTT > sinonimo Val GTC GCC > non sinonimo Val Ala numeratore Percorso I: CCC (Pro) CCA (Pro) CAA (Gln) 1 sinonimo e 1 non sinonimo Percorso II: CCC (Pro) CAC (His) CAA (Gln) 2 non sinonimi Approccio non pesato: Tutto è equiprobabile Nei and Gojobori Ma=differenze non sin: (1+2)/2 = 1.5 Ms=differenze sinonime: (1+0)/2 = 0.5
38 Distanze fra sequenze: coding sites Differenze syn M S (o nonsyn M A ) Tot basi syn N S (o nonsyn N A ) Metodi di Miyata & Yasunaga (1980) e Nei & Gojobori (1986) 4. Il numero di mutazioni sinonime per sito sinonimo p S = M S / N S Il numero di mutazioni non sinonime per sito non sinonimo p A = M A / N A Ma ricordate il problema delle multiple hits? > Usiamo Jukes e Cantor (o altro) per correggere
39 Distanze fra sequenze: coding sites Nei & Gojobori (1986)

40 Distanze fra sequenze: coding sites Nr Siti S Nr siti A Ka Ks Ka/Ks
Le L z e io i ne n 6 Co C n o f n ro r n o t n i i fra r a se s q e u q e u n e z n e z : e di d s i t s a t nz n e z, e allineamenti
 Lezione 6 Confronti fra sequenze: distanze, Confronti fra sequenze: distanze, allineamenti Distanze fra sequenze Per N siti ed n differenze: grado di divergenza = n/n AATGAAAGAA 10 siti; 3 differenze ACTGGAGGAA
Lezione 6 Confronti fra sequenze: distanze, Confronti fra sequenze: distanze, allineamenti Distanze fra sequenze Per N siti ed n differenze: grado di divergenza = n/n AATGAAAGAA 10 siti; 3 differenze ACTGGAGGAA
Lezione 8. Selezione positiva o darwiniana
 Lezione 8 Selezione positiva o darwiniana Graur and Li: Capitolo 4 (+Cap 2 p63-65) Graur: lecture 20 Ziheng Yang: computational molecular evolution In generale il tasso di sostituzione in geni e regioni
Lezione 8 Selezione positiva o darwiniana Graur and Li: Capitolo 4 (+Cap 2 p63-65) Graur: lecture 20 Ziheng Yang: computational molecular evolution In generale il tasso di sostituzione in geni e regioni
Lezione 1. Le molecole di base che costituiscono la vita
 Lezione 1 Le molecole di base che costituiscono la vita Le molecole dell ereditarietà 5 3 L informazione ereditaria di tutti gli organismi viventi, con l eccezione di alcuni virus, è a carico della molecola
Lezione 1 Le molecole di base che costituiscono la vita Le molecole dell ereditarietà 5 3 L informazione ereditaria di tutti gli organismi viventi, con l eccezione di alcuni virus, è a carico della molecola
Metodi di Distanza. G.Allegrucci riproduzione vietata
 Metodi di Distanza La misura più semplice della distanza tra due sequenze nucleotidiche è contare il numero di siti nucleotidici che differiscono tra le due sequenze Quando confrontiamo siti omologhi in
Metodi di Distanza La misura più semplice della distanza tra due sequenze nucleotidiche è contare il numero di siti nucleotidici che differiscono tra le due sequenze Quando confrontiamo siti omologhi in
Biologia Molecolare. CDLM in CTF La riparazione del DNA
 Biologia Molecolare CDLM in CTF 2010-2011 La riparazione del DNA I tipi di mutazione e le conseguenze Le classi di danno al DNA Meccanismi di riparazione La necessità di codificare l informazione L informazione
Biologia Molecolare CDLM in CTF 2010-2011 La riparazione del DNA I tipi di mutazione e le conseguenze Le classi di danno al DNA Meccanismi di riparazione La necessità di codificare l informazione L informazione
TRADUZIONE. 2. Transfer (legato agli aminoacidi) 3. Ribosomale (associato a proteine nei ribosomi)
 3. Ribosomale (associato a proteine nei ribosomi)") enhancer promotore regione trascritta TATA trascrizione 5 3 splicing mrna 5 3 traduzione Proteina NH2 COOH Funzione biologica TRADUZIONE I tre ruoli svolti dall RNA: 1. Messaggero 2. Transfer (legato agli
enhancer promotore regione trascritta TATA trascrizione 5 3 splicing mrna 5 3 traduzione Proteina NH2 COOH Funzione biologica TRADUZIONE I tre ruoli svolti dall RNA: 1. Messaggero 2. Transfer (legato agli
Maximum Likelihood. Giuliana Allegrucci - Riproduzione vietata
 Maximum Likelihood Potrebbe essere chiamato massima probabilità. E il metodo più nuovo E stato reso popolare da Joseph Felsenstein, Seattle, Washington. Il suo lento apparire nella comunità scientifica
Maximum Likelihood Potrebbe essere chiamato massima probabilità. E il metodo più nuovo E stato reso popolare da Joseph Felsenstein, Seattle, Washington. Il suo lento apparire nella comunità scientifica
Filogenesi molecolare
 Filogenesi molecolare Geni ortologhi e geni paraloghi Geni ortologhi: geni simili riscontrabili in organismi correlati tra loro. Il fenomeno della speciazione porta alla divergenza dei geni e quindi delle
Filogenesi molecolare Geni ortologhi e geni paraloghi Geni ortologhi: geni simili riscontrabili in organismi correlati tra loro. Il fenomeno della speciazione porta alla divergenza dei geni e quindi delle
COME CALCOLARE IL PUNTEGGIO DI UN ALLINEAMENTO? Il problema del calcolo del punteggio di un allineamento può essere considerato in due modi diversi
 COME CALCOLARE IL PUNTEGGIO DI UN ALLINEAMENTO? Il problema del calcolo del punteggio di un allineamento può essere considerato in due modi diversi che, però, sono le due facce di una stessa medaglia al
COME CALCOLARE IL PUNTEGGIO DI UN ALLINEAMENTO? Il problema del calcolo del punteggio di un allineamento può essere considerato in due modi diversi che, però, sono le due facce di una stessa medaglia al
RETI DI TELECOMUNICAZIONE
 RETI DI TELECOMUNICAZIONE CATENE DI MARKOV TEMPO CONTINUE Definizioni Sia dato un processo stocastico x(t) che può assumere valori discreti appartenenti ad un insieme se accade che il processo è una catena
RETI DI TELECOMUNICAZIONE CATENE DI MARKOV TEMPO CONTINUE Definizioni Sia dato un processo stocastico x(t) che può assumere valori discreti appartenenti ad un insieme se accade che il processo è una catena
EVOLUZIONE MOLECOLARE. Silvia Fuselli
 EVOLUZIONE MOLECOLARE Silvia Fuselli silvia.fuselli@unife.it TESTI Organizzazione del corso Graur and Li, Fundamentals of molecular evolution, Sinauer 2000 Michael Lynch, The Origins of Genome Architecture,
EVOLUZIONE MOLECOLARE Silvia Fuselli silvia.fuselli@unife.it TESTI Organizzazione del corso Graur and Li, Fundamentals of molecular evolution, Sinauer 2000 Michael Lynch, The Origins of Genome Architecture,
Le L z e io i ne n 7 Va V r a ia i z a io i n o e n e ne n i e i ta t s a s s i s i di d i so s s o t s it i uz u io i ne e nucleotidica
 Lezione 7 Variazione nei tassi di sostituzione Variazione nei tassi di sostituzione nucleotidica Graurand Li: Capitolo 4 Graur: lecture19 Origine comune 5 milioni diannifa 1500 milioni diannifa Le scale
Lezione 7 Variazione nei tassi di sostituzione Variazione nei tassi di sostituzione nucleotidica Graurand Li: Capitolo 4 Graur: lecture19 Origine comune 5 milioni diannifa 1500 milioni diannifa Le scale
Metodo della matrice a punti
 Metodo della matrice a punti proposto da Gibbs and McIntyre (1970) consente di evidenziare ripetizioni dirette o inverse nelle sequenze prevedere regioni complementari nell RNA che possano potenzialmente
Metodo della matrice a punti proposto da Gibbs and McIntyre (1970) consente di evidenziare ripetizioni dirette o inverse nelle sequenze prevedere regioni complementari nell RNA che possano potenzialmente
Struttura secondaria
 Struttura secondaria Struttura localmente ordinata Polimeri lineari ad unità monomeriche asimmetriche elica j = 0,1,2,..., N N = numero di residui z j = h z j + z 0 x j = r cos (j2p h z /P + d 0 ) y j
Struttura secondaria Struttura localmente ordinata Polimeri lineari ad unità monomeriche asimmetriche elica j = 0,1,2,..., N N = numero di residui z j = h z j + z 0 x j = r cos (j2p h z /P + d 0 ) y j
Markov Chains and Markov Chain Monte Carlo (MCMC)
") Markov Chains and Markov Chain Monte Carlo (MCMC) Alberto Garfagnini Università degli studi di Padova December 11, 2013 Catene di Markov Discrete dato un valore x t del sistema ad un istante di tempo fissato,
Markov Chains and Markov Chain Monte Carlo (MCMC) Alberto Garfagnini Università degli studi di Padova December 11, 2013 Catene di Markov Discrete dato un valore x t del sistema ad un istante di tempo fissato,
Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica. Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA.
 Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA. In genere si ottengono trattando il DNA con agenti chimici (es.
Mutagenesi: introduzione di alterazioni in una sequenza nucleotidica Mutagenesi random: le mutazioni avvengono a caso su un tratto di DNA. In genere si ottengono trattando il DNA con agenti chimici (es.
A W T V A S A V R T S I A Y T V A A A V R T S I A Y T V A A A V L T S I
 COME CALCOLARE IL PUNTEIO DI UN ALLINEAMENTO? Il problema del calcolo del punteggio di un allineamento può essere considerato in due modi diversi che, però, sono le due facce di una stessa medaglia al
COME CALCOLARE IL PUNTEIO DI UN ALLINEAMENTO? Il problema del calcolo del punteggio di un allineamento può essere considerato in due modi diversi che, però, sono le due facce di una stessa medaglia al
UNIVERSITÀ di ROMA TOR VERGATA
 UNIVERSITÀ di ROMA TOR VERGATA Corso di PS-Probabilità P.Baldi Tutorato 9, 19 maggio 11 Corso di Laurea in Matematica Esercizio 1 a) Volendo modellizzare l evoluzione della disoccupazione in un certo ambito
UNIVERSITÀ di ROMA TOR VERGATA Corso di PS-Probabilità P.Baldi Tutorato 9, 19 maggio 11 Corso di Laurea in Matematica Esercizio 1 a) Volendo modellizzare l evoluzione della disoccupazione in un certo ambito
LE AUGURO BUON LAVORO!
 Nome:. Cognome:...... Pt./137 Nota ESAME DI SCIENZE SPERIMENTALI: BIOLOGIA Indicazioni: 1. Risponda sempre negli spazi indicati; non separi né scriva sul retro dei fogli. 2. Scriva, o, laddove richiesto,
Nome:. Cognome:...... Pt./137 Nota ESAME DI SCIENZE SPERIMENTALI: BIOLOGIA Indicazioni: 1. Risponda sempre negli spazi indicati; non separi né scriva sul retro dei fogli. 2. Scriva, o, laddove richiesto,
Corso di Bioinformatica. Docente: Dr. Antinisca DI MARCO
 Corso di Bioinformatica Docente: Dr. Antinisca DI MARCO Email: antinisca.dimarco@univaq.it Analisi Filogenetica Gene Ancestrale duplicazione genica La filogenesi è lo studio delle relazioni evolutive tra
Corso di Bioinformatica Docente: Dr. Antinisca DI MARCO Email: antinisca.dimarco@univaq.it Analisi Filogenetica Gene Ancestrale duplicazione genica La filogenesi è lo studio delle relazioni evolutive tra
8. Energia e lavoro. 2 Teorema dell energia per un moto uniformemente
 1 Definizione di lavoro 8. Energia e lavoro Consideriamo una forza applicata ad un corpo di massa m. Per semplicità ci limitiamo, inizialmente ad una forza costante, come ad esempio la gravità alla superficie
1 Definizione di lavoro 8. Energia e lavoro Consideriamo una forza applicata ad un corpo di massa m. Per semplicità ci limitiamo, inizialmente ad una forza costante, come ad esempio la gravità alla superficie
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita
 -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita") MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
LEZIONE N. 11 ( a cura di MADDALENA BEI)
") LEZIONE N. 11 ( a cura di MADDALENA BEI) F- test Assumiamo l ipotesi nulla H 0 :β 1,...,Β k =0 E diverso dal verificare che H 0 :B J =0 In realtà F - test è più generale H 0 :Aβ=0 H 1 :Aβ 0 A è una matrice
LEZIONE N. 11 ( a cura di MADDALENA BEI) F- test Assumiamo l ipotesi nulla H 0 :β 1,...,Β k =0 E diverso dal verificare che H 0 :B J =0 In realtà F - test è più generale H 0 :Aβ=0 H 1 :Aβ 0 A è una matrice
Organizzazione del genoma umano
 Organizzazione del genoma umano Famiglie di geni o geniche Copie multiple di geni, tutte con sequenza identica o simile. La famiglia multigenica corrisponde a un insieme di geni correlati che si sono evoluti
Organizzazione del genoma umano Famiglie di geni o geniche Copie multiple di geni, tutte con sequenza identica o simile. La famiglia multigenica corrisponde a un insieme di geni correlati che si sono evoluti
REPLICAZIONE DEL DNA
 REPLICAZIONE DEL DNA La replicazione (o anche duplicazione) è il meccanismo molecolare attraverso cui il DNA produce una copia di sé stesso. Ogni volta che una cellula si divide, infatti, l'intero genoma
REPLICAZIONE DEL DNA La replicazione (o anche duplicazione) è il meccanismo molecolare attraverso cui il DNA produce una copia di sé stesso. Ogni volta che una cellula si divide, infatti, l'intero genoma
Analisi della varianza a una via
 Analisi della varianza a una via Statistica descrittiva e Analisi multivariata Prof. Giulio Vidotto PSY-NET: Corso di laurea online in Discipline della ricerca psicologico-sociale SOMMARIO Modelli statistici
Analisi della varianza a una via Statistica descrittiva e Analisi multivariata Prof. Giulio Vidotto PSY-NET: Corso di laurea online in Discipline della ricerca psicologico-sociale SOMMARIO Modelli statistici
ESERCIZI SUI PUNTI DI NON DERIVABILITÀ TRATTI DA TEMI D ESAME
 ESERCIZI SUI PUNTI DI NON DERIVABILITÀ TRATTI DA TEMI D ESAME a cura di Michele Scaglia FUNZIONI DERIVABILI Sia f : domf R una funzione e sia 0 domf di accumulazione per domf Chiamiamo derivata prima di
ESERCIZI SUI PUNTI DI NON DERIVABILITÀ TRATTI DA TEMI D ESAME a cura di Michele Scaglia FUNZIONI DERIVABILI Sia f : domf R una funzione e sia 0 domf di accumulazione per domf Chiamiamo derivata prima di
19 LIMITI FONDAMENTALI - II
 19 LIMITI FONDAMENTALI - II 3. Il ite che permette il calcolo di forme indeterminate in cui sono presenti funzioni logaritmiche è: log1 + = 1. La dimostrazione di questo ite si ha subito dal ite Esempio.
19 LIMITI FONDAMENTALI - II 3. Il ite che permette il calcolo di forme indeterminate in cui sono presenti funzioni logaritmiche è: log1 + = 1. La dimostrazione di questo ite si ha subito dal ite Esempio.
Classificazione. I complessi. Le pietre miliari della tassonomia. Tassonomia del genere Mycobacterium. Pietre miliari nella tassonomia dei micobatteri
 Le pietre miliari della tassonomia Tassonomia del genere Mycobacterium Enrico Tortoli Centro Regionale di Riferimento per i Micobatteri Firenze Adamo è autorizzato da Dio a dare un nome a tutti gli esseri
Le pietre miliari della tassonomia Tassonomia del genere Mycobacterium Enrico Tortoli Centro Regionale di Riferimento per i Micobatteri Firenze Adamo è autorizzato da Dio a dare un nome a tutti gli esseri
La sintesi delle proteine
 La sintesi delle proteine Struttura del trna In che modo l informazione contenuta sotto forma di sequenze nucleotidiche nel DNA e nell RNA si traduce nella sequenza amminoacidica delle proteine? Esperimenti
La sintesi delle proteine Struttura del trna In che modo l informazione contenuta sotto forma di sequenze nucleotidiche nel DNA e nell RNA si traduce nella sequenza amminoacidica delle proteine? Esperimenti
DNA E PROTEINE IL DNA E RACCHIUSO NEL NUCLEO, MENTRE LA SINTESI PROTEICA SI SVOLGE NEL CITOPLASMA: COME VIENE TRASPORTATA L INFORMAZIONE?
 DNA E PROTEINE NUMEROSI DATI SUGGERISCONO CHE IL DNA SVOLGA IL SUO RUOLO GENETICO CONTROLLANDO LA SINTESI DELLE PROTEINE, IN PARTICOLARE DETERMINANDONE LA SEQUENZA IN AMINOACIDI E NECESSARIO RISPONDERE
DNA E PROTEINE NUMEROSI DATI SUGGERISCONO CHE IL DNA SVOLGA IL SUO RUOLO GENETICO CONTROLLANDO LA SINTESI DELLE PROTEINE, IN PARTICOLARE DETERMINANDONE LA SEQUENZA IN AMINOACIDI E NECESSARIO RISPONDERE
IPOTESI UN GENE-UN ENZIMA
 IPOTESI UN GENE-UN ENZIMA DNA: contiene tutte le informazioni per definire lo sviluppo e la fisiologia della cellula: ma come svolge questa funzione? Beadle e Tatum (1941): studiando mutanti della comune
IPOTESI UN GENE-UN ENZIMA DNA: contiene tutte le informazioni per definire lo sviluppo e la fisiologia della cellula: ma come svolge questa funzione? Beadle e Tatum (1941): studiando mutanti della comune
Contenuti: Capitolo 14 del libro di testo
 Test d Ipotesi / TIPICI PROBLEMI DI VERIFICA DI IPOTESI SONO Test per la media Test per una proporzione Test per la varianza Test per due campioni indipendenti Test di indipendenza Contenuti Capitolo 4
Test d Ipotesi / TIPICI PROBLEMI DI VERIFICA DI IPOTESI SONO Test per la media Test per una proporzione Test per la varianza Test per due campioni indipendenti Test di indipendenza Contenuti Capitolo 4
Riconoscimento e recupero dell informazione per bioinformatica
 Riconoscimento e recupero dell informazione per bioinformatica Filogenesi Manuele Bicego Corso di Laurea in Bioinformatica Dipartimento di Informatica - Università di Verona Sommario Introduzione alla
Riconoscimento e recupero dell informazione per bioinformatica Filogenesi Manuele Bicego Corso di Laurea in Bioinformatica Dipartimento di Informatica - Università di Verona Sommario Introduzione alla
Esame di Sistemi ad Eventi Discreti
 Esame di Sistemi ad Eventi Discreti - 16.12.2008 Esercizio 1 (esame completo/recupero prima parte) Un macchinario è programmato per task. Ciascun task è composto da subtask, che possono essere di tipo
Esame di Sistemi ad Eventi Discreti - 16.12.2008 Esercizio 1 (esame completo/recupero prima parte) Un macchinario è programmato per task. Ciascun task è composto da subtask, che possono essere di tipo
Lezione 6. Lo string matching
 Lezione 6 Lo string matching String matching Date due stringhe (sequenze di caratteri) vogliamo stabilire se sono uguali Nel caso dello string matching, due stringhe sono uguali se... sono uguali ( DNA
Lezione 6 Lo string matching String matching Date due stringhe (sequenze di caratteri) vogliamo stabilire se sono uguali Nel caso dello string matching, due stringhe sono uguali se... sono uguali ( DNA
Caratteristiche generali
 AMMINOACIDI Gli amminoacidi sono le unità costruttive (building blocks) delle proteine. Come dice il termine, gli amminoacidi naturali sono costituiti da un gruppo amminico (-NH 2 ) e da un gruppo carbossilico
AMMINOACIDI Gli amminoacidi sono le unità costruttive (building blocks) delle proteine. Come dice il termine, gli amminoacidi naturali sono costituiti da un gruppo amminico (-NH 2 ) e da un gruppo carbossilico
lezione 8 AA Paolo Brunori
 AA 2016-2017 Paolo Brunori regressione multipla con n = k Immaginate di voler studiare i determinanti del voto all esame di econometria Y = β 1 X 1 + u Y i = β 1 H i + u i H=ore studiate alla settimana
AA 2016-2017 Paolo Brunori regressione multipla con n = k Immaginate di voler studiare i determinanti del voto all esame di econometria Y = β 1 X 1 + u Y i = β 1 H i + u i H=ore studiate alla settimana
Esame di Statistica del 19 settembre 2006 (Corso di Laurea Triennale in Biotecnologie, Università degli Studi di Padova).
.") Esame di Statistica del 19 settembre 2006 (Corso di Laurea Triennale in Biotecnologie, Università degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione: si
Esame di Statistica del 19 settembre 2006 (Corso di Laurea Triennale in Biotecnologie, Università degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione: si
Note sulle Catene di Markov
 Note sulle Catene di Markov ELAUT Prof. Giuseppe C. Calafiore Sommario Queste note contengono un estratto schematico ridotto di parte del materiale relativo alle Catene di Markov a tempo continuo e a tempo
Note sulle Catene di Markov ELAUT Prof. Giuseppe C. Calafiore Sommario Queste note contengono un estratto schematico ridotto di parte del materiale relativo alle Catene di Markov a tempo continuo e a tempo
Metodi statistici per l economia (Prof. Capitanio) Slide n. 10. Materiale di supporto per le lezioni. Non sostituisce il libro di testo
 Slide n. 10. Materiale di supporto per le lezioni. Non sostituisce il libro di testo") Metodi statistici per l economia (Prof. Capitanio) Slide n. 10 Materiale di supporto per le lezioni. Non sostituisce il libro di testo 1 REGRESSIONE LINEARE Date due variabili quantitative, X e Y, si è
Metodi statistici per l economia (Prof. Capitanio) Slide n. 10 Materiale di supporto per le lezioni. Non sostituisce il libro di testo 1 REGRESSIONE LINEARE Date due variabili quantitative, X e Y, si è
5. Applicazione ai dati sperimentali, un modello di previsione delle temperature
 5. Applicazione ai dati sperimentali, un modello di previsione delle temperature 5.1 Ricostruzione dello spazio delle fasi L utilizzo del teorema di embedding per ricostruire lo spazio delle fasi relativo
5. Applicazione ai dati sperimentali, un modello di previsione delle temperature 5.1 Ricostruzione dello spazio delle fasi L utilizzo del teorema di embedding per ricostruire lo spazio delle fasi relativo
ESERCIZI sui VETTORI
 ESERCIZI sui VETTORI 1. Calcolare la somma di v 1 (2, 3) e v 2 (1, 4). 2. Calcolare la somma di v 1 (1, 5, 4) e v 2 (6, 8, 2). 3. Calcolare il prodotto di α = 2 e v 1 (1, 4). 4. Calcolare il prodotto di
ESERCIZI sui VETTORI 1. Calcolare la somma di v 1 (2, 3) e v 2 (1, 4). 2. Calcolare la somma di v 1 (1, 5, 4) e v 2 (6, 8, 2). 3. Calcolare il prodotto di α = 2 e v 1 (1, 4). 4. Calcolare il prodotto di
Statistica Applicata all edilizia: il modello di regressione
 Statistica Applicata all edilizia: il modello di regressione E-mail: orietta.nicolis@unibg.it 27 aprile 2009 Indice Il modello di Regressione Lineare 1 Il modello di Regressione Lineare Analisi di regressione
Statistica Applicata all edilizia: il modello di regressione E-mail: orietta.nicolis@unibg.it 27 aprile 2009 Indice Il modello di Regressione Lineare 1 Il modello di Regressione Lineare Analisi di regressione
Teoria e tecniche dei test
 Teoria e tecniche dei test Lezione 9 LA STANDARDIZZAZIONE DEI TEST. IL PROCESSO DI TARATURA: IL CAMPIONAMENTO. Costruire delle norme di riferimento per un test comporta delle ipotesi di fondo che è necessario
Teoria e tecniche dei test Lezione 9 LA STANDARDIZZAZIONE DEI TEST. IL PROCESSO DI TARATURA: IL CAMPIONAMENTO. Costruire delle norme di riferimento per un test comporta delle ipotesi di fondo che è necessario
STATISTICA (2) ESERCITAZIONE Dott.ssa Antonella Costanzo
 ESERCITAZIONE Dott.ssa Antonella Costanzo") STATISTICA (2) ESERCITAZIONE 7 11.03.2014 Dott.ssa Antonella Costanzo Esercizio 1. Test di indipendenza tra mutabili In un indagine vengono rilevate le informazioni su settore produttivo (Y) e genere (X)
STATISTICA (2) ESERCITAZIONE 7 11.03.2014 Dott.ssa Antonella Costanzo Esercizio 1. Test di indipendenza tra mutabili In un indagine vengono rilevate le informazioni su settore produttivo (Y) e genere (X)
B =3 0 1=3 0 1=3
 Corsi di Probabilità, Statistica e Processi stocastici per Ing. dell Automazione, Informatica e Inf.Gest.Azienda, a.a. / // Esercizio. Un PC comprato da alcuni mesi, all accensione compie alcune operazioni
Corsi di Probabilità, Statistica e Processi stocastici per Ing. dell Automazione, Informatica e Inf.Gest.Azienda, a.a. / // Esercizio. Un PC comprato da alcuni mesi, all accensione compie alcune operazioni
Genomica Evoluzione e cambiamenti dei genomi. Dott.ssa Inga Prokopenko
 Genomica Evoluzione e cambiamenti dei genomi Dott.ssa Inga Prokopenko Sistematica in biologia La varietà degli organismi viventi richiede organizzaione delle nostre osservazioni Tassonomia in biologia
Genomica Evoluzione e cambiamenti dei genomi Dott.ssa Inga Prokopenko Sistematica in biologia La varietà degli organismi viventi richiede organizzaione delle nostre osservazioni Tassonomia in biologia
LEZIONE 9. Figura 9.1.1
 LEZIONE 9 9.1. Equazioni cartesiane di piani. Abbiamo visto come rappresentare parametricamente un piano. Un altro interessante metodo di rappresentazione di un piano nello spazio è tramite la sua equazione
LEZIONE 9 9.1. Equazioni cartesiane di piani. Abbiamo visto come rappresentare parametricamente un piano. Un altro interessante metodo di rappresentazione di un piano nello spazio è tramite la sua equazione
Metodi Stocastici per la Finanza
 Metodi Stocastici per la Finanza Tiziano Vargiolu vargiolu@math.unipd.it 1 1 Università degli Studi di Padova Anno Accademico 2013-2014 Lezione 4 Indice 1 Convergenza in legge di processi stocastici 2
Metodi Stocastici per la Finanza Tiziano Vargiolu vargiolu@math.unipd.it 1 1 Università degli Studi di Padova Anno Accademico 2013-2014 Lezione 4 Indice 1 Convergenza in legge di processi stocastici 2
TEORIA DELL INFORMAZIONE ED ENTROPIA FEDERICO MARINI
 TEORIA DELL INFORMAZIONE ED ENTROPIA DI FEDERICO MARINI 1 OBIETTIVO DELLA TEORIA DELL INFORMAZIONE Dato un messaggio prodotto da una sorgente, l OBIETTIVO è capire come si deve rappresentare tale messaggio
TEORIA DELL INFORMAZIONE ED ENTROPIA DI FEDERICO MARINI 1 OBIETTIVO DELLA TEORIA DELL INFORMAZIONE Dato un messaggio prodotto da una sorgente, l OBIETTIVO è capire come si deve rappresentare tale messaggio
Esercitazione. 24 Aprile 2012
 Esercitazione 24 Aprile 2012 Il modello di regressione logistica viene utilizzato quando si è interessati a studiare o analizzare la relazione causale tra una variabile dipendente dicotomica e una o più
Esercitazione 24 Aprile 2012 Il modello di regressione logistica viene utilizzato quando si è interessati a studiare o analizzare la relazione causale tra una variabile dipendente dicotomica e una o più
GENETICA E PROBABILITA
 GENETICA E PROBABILITA Cromosoma:corpo cellulare contenete i geni disposti in ordine lineare, appare nel corso della mitosi (o della meiosi) come un filamento o un bastoncello di cromatina Gene:Unità di
GENETICA E PROBABILITA Cromosoma:corpo cellulare contenete i geni disposti in ordine lineare, appare nel corso della mitosi (o della meiosi) come un filamento o un bastoncello di cromatina Gene:Unità di
Mutazioni. Un cambiamento nel materiale genetico che non venga riparato dai meccanismi di riparo costituisce una mutazione
 Mutazioni Un cambiamento nel materiale genetico che non venga riparato dai meccanismi di riparo costituisce una mutazione Le mutazioni possono essere spontanee oppure causate da agenti fisici, chimici
Mutazioni Un cambiamento nel materiale genetico che non venga riparato dai meccanismi di riparo costituisce una mutazione Le mutazioni possono essere spontanee oppure causate da agenti fisici, chimici
Naïve Bayesian Classification
 Naïve Bayesian Classification Di Alessandro rezzani Sommario Naïve Bayesian Classification (o classificazione Bayesiana)... 1 L algoritmo... 2 Naive Bayes in R... 5 Esempio 1... 5 Esempio 2... 5 L algoritmo
Naïve Bayesian Classification Di Alessandro rezzani Sommario Naïve Bayesian Classification (o classificazione Bayesiana)... 1 L algoritmo... 2 Naive Bayes in R... 5 Esempio 1... 5 Esempio 2... 5 L algoritmo
GENOMA. Analisi di sequenze -- Analisi di espressione -- Funzione delle proteine CONTENUTO FUNZIONE. Progetti genoma in centinaia di organismi
 GENOMA EVOLUZIONE CONTENUTO FUNZIONE STRUTTURA Analisi di sequenze -- Analisi di espressione -- Funzione delle proteine Progetti genoma in centinaia di organismi Importante la sintenia tra i genomi The
GENOMA EVOLUZIONE CONTENUTO FUNZIONE STRUTTURA Analisi di sequenze -- Analisi di espressione -- Funzione delle proteine Progetti genoma in centinaia di organismi Importante la sintenia tra i genomi The
Modelli a effetti misti
 Probabilità e Statistica per l analisi di dati sperimentali Modelli a effetti misti Sviluppo e gestione di Data Center per il calcolo scientifico ad alte prestazioni Master Progetto PRISMA, UniBA/INFN
Probabilità e Statistica per l analisi di dati sperimentali Modelli a effetti misti Sviluppo e gestione di Data Center per il calcolo scientifico ad alte prestazioni Master Progetto PRISMA, UniBA/INFN
lezione n. 6 (a cura di Gaia Montanucci) Verosimiglianza: L = = =. Parte dipendente da β 0 e β 1
 Verosimiglianza: L = = =. Parte dipendente da β 0 e β 1") lezione n. 6 (a cura di Gaia Montanucci) METODO MASSIMA VEROSIMIGLIANZA PER STIMARE β 0 E β 1 Distribuzione sui termini di errore ε i ε i ~ N (0, σ 2 ) ne consegue : ogni y i ha ancora distribuzione normale,
lezione n. 6 (a cura di Gaia Montanucci) METODO MASSIMA VEROSIMIGLIANZA PER STIMARE β 0 E β 1 Distribuzione sui termini di errore ε i ε i ~ N (0, σ 2 ) ne consegue : ogni y i ha ancora distribuzione normale,
MATEMATICA E STATISTICA CORSO A CORSO DI LAUREA IN SCIENZE BIOLOGICHE MOLECOLARI
 MATEMATICA E STATISTICA CORSO A CORSO DI LAUREA IN SCIENZE BIOLOGICHE MOLECOLARI ESERCITATI CON ME! I ESERCITAZIONE 1) Misure ripetute (materiale secco su vetrino) della lunghezza del diametro maggiore
MATEMATICA E STATISTICA CORSO A CORSO DI LAUREA IN SCIENZE BIOLOGICHE MOLECOLARI ESERCITATI CON ME! I ESERCITAZIONE 1) Misure ripetute (materiale secco su vetrino) della lunghezza del diametro maggiore
 LEZIONI ED ESERCITAZIONI DI FISICA Prof. Francesco Marchi 1 Esercitazione su: angoli, funzioni e formule goniometriche Indice 1 Goniometriche 1.1 Introduzione.............................. 1. La soluzione
LEZIONI ED ESERCITAZIONI DI FISICA Prof. Francesco Marchi 1 Esercitazione su: angoli, funzioni e formule goniometriche Indice 1 Goniometriche 1.1 Introduzione.............................. 1. La soluzione
Convergenza non condizionata
 Economia Internazionale Economia dello Sviluppo Lezione 5 La convergenza nelle dinamiche di crescita A.A 2007-2008 Stefano Usai Convergenza non L ipotesi di convergenza non e basata sull assunzione che
Economia Internazionale Economia dello Sviluppo Lezione 5 La convergenza nelle dinamiche di crescita A.A 2007-2008 Stefano Usai Convergenza non L ipotesi di convergenza non e basata sull assunzione che
IL CONFRONTO TRA LE VARIANZE DI DUE POPOLAZIONI
 IL CONFRONTO TRA LE VARIANZE DI DUE POPOLAZIONI Perchè confrontare le varianze stimate in due campioni? Torniamo all'esempio dei frinosomi Per poter applicare il test t avevamo detto che le varianze, e
IL CONFRONTO TRA LE VARIANZE DI DUE POPOLAZIONI Perchè confrontare le varianze stimate in due campioni? Torniamo all'esempio dei frinosomi Per poter applicare il test t avevamo detto che le varianze, e
0.1 Condizione sufficiente di diagonalizzabilità
 0.1. CONDIZIONE SUFFICIENTE DI DIAGONALIZZABILITÀ 1 0.1 Condizione sufficiente di diagonalizzabilità È naturale porsi il problema di sapere se ogni matrice sia o meno diagonalizzabile. Abbiamo due potenziali
0.1. CONDIZIONE SUFFICIENTE DI DIAGONALIZZABILITÀ 1 0.1 Condizione sufficiente di diagonalizzabilità È naturale porsi il problema di sapere se ogni matrice sia o meno diagonalizzabile. Abbiamo due potenziali
ESERCITAZIONE SUI PUNTI STAZIONARI DI FUNZIONI LIBERE E SULLE FUNZIONI OMOGENEE
 ESERCITAZIONE SUI PUNTI STAZIONARI DI FUNZIONI LIBERE E SULLE FUNZIONI OMOGENEE 1 Funzioni libere I punti stazionari di una funzione libera di più variabili si ottengono risolvendo il sistema di equazioni
ESERCITAZIONE SUI PUNTI STAZIONARI DI FUNZIONI LIBERE E SULLE FUNZIONI OMOGENEE 1 Funzioni libere I punti stazionari di una funzione libera di più variabili si ottengono risolvendo il sistema di equazioni
Capitolo 9 Verifica di ipotesi: test basati su un campione
 Levine, Krehbiel, Berenson Statistica II ed. 2006 Apogeo Capitolo 9 Verifica di ipotesi: test basati su un campione Insegnamento: Statistica Corsi di Laurea Triennale in Economia Facoltà di Economia, Università
Levine, Krehbiel, Berenson Statistica II ed. 2006 Apogeo Capitolo 9 Verifica di ipotesi: test basati su un campione Insegnamento: Statistica Corsi di Laurea Triennale in Economia Facoltà di Economia, Università
Dal gene alla proteina
 Dal gene alla proteina Il collegamento tra geni e proteine La trascrizione e la traduzione sono i due principali processi che legano il gene alla proteina: uno sguardo panoramico Le informazioni genetiche
Dal gene alla proteina Il collegamento tra geni e proteine La trascrizione e la traduzione sono i due principali processi che legano il gene alla proteina: uno sguardo panoramico Le informazioni genetiche
Lezione 7. Allineamento di sequenze biologiche
 Lezione 7 Allineamento di sequenze biologiche Allineamento di sequenze Determinare la similarità e dedurre l omologia Allineare Definire il numero di passi necessari per trasformare una sequenza nell altra
Lezione 7 Allineamento di sequenze biologiche Allineamento di sequenze Determinare la similarità e dedurre l omologia Allineare Definire il numero di passi necessari per trasformare una sequenza nell altra
06_citologia_SER_golgi 1
 1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
Definizione Impiego Esempi Fonti:
 I rapporti statistici II parte Tassi: generici specifici standardizzati Definizione Impiego Esempi Fonti: Numeri indici: semplici composti TASSI Nel caso di popolazioni p statistiche, le relative statistiche
I rapporti statistici II parte Tassi: generici specifici standardizzati Definizione Impiego Esempi Fonti: Numeri indici: semplici composti TASSI Nel caso di popolazioni p statistiche, le relative statistiche
POPOLAZIONE. Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente
 POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l'unità di base del cambiamento evolutivo, perché permette
POPOLAZIONE Gruppo di individui della stessa specie che occupa una particolare area geografica nella quale essi possono accoppiarsi liberamente E l'unità di base del cambiamento evolutivo, perché permette
RETI DI TELECOMUNICAZIONE
 RETI DI TELECOMUNICAZIONE Modelli delle Sorgenti di Traffico Generalità Per la realizzazione di un modello analitico di un sistema di telecomunicazione dobbiamo tenere in considerazione 3 distinte sezioni
RETI DI TELECOMUNICAZIONE Modelli delle Sorgenti di Traffico Generalità Per la realizzazione di un modello analitico di un sistema di telecomunicazione dobbiamo tenere in considerazione 3 distinte sezioni
Analisi della regressione multipla
 Analisi della regressione multipla y = β 0 + β 1 x 1 + β 2 x 2 +... β k x k + u 2. Inferenza Assunzione del Modello Classico di Regressione Lineare (CLM) Sappiamo che, date le assunzioni Gauss- Markov,
Analisi della regressione multipla y = β 0 + β 1 x 1 + β 2 x 2 +... β k x k + u 2. Inferenza Assunzione del Modello Classico di Regressione Lineare (CLM) Sappiamo che, date le assunzioni Gauss- Markov,
Il modello di regressione lineare multipla. Il modello di regressione lineare multipla
 Introduzione E la generalizzazione del modello di regressione lineare semplice: per spiegare il fenomeno d interesse Y vengono introdotte p, con p > 1, variabili esplicative. Tale generalizzazione diventa
Introduzione E la generalizzazione del modello di regressione lineare semplice: per spiegare il fenomeno d interesse Y vengono introdotte p, con p > 1, variabili esplicative. Tale generalizzazione diventa
Alberi filogenetici. File: alberi_filogenetici.odp. Riccardo Percudani 02/03/04
 Alberi filogenetici The tree of life Albero filogenetico costruito con le sequenze della subunità piccola dell RNA ribosomale. Tutte le forme viventi condividono un comune ancestore (LCA, last common ancestor
Alberi filogenetici The tree of life Albero filogenetico costruito con le sequenze della subunità piccola dell RNA ribosomale. Tutte le forme viventi condividono un comune ancestore (LCA, last common ancestor
CAPITOLO 1 LA FUNZIONE DI PRODUZIONE E LA CRESCITA ECONOMICA
 CAPITOLO 1 LA FUZIOE DI PRODUZIOE E LA CRESCITA ECOOMICA 11 La funzione di produzione Data una funzione di produzione in cui la quantità prodotta () dipende dalla quantità di capitale () e di lavoro ()
CAPITOLO 1 LA FUZIOE DI PRODUZIOE E LA CRESCITA ECOOMICA 11 La funzione di produzione Data una funzione di produzione in cui la quantità prodotta () dipende dalla quantità di capitale () e di lavoro ()
Cenni di apprendimento in Reti Bayesiane
 Sistemi Intelligenti 216 Cenni di apprendimento in Reti Bayesiane Esistono diverse varianti di compiti di apprendimento La struttura della rete può essere nota o sconosciuta Esempi di apprendimento possono
Sistemi Intelligenti 216 Cenni di apprendimento in Reti Bayesiane Esistono diverse varianti di compiti di apprendimento La struttura della rete può essere nota o sconosciuta Esempi di apprendimento possono
7. Gruppi genici e sequenza ripetute
 7. Gruppi genici e sequenza ripetute contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale La duplicazione e la delezione di
7. Gruppi genici e sequenza ripetute contiene materiale protetto da copyright, ad esclusivo uso personale; non è consentita diffusione ed utilizzo di tipo commerciale La duplicazione e la delezione di
Filogenesi molecolare
 Filogenesi molecolare Evoluzione dei geni Gene ancestrale Gene duplicazione genica Gene speciazione Gene 1 Gene 1 ortologhi paraloghi ortologhi Gene 2 Gene 2 Specie 1 Specie 2 Proteine o acidi nucleici?
Filogenesi molecolare Evoluzione dei geni Gene ancestrale Gene duplicazione genica Gene speciazione Gene 1 Gene 1 ortologhi paraloghi ortologhi Gene 2 Gene 2 Specie 1 Specie 2 Proteine o acidi nucleici?
Relazione sequenza-struttura e funzione
 Biotecnologie applicate alla progettazione e sviluppo di molecole biologicamente attive A.A. 2010-2011 Modulo di Biologia Strutturale Relazione sequenza-struttura e funzione Marco Nardini Dipartimento
Biotecnologie applicate alla progettazione e sviluppo di molecole biologicamente attive A.A. 2010-2011 Modulo di Biologia Strutturale Relazione sequenza-struttura e funzione Marco Nardini Dipartimento
Esame di Istituzioni di Matematiche II del 20 giugno 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola
. Cognome Nome Matricola") Esame di Istituzioni di Matematiche II del 20 giugno 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione:
Esame di Istituzioni di Matematiche II del 20 giugno 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione:
Processi di Markov. Processi di Markov
 Processi Stocastici Processi Stocastici Processi Stocastici Catene o Catene o Catene di M Processi Stocastici Processi Stocastici Processi Stocastici Catene o Catene o Catene di M Processi Stocastici Un
Processi Stocastici Processi Stocastici Processi Stocastici Catene o Catene o Catene di M Processi Stocastici Processi Stocastici Processi Stocastici Catene o Catene o Catene di M Processi Stocastici Un
Esercitazione 4: Calcolo e relazioni tra le principali funzioni biometriche delle tavole. Misure di sintesi. Viviana Amati
 Esercitazione 4: Calcolo e relazioni tra le principali funzioni biometriche delle tavole. Misure di sintesi. Viviana Amati 13/05/2009 Le funzioni biometriche della tavola di mortalità Nell esercitazione
Esercitazione 4: Calcolo e relazioni tra le principali funzioni biometriche delle tavole. Misure di sintesi. Viviana Amati 13/05/2009 Le funzioni biometriche della tavola di mortalità Nell esercitazione
Esercitazione 03: Sistemi a tempo discreto
 0 aprile 06 (h) Alessandro Vittorio Papadopoulos alessandro.papadopoulos@polimi.it Fondamenti di Automatica Prof. M. Farina Analisi di investimenti Una banca propone un tasso d interesse i = 3% trimestrale
0 aprile 06 (h) Alessandro Vittorio Papadopoulos alessandro.papadopoulos@polimi.it Fondamenti di Automatica Prof. M. Farina Analisi di investimenti Una banca propone un tasso d interesse i = 3% trimestrale
Riconoscimento e recupero dell informazione per bioinformatica
 Riconoscimento e recupero dell informazione per bioinformatica Clustering: introduzione Manuele Bicego Corso di Laurea in Bioinformatica Dipartimento di Informatica - Università di Verona Una definizione
Riconoscimento e recupero dell informazione per bioinformatica Clustering: introduzione Manuele Bicego Corso di Laurea in Bioinformatica Dipartimento di Informatica - Università di Verona Una definizione
Nulla in biologia ha senso, se non alla luce dell evoluzione
 Nulla in biologia ha senso, se non alla luce dell evoluzione Theodosius Dobzhansky 1973 cambiamento delle proprietà di un gruppo di organismi nel corso delle generazioni Gao et al., 1999 Origin of HIV-1
Nulla in biologia ha senso, se non alla luce dell evoluzione Theodosius Dobzhansky 1973 cambiamento delle proprietà di un gruppo di organismi nel corso delle generazioni Gao et al., 1999 Origin of HIV-1
Capitolo 8. Intervalli di confidenza. Statistica. Levine, Krehbiel, Berenson. Casa editrice: Pearson. Insegnamento: Statistica
 Levine, Krehbiel, Berenson Statistica Casa editrice: Pearson Capitolo 8 Intervalli di confidenza Insegnamento: Statistica Corso di Laurea Triennale in Economia Dipartimento di Economia e Management, Università
Levine, Krehbiel, Berenson Statistica Casa editrice: Pearson Capitolo 8 Intervalli di confidenza Insegnamento: Statistica Corso di Laurea Triennale in Economia Dipartimento di Economia e Management, Università
Esame di Istituzioni di Matematiche II del 11 luglio 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola
. Cognome Nome Matricola") Esame di Istituzioni di Matematiche II del 11 luglio 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione:
Esame di Istituzioni di Matematiche II del 11 luglio 2001 (Corso di Laurea in Biotecnologie, Universitá degli Studi di Padova). Cognome Nome Matricola Es. 1 Es. 2 Es. 3 Es. 4 Somma Voto finale Attenzione:
Secondo parziale di Matematica per l Economia (esempio)
") Corso di Laurea in Economia e Management Secondo parziale di Matematica per l Economia (esempio) lettere E-Z, a.a. 206 207 prof. Gianluca Amato Regole generali Si svolga il primo esercizio e, a scelta
Corso di Laurea in Economia e Management Secondo parziale di Matematica per l Economia (esempio) lettere E-Z, a.a. 206 207 prof. Gianluca Amato Regole generali Si svolga il primo esercizio e, a scelta
UN ANALISI COMPARATA DEL SETTORE DEI TABACCHI LAVORATI IN ITALIA. CASMEF Luiss Guido Carli Audizione presso la Camera dei Deputati 26 Novembre 2013
 UN ANALISI COMPARATA DEL SETTORE DEI TABACCHI LAVORATI IN ITALIA Luiss Guido Carli Audizione presso la Camera dei Deputati 26 Novembre 2013 PREMESSA Situazione corrente Le entrate del Governo nel 2013
UN ANALISI COMPARATA DEL SETTORE DEI TABACCHI LAVORATI IN ITALIA Luiss Guido Carli Audizione presso la Camera dei Deputati 26 Novembre 2013 PREMESSA Situazione corrente Le entrate del Governo nel 2013
ANALISI DELLE SERIE STORICHE
 ANALISI DELLE SERIE STORICHE De Iaco S. s.deiaco@economia.unile.it UNIVERSITÀ del SALENTO DIP.TO DI SCIENZE ECONOMICHE E MATEMATICO-STATISTICHE FACOLTÀ DI ECONOMIA 24 settembre 2012 Indice 1 Funzione di
ANALISI DELLE SERIE STORICHE De Iaco S. s.deiaco@economia.unile.it UNIVERSITÀ del SALENTO DIP.TO DI SCIENZE ECONOMICHE E MATEMATICO-STATISTICHE FACOLTÀ DI ECONOMIA 24 settembre 2012 Indice 1 Funzione di
4 ESERCITAZIONE. Esercizi svolti: Capitolo 5 Modello IS-LM. Esercizio 3 (1 parte)
") 4 ESERCITAZIONE Esercizi svolti: Capitolo 5 Modello IS-LM Esercizio 3 (1 parte) 1 ESERCIZI CAPITOLO 5 MODELLO IS-LM Finora abbiamo considerato SEPARATAMENTE: Equilibrio del mercato dei beni (Cap. 3) 1
4 ESERCITAZIONE Esercizi svolti: Capitolo 5 Modello IS-LM Esercizio 3 (1 parte) 1 ESERCIZI CAPITOLO 5 MODELLO IS-LM Finora abbiamo considerato SEPARATAMENTE: Equilibrio del mercato dei beni (Cap. 3) 1
Ricevimento Studenti: Lunedì previa prenotazione. Cenci lab
 Cenci lab Giovanni Cenci Dip.to Biologia e Biotecnologie C. Darwin Sezione Genetica Piano 2 -Citofono 3/4 0649912-655 (office) 0649912-843 (lab) giovanni.cenci@uniroma1.it Ricevimento Studenti: Lunedì
Cenci lab Giovanni Cenci Dip.to Biologia e Biotecnologie C. Darwin Sezione Genetica Piano 2 -Citofono 3/4 0649912-655 (office) 0649912-843 (lab) giovanni.cenci@uniroma1.it Ricevimento Studenti: Lunedì
Esercizi di Matematica di Base Scienze biologiche e Scienze e Tecnologie dell Ambiente
 Esercizi di Matematica di Base Scienze biologiche e Scienze e Tecnologie dell Ambiente Dati i vettori di R (i) Calcolare il prodotto scalare v w, (ii) Stabilire se v e w sono ortogonali, (ii) Stabilire
Esercizi di Matematica di Base Scienze biologiche e Scienze e Tecnologie dell Ambiente Dati i vettori di R (i) Calcolare il prodotto scalare v w, (ii) Stabilire se v e w sono ortogonali, (ii) Stabilire
Un modello di Markov per la determinazione del rendimento atteso di un obbligazione rischiosa
 Un modello di Markov per la determinazione del rendimento atteso di un obbligazione rischiosa Analisi dei Sistemi Finanziari 1 Giugno, 2007 Cristina Manfredotti Dipartimento di Informatica, Sistemistica
Un modello di Markov per la determinazione del rendimento atteso di un obbligazione rischiosa Analisi dei Sistemi Finanziari 1 Giugno, 2007 Cristina Manfredotti Dipartimento di Informatica, Sistemistica
PROCESSI STOCASTICI 1: ESERCIZI
 PROCESSI STOCASTICI 1: ESERCIZI (1) ESERCIZIO: Date P e Q matrici stocastiche, dimostrare che la matrice prodotto P Q è una matrice stocastica. Dedurre che la potenza P n e il prodotto P 1 P 2 P n sono
PROCESSI STOCASTICI 1: ESERCIZI (1) ESERCIZIO: Date P e Q matrici stocastiche, dimostrare che la matrice prodotto P Q è una matrice stocastica. Dedurre che la potenza P n e il prodotto P 1 P 2 P n sono
IL FLUSSO DELL INFORMAZIONE GENETICA. DNA à RNA. RNA à PROTEINA. DNA à RNA à PROTEINA. Dogma centrale della biologia molecolare di Francis Crick, 1957
 IL FLUSSO DELL INFORMAZIONE GENETICA DNA à RNA à PROTEINA Dogma centrale della biologia molecolare di Francis Crick, 1957 DNA/RNA (seq polinucleotidica) DNA à RNA mrna trna rrna RNA à PROTEINA Proteina
IL FLUSSO DELL INFORMAZIONE GENETICA DNA à RNA à PROTEINA Dogma centrale della biologia molecolare di Francis Crick, 1957 DNA/RNA (seq polinucleotidica) DNA à RNA mrna trna rrna RNA à PROTEINA Proteina
Corso di Statistica Esercitazione 1.8
 Corso di Statistica Esercitazione.8 Test su medie e proporzioni Prof.ssa T. Laureti a.a. 202-203 Esercizio Un produttore vuole monitorare i valori dei livelli di impurità contenute nella merce che gli
Corso di Statistica Esercitazione.8 Test su medie e proporzioni Prof.ssa T. Laureti a.a. 202-203 Esercizio Un produttore vuole monitorare i valori dei livelli di impurità contenute nella merce che gli
07/01/2015. Come si ferma una macchina in corsa? Il terminatore. Terminazione intrinseca (rho-indipendente)
") Come si ferma una macchina in corsa? Il terminatore Terminazione intrinseca (rho-indipendente) Terminazione dipendente dal fattore Rho (r) 1 Operoni: gruppi di geni parte di una unica unità trascrizionale
Come si ferma una macchina in corsa? Il terminatore Terminazione intrinseca (rho-indipendente) Terminazione dipendente dal fattore Rho (r) 1 Operoni: gruppi di geni parte di una unica unità trascrizionale
Ulteriori conoscenze di informatica Elementi di statistica Esercitazione3
 Ulteriori conoscenze di informatica Elementi di statistica Esercitazione3 Sui PC a disposizione sono istallati diversi sistemi operativi. All accensione scegliere Windows. Immettere Nome utente b## (##
Ulteriori conoscenze di informatica Elementi di statistica Esercitazione3 Sui PC a disposizione sono istallati diversi sistemi operativi. All accensione scegliere Windows. Immettere Nome utente b## (##