Regolazione del ciclo cellulare
|
|
|
- Raffaele Nardi
- 5 anni fa
- Visualizzazioni
Transcript



1 Regolazione del ciclo cellulare Copyright by Giorgio Sartor. All rights reserved. Versione /10/ :57 Quattro comportamenti di una cellula in risposta all ambiente v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 2 1

2 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 3 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 4 2


3 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 5 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 6 3

4 3-4h 1h 6-8h 6-12h v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 7 Alberts et al., L ESSENZIALE DI BIOLOGIA MOLECOLARE DELLA CELLULA 3/E, Zanichelli editore S.p.A. Copyright 2011 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 8 4
 (serina-treonina")

5 Controllo È un processo finemente regolato È guidato da 2 classi di molecole: le cicline e le Cyclin-dependent kinases (CDK) (serina-treonina kinasi) che fosforilano altre proteine. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 9 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 10 5

6 Regolazione Il processo si è evoluto negli organismi cellulari più complessi, una sola CDK (CDK1) dei lieviti, che deriva la sua specificità dal legame con diverse Cicline, è stata rimpiazzata da una famiglia di molecole fase-specifiche comprendente circa 20 CDK; Attraverso il legame CDK-Cicline si ha un controllo del ciclo cellulare attraverso diverse combinazioni, in cui le Cicline conferiscono la specificità del substrato e le CDK la funzione enzimatica necessaria per l attivazione dei complessi; Il livello delle Cicline varia nelle fasi del ciclo cellulare mentre quello delle CDK rimane costante essendo la loro attività enzimatica ad essere accesa o spenta dal legame con le Cicline stesse; Il sistema CDK-Cicline fa da ponte tra il segnale mediato dai recettori di superficie a monte e i fattori trascrizionale a valle. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 11 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 12 6


7 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 13 Cicline e CDK v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 14 7
8 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 15 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 16 8
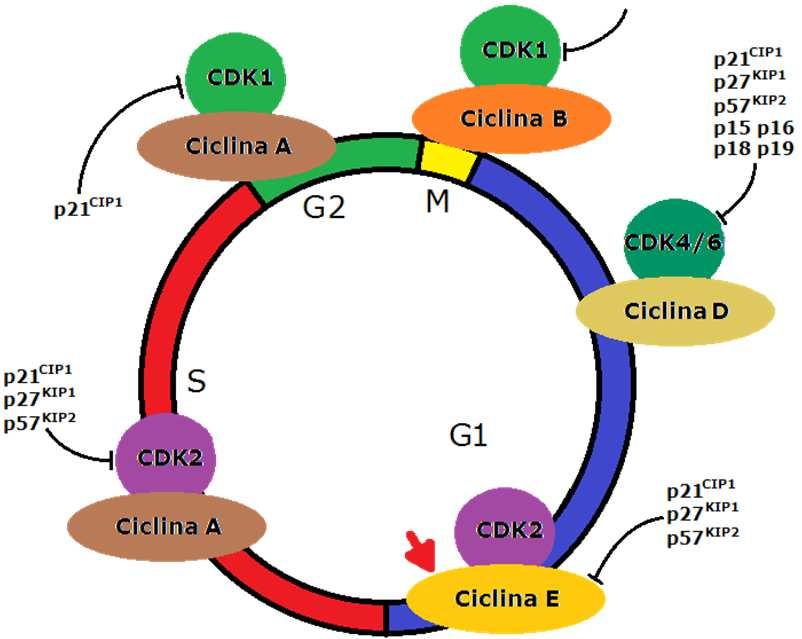
9 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 17 Cicline e CDK CDK Ciclina Funzione CDC2 (CDK1) Ciclina B Fase M CDK2 Ciclina E Fase G1, partenze di Fase S CDK2 Ciclina A Progressione Fase S CDK1 Ciclina A Progressione Fase G2 CDK3? G1 (?) CDK4 Ciclina D Progressione Fase G1 CDK5 p35 Differenziazione Neuronale CDK6 Ciclina D Progressione Fase G1 CDK7 (MO15) Ciclina H CAK, trascrizione CDK8 Ciclina C Trascrizione? Ciclina F Fase G2 o Fase S (?)? Ciclina G? v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 18 9
10 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 19 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 20 10
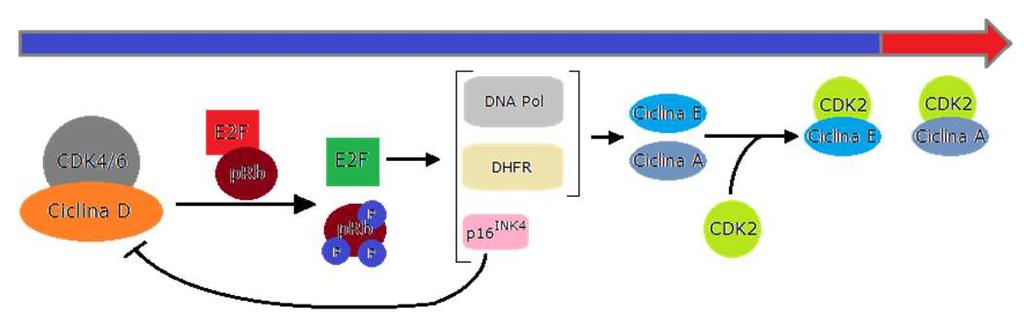
11 Complessi CDK-Cicline v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 21 In fase G1 precoce CDK4/6 sono attivate dalle Cicline D e iniziano la fosforilazione delle proteine del prb; Ciò comporta il rilascio dei fattori di trascrizione E2F cui consegue l attivazione e la trascrizione dei geni responsivi richiesti per la progressione del ciclo cellulare, incluse le Cicline E ed A. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 22 11

 Nell uomo la proteina è codificata dal gene RB1 nel cromosoma 13 (13q14.1-q14.2).")
12 Retinoblastoma protein (prb, RB, Rb o RB1) La proteina retinoblastoma è una tumor suppressor protein. Una funzione di prb consiste nell inibire la progressione del ciclo cellulare fino a quando la cellula non sia pronta per la divisione Si lega ad enzimi che modellano la cromatina (metilasi, acetilasi e deacetilasi) prb appartiene alla famiglia delle pocket protein family che consiste in proteine che possiedono una tasca per il legame ad altre proteine. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 23 Retinoblastoma protein (prb, RB, Rb o RB1) Nell uomo la proteina è codificata dal gene RB1 nel cromosoma 13 (13q14.1-q14.2). Se entrambi gli alleli del gene sono mutati la proteina è inattivata e ciò porta allo sviluppo del retinoblastoma: le cellule della retina non sono rimosse o rimpiazzate e sono quindi soggette ad alti livelli di mutazioni da parte di UV. Sono documentati casi anche in alcuni tumori della pelle di pazienti in Nuova Zelanda dove la radiazione UV è significativamente maggiore. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 24 12
13 Retinoblastoma protein (prb, RB o RB1) La funzione di prb è quella di prevenire la progressione nel ciclo (da G1 a S) di cellule con DNA danneggiato; prb lega ed inibisce i fattori di trascrizione della famiglia E2F composti di eterodimeri E2F e DP (dimerization partner); I complessi E2F-DP attivati spingono la cellula nella fase S mentre se sono inattivati da prb la cellula rimane in fase G1; Il complesso prb-e2f/dp è inoltre coinvolto nell interazione dell istone deacetilasi con la cromatina riducendo la trascrizione di promotori coinvolti nel passaggio in fase S riducendo ulteriormente la sintesi del DNA. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 25 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 26 13
. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 27 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 28 14")
14 E2F Fattori di trascrizioni che legano la sequenza TTTCCCGC (o varianti) CycA: dominio di legame della Ciclina A DNA: dominio di legame al DNA DP1,2: dominio di dimerizzazione con DP1 e DP2 TA: dominio di attivazione trascrizionale PB: dominio di legame della pocket protein (prb). v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 27 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 28 14


15 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 29 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 30 15

16 Funzionamento di prb La proteina presenta un pocket domain composto di due regioni: A- box e B-box; Il cristallo del pocket domain di prb complessato con il peptide di 18 AA del dominio di transattivazione di E2F-2 mostra che esso si lega nell interfaccia altamente conservata tra A-box e B-box. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 31 A-box 1N4M B-box Structural basis for the recognition of the E2F transactivation domain by the retinoblastoma tumor suppressor. Lee, C., Chang, J.H., Lee, H.S., Cho, Y. GENES DEV. (2002) 16: v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 32 16
 è essenziale per il legame di E2F al Rb")
17 Funzionamento di prb Il segmento N-terminale del peptide E2F-2 è in una struttura estesa simile a un β-strand che interagisce con le α-eliche del solco all interfaccia A-B; Il segmento C-terminale del peptide contiene un elica 3 10 che si lega alle eliche di A-box La flessibilità della porzione mediana del peptide di transattivazione di E2F-2 (Gly) è essenziale per il legame di E2F al Rb pocket. Il legame di prb al peptide di E2F-2 peptide nasconde diversi residui conservati di E2F che sono cruciali per l attivazione della trascrizione. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 33 N-term A-box 1N4M B-box C-term Structural basis for the recognition of the E2F transactivation domain by the retinoblastoma tumor suppressor. Lee, C., Chang, J.H., Lee, H.S., Cho, Y. GENES DEV. (2002) 16: v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 34 17
18 N-term A-box 1N4M B-box C-term Structural basis for the recognition of the E2F transactivation domain by the retinoblastoma tumor suppressor. Lee, C., Chang, J.H., Lee, H.S., Cho, Y. GENES DEV. (2002) 16: v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 35 Ser 1N4M Thr v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 36 18



19 Ser 1N4M Thr v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 37 Interazione prb-e2f 1N4M 2AZE Oltre la tasca di prb è anche necessario, per il legame con E2F, anche il dominio C-terminale La struttura del complesso RbC-E2F1-DP1 mostra un avvolgimento nel quale l eterodimero E2F-DP è a contatto con il dominio C-terminale di prb. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 38 19
20 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 39 Funzionamento di prb Molte proteine virali (Papilloma virus) e cellulari che inattivano prb legando la regione B-box attraverso una sequenza LxCxE; La struttura del pocket domain di prb, legato ad un nonapeptide che contiene il motivo LxCxE, (1GUX) dimostra che il peptide lega una sequenza altamente conservata che genera un solco in B-box; Il dominio A-box sembra essere anche necessario per la stabilizzazione di B-box. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 40 20


 391: 859-865 v. 0.")
21 prb 1GUX 1N4M v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 41 Glu Cys Leu Leu Glu B-box A-box 1GUX Structure of the retinoblastoma tumour-suppressor pocket domain bound to a peptide from HPV E7. Lee, J.O., Russo, A.A., Pavletich, N.P. Nature (1998) 391: v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 42 21


22 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 43 Cicline e Cdk In fase G1 precoce CDK4/6 sono attivate dalle cicline D e si inizia la fosforilazione di prb; Si rilasciano i fattori di trascrizione E2F; Si ha l attivazione e la trascrizione dei geni responsivi richiesti per la progressione del ciclo cellulare, incluse quelli delle cicline E ed A; In fase G1 tardiva CDK2 è attivata dal legame alla ciclina E e completa la fosforilazione delle proteine prb portando alla ulteriore attivazione dei fattori di trascrizione E2F v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 44 22
, elica PSTAIRE.")

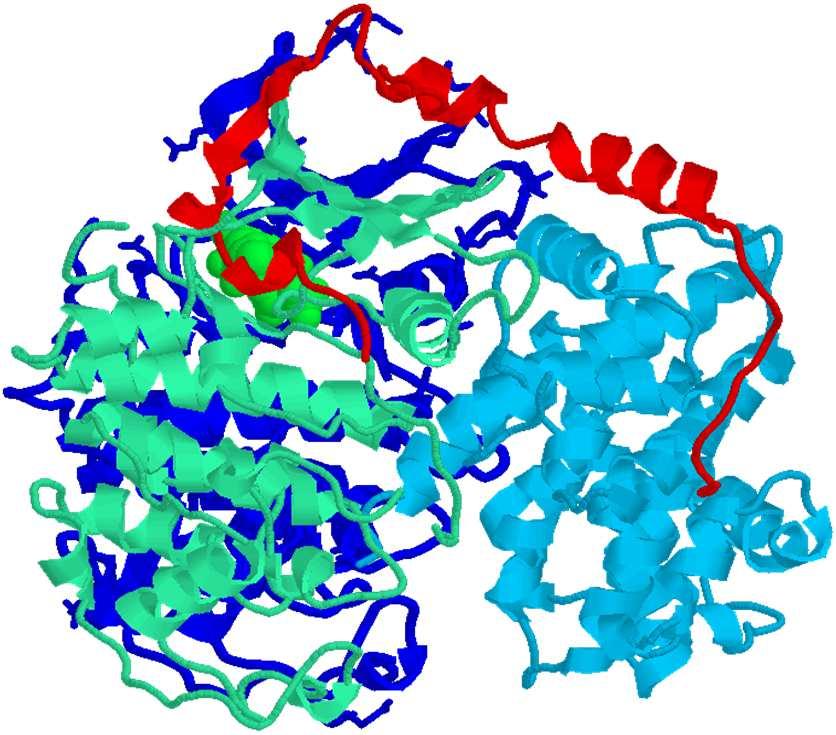
23 Cdk2-Ciclina A La CDK2 possiede due domini: Dominio amminoterminale, costituito da una singola α elica e da un foglietto β, a 5 filamenti. L α elica contiene una sequenza di sei residui (Pro-Ser-Thr-Ala-Ile-Arg- Glu), elica PSTAIRE. uno più grande formato da α eliche, possiede una regione flessibile (T loop) contenente un residuo di treonina, Thr160 fosforilato nella CDK attiva. L ATP è legato in una fenditura tra i 2 domini. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 45 Cdk2-Ciclina A La CDK2 possiede due domini: Dominio amminoterminale, costituito da una singola α elica e da un foglietto β, a 5 filamenti. L α elica contiene una sequenza di sei residui (Pro-Ser-Thr-Ala-Ile-Arg- Glu), elica PSTAIRE. uno più grande formato da α eliche, possiede una regione flessibile (T loop) contenente un residuo di treonina, Thr160 fosforilato nella CDK attiva. L ATP è legato in una fenditura tra i 2 domini. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 46 23

24 Cdk2-Ciclina A Il frammento della ciclina A ( ) è costituito da due domini, caratterizzati dalla presenza di α eliche, questo frammento è in grado di attivare la CDK2. Il primo dominio possiede una sequenza box fortemente conservata in tutte le cicline. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 47 Cdk2-Ciclina A Nel complesso ciclina A- CDK2, la sequenza della ciclina interagisce con la CDK2, principalmente con l elica PSTAIRE ed il loop T; La struttura della ciclina A all interno del complesso non subisce modificazioni rispetto alla molecola isolata, mentre la struttura dell CDK2 e soggetta a cambiamenti conformazionali rilevanti. L intero dominio N-terminale presenta un orientamento diverso rispetto al C- terminale. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 48 24


25 Cdk2-Ciclina A 1FIN Nel complesso ciclina A- Cdk2, la sequenza della ciclina interagisce con la Cdk2, principalmente con l elica PSTAIRE ed il loop T; La struttura della ciclina A all interno del complesso non subisce modificazioni rispetto alla molecola isolata, mentre la struttura dell Cdk2 e soggetta a cambiamenti conformazionali rilevanti. L intero dominio N-terminale presenta un orientamento diverso rispetto al C- terminale. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 49 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 50 25

26 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 51 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 52 26



27 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 53 Elica PSTAIRE L elica PSTAIRE si sposta verso la fenditura del sito attivo della Cdk2 ruotando di circa 90 ; Il residuo catalitico essenziale di Glu51 risulta diretto verso l interno della fenditura invece che all esterno, come succede nella Cdk2 isolata. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 54 27
28 Cdk 1FIN v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 55 Elica PSTAIRE ATP 1HCL 1FIN v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 56 28



29 T-Loop 1HCL 1FIN v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 57 Cdk 1HCL 1FIN v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 58 29


30 La conseguenza di questi movimenti concertati è il movimento di alcuni atomi della catena principale dell elica. Oltre alla variazione della posizione dell elica PSTAIRE, si ha anche un riarrangiamento del T loop; Nell adottare una posizione completamente nuova, il T loop subisce anche una profonda trasformazione strutturale: nella Cdk2 isolata possiede una struttura ad α-elica assume all interno del complesso una struttura a filamento β. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 59 ATP Mg ++ 1JST v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 60 30


31 L attività dei complessi CDK/ciclina viene implementata da chinasi specifiche: CDK activating kinases (CAK) attraverso due meccanismi complementari: La fosforilazione in residui di treonina nel dominio T-loop che possono o incrementare l attività del complesso CDK/ciclina già formato o favorire il legame di CDK con la ciclina. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 61 Nella molecola di Cdk2 isolata, la fenditura del sito attivo è ostruita dal T loop e il residuo Thr160 è all interno. In queste condizioni i substrati non possono legarsi e la Thr160 non può essere fosforilata: la Cdk2 isolata è inattiva. I cambiamenti conformazionali indotti dal legame della Ciclina A espongono la fenditura del sito attivo, in modo che l ATP e le proteine substrato possono essere legate e determinano anche un riarrangiamento strutturale a livello dei residui del sito attivo che rende l enzima cataliticamente competente. Il residuo Thr160 viene esposto e fosforilabile, con aumento dell attività catalitica. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 62 31

32 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 63 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 64 32
 nel dominio di legame con l ATP che funzionano, quindi, di inibitori o indotta")

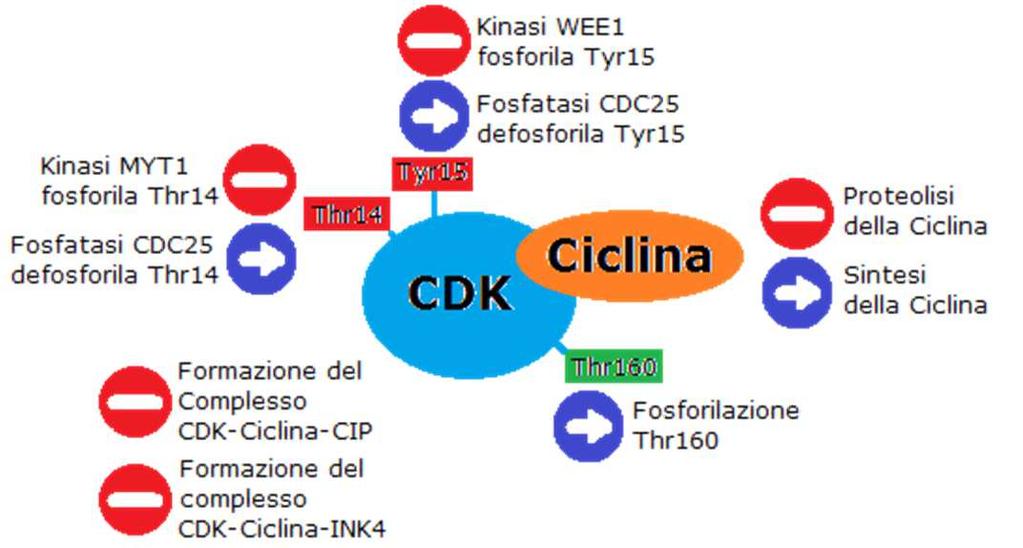
33 L attività kinasica di CDK può essere: repressa da due chinasi WEE1 e MYT1 che fosforilano due residui di CDK1 (Thr14 e Tyr15) nel dominio di legame con l ATP che funzionano, quindi, di inibitori o indotta da una classe di fosfatasi CDC25 che defosforila tali residui, riattivando il complesso. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 65 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 66 33
 che inibiscono i complessi CDK2/ciclina E, CDK2/ciclina A e CDK1/ciclina B,")
34 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 67 Inibizione L attività dei complessi CDK/ciclina viene inibita da due famiglie di CDK inhibitors: INK4 (p16, p15, p18 e p19) che specificatamente legano CDK4 e CDK6 e prevengono l attivazione da parte delle cicline D CIP/KIP (p21, p27 e p57) che inibiscono i complessi CDK2/ciclina E, CDK2/ciclina A e CDK1/ciclina B, intervenendo in tutte le fasi del ciclo cellulare. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 68 34

35 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 69 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 70 35
. v. 0.")
36 INK4 I quattro membri della famiglia genica INK4 (p16 INK4a, p15 INK4b, p18 INK4c e p19 INK4d ) inibiscono Cdk4 e Cdk6 e partecipano alla regolazione della transizione G1 S; La perdita della funzione del gene INK4, in particolare di p16 INK4a, è presente in molti tumori umani; Meno frequenti sono i difetti associati a p19 INK4d (osteosarcoma). v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 71 P16 INK4a è un inibitore specifico per la Cdk4/6, appartiene a una famiglia includendo p15 INK4b, p18 INK4c e p19 INK4d p16 e p19 sono costituite, rispettivamente, da 4 o 5 motivi ad anchirina Il motivo strutturale è costituito da residui di circa 30 aa, ha una struttura a L con due α-eliche antiparallele, che formano il gambo e la struttura β a forcina che forma la base. INK4 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 72 36



37 INK4 p16 e p19 hanno delle regioni conservate che sono quelle che servono per l interazione con le Cdk4/6. I motivi ad anchirina mantengono conservati gli elementi di struttura secondaria. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 73 p16 INK4a e p19 INK4d 2A5E 1BD8 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 74 37


38 Meccanismo p16 interagisce con la Cdk6 in un solco opposto a quello della ciclina, ma localizzato tra i 2 domini. La superficie concava della p16 lega la porzione N-terminale a foglietto β, mentre con i motivi ad anchirina interagisce con la parte C-terminale. Il legame e il riconoscimento sono mediati all inizio da legami idrogeno con diversi residui che partecipano in questa interazione. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 75 Meccanismo p16 lega la Cdk6 e causa una modificazione conformazionale con conseguente inattivazione enzimatica. La p16 prende contatto con la Cdk6 in un solco tra le catene N- terminale e C- terminale alterando il sito di legame per la ciclina e bloccando la traslocazione dell elica PSTAIRE nel solco catalitico. Genes Dev December 15; 14(24): v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 76 38


39 p16 INK4a e p19 INK4d - Cdk6 1BI7 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 77 p16 INK4a 1BI7 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 78 39


40 p19 INK4d 1BLX v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 79 Cdk2 Ciclina A p16 INK4a Cdk6 1JST 1BI7 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 80 40
: 3115 3125. The ATP-binding site of p18 Cdk6 K-cyclin and Cdk2 cyclina.")

41 Il networks di legami idrogeno sono conservati nel II e III motivo ad anchirina sia nella p16 che nella p19 e giocano un ruolo importante nel legame con la Cdk6. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 81 Genes Dev December 15; 14(24): The ATP-binding site of p18 Cdk6 K-cyclin and Cdk2 cyclina. Active site residues implicated in ATP binding and catalysis are displaced in the p18 Cdk6 K-cyclin complex relative to the active Cdk2 cyclina conformation. Cdk2 and Cdk6 were superimposed on their C lobes. Cdk6 is shown in cyan, p18 in yellow, Cdk2 in gray. Movement of active site residues is indicated by red arrows. p18 displaces the N lobe relative to the C lobe, causing the hydrophobic residues (Ile 19, Val 27, Ala 41, Leu 152) that sandwich the adenine ring of ATP to move by up to 4.5 A. The p18 inhibitor also distorts the edge of the active site via Phe 82, affecting hydrogen bonding interactions with the edge of the ATP ring. The related shift of the PSTAIRE helix on the other side of the active site displaces an active site residue (Glu 61). The T loop of Cdk6 diverges from that of Cdk2 between Phe 164 and Val 181, and these residues are omitted for clarity. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 82 41

42 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 83 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 84 42
43 1JSU CIP/KIP CIP/KIP sono una famiglia di inibitori, in cui si riconoscono principalmente p21 e p27 Si legano al complesso Cdk2- Ciclina A fosforilato interagendo sia con la Cdk2 che con la Ciclina A. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 85 1JSU CIP/KIP p27 KIP1 lega il complesso attraverso interazioni sia con la Ciclina A che con Cdk2. Si lega alla Ciclina A nel solco formato dai residui conservati Si lega a Cdk2 e riarrangia il dominio amino terminale Su CdK2 si inserisce nel sito di legame dell ATP. v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 86 43


44 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 87 1JSU 1FIN v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 88 44

45 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 89 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 90 45


46 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 91 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 92 46

47 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 93 Crediti e autorizzazioni all utilizzo Questo materiale è stato assemblato da informazioni raccolte dai seguenti testi di Biochimica: CHAMPE Pamela, HARVEY Richard, FERRIER Denise R. LE BASI DELLA BIOCHIMICA [ISBN ] Zanichelli NELSON David L., COX Michael M. I PRINCIPI DI BIOCHIMICA DI LEHNINGER - Zanichelli GARRETT Reginald H., GRISHAM Charles M. BIOCHIMICA con aspetti molecolari della Biologia cellulare - Zanichelli VOET Donald, VOET Judith G, PRATT Charlotte W FONDAMENTI DI BIOCHIMICA [ISBN ] - Zanichelli E dalla consultazione di svariate risorse in rete, tra le quali: Kegg: Kyoto Encyclopedia of Genes and Genomes Brenda: Protein Data Bank: Il materiale è stato inoltre rivisto e corretto dalla Prof. Giancarla Orlandini dell Università di Parma alla quale va il mio sentito ringraziamento. Questo ed altro materiale può essere reperito a partire da: gsartor.org/pro Il materiale di questa presentazione è di libero uso per didattica e ricerca e può essere usato senza limitazione, purché venga riconosciuto l autore usando questa frase: Materiale ottenuto dal Prof. Giorgio Sartor Alma Mater Studiorum Giorgio Sartor - giorgio.sartor@unibo.it 18/10/ :57:12 v. 0.7 gsartor 2018 Q01 - Regolazione del ciclo cellulare 94 47
Regolazione del ciclo cellulare
 Regolazione del ciclo cellulare Copyright 2001-2013 by Giorgio Sartor. All rights reserved. Versione 0.1 nov2013 Quattro comportamenti di una cellula in risposta all ambiente v. 0.1 gsartor 2013 Q01 -
Regolazione del ciclo cellulare Copyright 2001-2013 by Giorgio Sartor. All rights reserved. Versione 0.1 nov2013 Quattro comportamenti di una cellula in risposta all ambiente v. 0.1 gsartor 2013 Q01 -
Regolazione del ciclo cellulare
 Regolazione del ciclo cellulare Copyright 2001-2013 by Giorgio Sartor. All rights reserved. Versione 0.1 nov2013 Quattro comportamenti di una cellula in risposta all ambiente v. 0.1 gsartor 2013 Q01 -
Regolazione del ciclo cellulare Copyright 2001-2013 by Giorgio Sartor. All rights reserved. Versione 0.1 nov2013 Quattro comportamenti di una cellula in risposta all ambiente v. 0.1 gsartor 2013 Q01 -
Regolazione del ciclo cellulare
 Regolazione del ciclo cellulare Copyright 2001-2014 by Giorgio Sartor. All rights reserved. Versione 0.5 dec 2014 Quattro comportamenti di una cellula in risposta all ambiente v. 0.5 gsartor 2014 Q01 -
Regolazione del ciclo cellulare Copyright 2001-2014 by Giorgio Sartor. All rights reserved. Versione 0.5 dec 2014 Quattro comportamenti di una cellula in risposta all ambiente v. 0.5 gsartor 2014 Q01 -
MOVIMENTO. Motori molecolari. IV Movimento Miosina e actina. Prof. Giorgio Sartor. P04 - Versione 2.0 oct 2013
 Prof. Giorgio Sartor Motori molecolari IV Movimento Miosina e actina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P04 - Versione 2.0 oct 2013 MOVIMENTO Miosina Actina P04 v. 2.0 gsartor
Prof. Giorgio Sartor Motori molecolari IV Movimento Miosina e actina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P04 - Versione 2.0 oct 2013 MOVIMENTO Miosina Actina P04 v. 2.0 gsartor
Prof. Giorgio Sartor. Invertebrati marini. I nutrienti. Fonti di carbonio ed energia. Fonte di carbonio. Fonte di energia. Composti organici.
 Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
Prof. Giorgio Sartor Invertebrati marini I nutrienti Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.1 nov 2008 Fonti di carbonio ed energia Organismo Fonte di carbonio Fonte di
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2008 by Giorgio Sartor. All rights reserved. Versione 1.4 oct 2008 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
MIOSINA. Motori molecolari. II Strutture per il movimento Miosina, Kinesina e Dineina. Una struttura per il movimento. Prof.
 Prof. Giorgio Sartor Motori molecolari II Strutture per il movimento Miosina, Kinesina e Dineina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P02 - Versione 2.0 oct 2013 MIOSINA Una struttura
Prof. Giorgio Sartor Motori molecolari II Strutture per il movimento Miosina, Kinesina e Dineina Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P02 - Versione 2.0 oct 2013 MIOSINA Una struttura
Prof. Giorgio Sartor. Il metabolismo. Metabolismo. È il processo che permette di ricavare energia da legami chimici (sottoforma di
 Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Prof. Giorgio Sartor Il metabolismo Copyright 2001-2017 by Giorgio Sartor. All rights reserved. Versione 1.6.1 Mar-17 Metabolismo È il processo che permette di ricavare energia da legami chimici (sottoforma
Geni che regolano la divisione cellulare
 Geni che regolano la divisione cellulare!geni il cui prodotto proteico promuove un aumento del numero di cellule (oncogeni) Oncogeni = acceleratori!geni il cui prodotto proteico induce una riduzione del
Geni che regolano la divisione cellulare!geni il cui prodotto proteico promuove un aumento del numero di cellule (oncogeni) Oncogeni = acceleratori!geni il cui prodotto proteico induce una riduzione del
MIOSINE. Motori molecolari. II Miosina, Kinesine e Dineine Strutture per il movimento. Prof. Giorgio Sartor. M09 - Versione 1.
 Prof. Giorgio Sartor Motori molecolari II Miosina, Kinesine e Dineine Strutture per il movimento Copyright 2001-2018 by Giorgio Sartor. All rights reserved. M09 - Versione 1.7 02/10/2018 MIOSINE 1 Miosina/e
Prof. Giorgio Sartor Motori molecolari II Miosina, Kinesine e Dineine Strutture per il movimento Copyright 2001-2018 by Giorgio Sartor. All rights reserved. M09 - Versione 1.7 02/10/2018 MIOSINE 1 Miosina/e
Metabolismo dei nucleotidi
 rof. Giorgio Sartor Metabolismo dei nucleotidi 1: introduzione 2: biosintesi de-novo delle purine 3: biosintesi de-novo delle pirimidine 4: catabolismo delle purine 5: catabolismo delle pirimidine 6: sintesi
rof. Giorgio Sartor Metabolismo dei nucleotidi 1: introduzione 2: biosintesi de-novo delle purine 3: biosintesi de-novo delle pirimidine 4: catabolismo delle purine 5: catabolismo delle pirimidine 6: sintesi
KINESINA. Motori molecolari. V Movimento Kinesina, Dineina su microtubuli. Prof. Giorgio Sartor. P05 - Versione 2.0 oct 2013
 Prof. Giorgio Sartor Motori molecolari V Movimento Kinesina, Dineina su microtubuli Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P05 - Versione 2.0 oct 2013 KINESINA C-terminale (coda) Kinesina
Prof. Giorgio Sartor Motori molecolari V Movimento Kinesina, Dineina su microtubuli Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P05 - Versione 2.0 oct 2013 KINESINA C-terminale (coda) Kinesina
I FLAGELLI NEI BATTERI
 Prof. Giorgio Sartor Motori molecolari VI Flagello batterico Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P06 - Versione 2.0 oct 2013 I FLAGELLI NEI BATTERI Un diverso movimento Un diverso
Prof. Giorgio Sartor Motori molecolari VI Flagello batterico Copyright 2001-2013 by Giorgio Sartor. All rights reserved. P06 - Versione 2.0 oct 2013 I FLAGELLI NEI BATTERI Un diverso movimento Un diverso
REGOLAZIONE DEL CICLO CELLULARE.
 REGOLAZIONE DEL CICLO CELLULARE www.fisiokinesiterapia.biz Siti utili Animazione sul ciclo cellulare e su come è stata scoperta la sua regolazione: http://www.usask.ca/biology/biol316/visgen/cell.mov Sito
REGOLAZIONE DEL CICLO CELLULARE www.fisiokinesiterapia.biz Siti utili Animazione sul ciclo cellulare e su come è stata scoperta la sua regolazione: http://www.usask.ca/biology/biol316/visgen/cell.mov Sito
I FLAGELLI NEI BATTERI
 Prof. Giorgio Sartor Motori molecolari V Flagello batterico Copyright 2001-2015 by Giorgio Sartor. All rights reserved. M09 - Versione 1.7 oct 2015 I FLAGELLI NEI BATTERI Un diverso movimento Un diverso
Prof. Giorgio Sartor Motori molecolari V Flagello batterico Copyright 2001-2015 by Giorgio Sartor. All rights reserved. M09 - Versione 1.7 oct 2015 I FLAGELLI NEI BATTERI Un diverso movimento Un diverso
Omnis cellula e cellula
 ogni cellula deriva da un altra cellula Omnis cellula e cellula Virchow 1858 LA MAGGIOR PARTE DEI PROCESSI NUCLEARI E CELLULARI DURANTE LA MITOSI E INDISTINGUIBILE IN PIANTE,ANIMALI E FUNGHI. DIVISIONE
ogni cellula deriva da un altra cellula Omnis cellula e cellula Virchow 1858 LA MAGGIOR PARTE DEI PROCESSI NUCLEARI E CELLULARI DURANTE LA MITOSI E INDISTINGUIBILE IN PIANTE,ANIMALI E FUNGHI. DIVISIONE
REGOLAZIONE DELL ATTIVITA ENZIMATICA 1) MODULAZIONE ALLOSTERICA NON-COVALENTE (REVERSIBILE)
 MODULAZIONE ALLOSTERICA NON-COVALENTE (REVERSIBILE)") REGOLAZIONE DELL ATTIVITA ENZIMATICA - Regolazione a lungo termine: la quantità di enzima può essere controllata regolando la velocità della sua sintesi o degradazione (minuti o ore) - Regolazione a breve
REGOLAZIONE DELL ATTIVITA ENZIMATICA - Regolazione a lungo termine: la quantità di enzima può essere controllata regolando la velocità della sua sintesi o degradazione (minuti o ore) - Regolazione a breve
REGOLAZIONE DELLA TRASCRIZIONE NEGLI EUCARIOTI
 LEZIONE XI REGOLAZIONE DELLA TRASCRIZIONE NEGLI EUCARIOTI Dott. Paolo Cascio IL PROMOTORE DEL VIRUS SV 40 PRESENTA 1 SEQUENZA TATA E 3 CG BOX. PIU LONTANO, PERO, SONO SITUATE ALTRE 2 REGIONI PIUTTOSTO
LEZIONE XI REGOLAZIONE DELLA TRASCRIZIONE NEGLI EUCARIOTI Dott. Paolo Cascio IL PROMOTORE DEL VIRUS SV 40 PRESENTA 1 SEQUENZA TATA E 3 CG BOX. PIU LONTANO, PERO, SONO SITUATE ALTRE 2 REGIONI PIUTTOSTO
Emivita delle proteine
 Emivita delle proteine Ornitina decarbossilasi: 30 min, HMGCoA red: ~ 2-4 h Possiedono alto turn over inoltre: Enzimi nella biosintesi dei nucleotidi timinici (Ribonucleotide red, timidilato sintetasi)
Emivita delle proteine Ornitina decarbossilasi: 30 min, HMGCoA red: ~ 2-4 h Possiedono alto turn over inoltre: Enzimi nella biosintesi dei nucleotidi timinici (Ribonucleotide red, timidilato sintetasi)
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata. Ciclo Cellulare
 Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
Struttura delle proteine
 Struttura delle proteine I II III Copyright 2001-2015 by Giorgio Sartor. All rights reserved. Versione 1.0.2 oct 2015 Struttura quaternaria Èil livello di organizzazione per il quale si formano strutture
Struttura delle proteine I II III Copyright 2001-2015 by Giorgio Sartor. All rights reserved. Versione 1.0.2 oct 2015 Struttura quaternaria Èil livello di organizzazione per il quale si formano strutture
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare
 Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
Le cellule eucariotiche svolgono durante la loro vita una serie ordinata di eventi che costituiscono il Ciclo Cellulare Interfase comprende le fasi G 1, S, and G 2 Sintesi di macromolecole durante la
Alberts et al., L ESSENZIALE DI BIOLOGIA MOLECOLARE DELLA CELLULA, Zanichelli editore S.p.A. Copyright 2005
 La conversione dell informazione da una forma ad un altra è il punto critico della trasmissione e prende il nome di: TRASDUZIONE DEL SEGNALE Nella comunicazione cellulare: Molecola segnale Proteina recettore
La conversione dell informazione da una forma ad un altra è il punto critico della trasmissione e prende il nome di: TRASDUZIONE DEL SEGNALE Nella comunicazione cellulare: Molecola segnale Proteina recettore
Acido etilendiamminotetraacetico. Spettrometria di massa ESI. Isopropil-β-D-tiogalattopiranoside
 ABBREVIAZIONI ATP cdna DTT EDTA ESI ESI/MS HEPES IPTG LB MALDI MALDI/MS MOPS p27 wt PMSF SDS TFA TRIS t-rna Adenosina 5 -trifosfato DNA copia Ditiotreitolo Acido etilendiamminotetraacetico Electrospray
ABBREVIAZIONI ATP cdna DTT EDTA ESI ESI/MS HEPES IPTG LB MALDI MALDI/MS MOPS p27 wt PMSF SDS TFA TRIS t-rna Adenosina 5 -trifosfato DNA copia Ditiotreitolo Acido etilendiamminotetraacetico Electrospray
Tirosin chinasi senza o con funzione recettoriale
 Tirosin chinasi senza o con funzione recettoriale elica di poliprolina Il dominio chinasico di Src è mantenuto nello stato inattivo mediante due distinte interazioni intramolecolari: il legame del dominio
Tirosin chinasi senza o con funzione recettoriale elica di poliprolina Il dominio chinasico di Src è mantenuto nello stato inattivo mediante due distinte interazioni intramolecolari: il legame del dominio
2) La presenza di gruppi funzionali specifici che partecipano alla catalisi (quelli delle catene laterali dei suoi residui amminoacidici e/o quelli
 La presenza di gruppi funzionali specifici che partecipano alla catalisi (quelli delle catene laterali dei suoi residui amminoacidici e/o quelli") ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari (sono in grado di aumentare la velocità delle reazioni biologiche anche di 10 20 volte). Durante
ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari (sono in grado di aumentare la velocità delle reazioni biologiche anche di 10 20 volte). Durante
REGOLAZIONE DEL CICLO CELLULARE
 REGOLAZIONE DEL CICLO CELLULARE Il sistema di controllo che regola la progressione del ciclo cellulare deve: 1) Garantire che tutti i processi associati con le diverse fasi siano portati a termine al tempo
REGOLAZIONE DEL CICLO CELLULARE Il sistema di controllo che regola la progressione del ciclo cellulare deve: 1) Garantire che tutti i processi associati con le diverse fasi siano portati a termine al tempo
Mutazioni nelle cellule somatiche e genetica del cancro. (Teoria Boveri - Sutton)
") Mutazioni nelle cellule somatiche e genetica del cancro (Teoria Boveri - Sutton) Walter Stanborough Sutton Theodor Boveri Walter S. Sutton: Teoria dell ereditarietà dei cromosomi 1914: Teoria cromosomica
Mutazioni nelle cellule somatiche e genetica del cancro (Teoria Boveri - Sutton) Walter Stanborough Sutton Theodor Boveri Walter S. Sutton: Teoria dell ereditarietà dei cromosomi 1914: Teoria cromosomica
Recettori di superficie
 Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
Recettori di superficie
 Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
Recettori di superficie Esistono 3 classi principali di recettori di superficie 1. Recettori annessi a canali ionici 2. Recettori accoppiati alle proteine G 3. Recettori associati ad enzimi Recettori
I meccanismi di catalisi della sintesi di DNA e RNA sono identici
 I meccanismi di catalisi della sintesi di DNA e RNA sono identici U U OH Ribo-nucleotide trifosfato TRASCRIZIONE La RNA polimerasi è totalmente processiva Non ha bisogno di innesco Inizia a livello di
I meccanismi di catalisi della sintesi di DNA e RNA sono identici U U OH Ribo-nucleotide trifosfato TRASCRIZIONE La RNA polimerasi è totalmente processiva Non ha bisogno di innesco Inizia a livello di
Regolazione dell espressione genica negli eucarioti. Prof. Savino; dispense di Biologia Molecolare, Corso di Laurea in Biotecnologie
 Regolazione dell espressione genica negli eucarioti Il promotore eucariotico L inizio della trascrizione negli eucarioti necessita della RNA polimerasi e dei fattori di trascrizione. Qualsiasi proteina
Regolazione dell espressione genica negli eucarioti Il promotore eucariotico L inizio della trascrizione negli eucarioti necessita della RNA polimerasi e dei fattori di trascrizione. Qualsiasi proteina
La regolazione genica negli eucarioti
 La regolazione genica negli eucarioti neuroni globuli rossi globulo bianco fibroblasti adipociti Sezione di testicolo Surrene Come mai alcuni geni sono trascritti e tradotti in alcune cellule ma non in
La regolazione genica negli eucarioti neuroni globuli rossi globulo bianco fibroblasti adipociti Sezione di testicolo Surrene Come mai alcuni geni sono trascritti e tradotti in alcune cellule ma non in
La trascrizione nei procarioti SINTESI RNA
 La trascrizione nei procarioti SINTESI RNA https://www.youtube.com/watch?v=w8a7kxz3eqo https://www.youtube.com/watch?v=j2rb8dpclei La replicazione è finalizzata alla proliferazione cellulare La trascrizione
La trascrizione nei procarioti SINTESI RNA https://www.youtube.com/watch?v=w8a7kxz3eqo https://www.youtube.com/watch?v=j2rb8dpclei La replicazione è finalizzata alla proliferazione cellulare La trascrizione
Fattori di crescita. Membrana citoplasmatica. Recettori di fattori di crescita. Proteine trasduttrici del segnale. Nucleo. Fattori trascrizionali
 Fattori di crescita Recettori di fattori di crescita Membrana citoplasmatica roteine trasduttrici del segnale Nucleo Fattori trascrizionali roteine del ciclo cellulare Ciclo di divisione cellulare Fattori
Fattori di crescita Recettori di fattori di crescita Membrana citoplasmatica roteine trasduttrici del segnale Nucleo Fattori trascrizionali roteine del ciclo cellulare Ciclo di divisione cellulare Fattori
De Leo - Fasano - Ginelli Biologia e Genetica, II Ed. Capitolo 7. Replicazione del DNA Preparazione alla divisione Citodieresi
 Replicazione del DNA Preparazione alla divisione Citodieresi Durata del ciclo cellulare: 24 ore nei fibroblasti; 2 ore nel lievito; 30 minuti nei procarioti Quando una cellula non si divide per un periodo
Replicazione del DNA Preparazione alla divisione Citodieresi Durata del ciclo cellulare: 24 ore nei fibroblasti; 2 ore nel lievito; 30 minuti nei procarioti Quando una cellula non si divide per un periodo
CATALISI COVALENTE. H 2 O Enz-R-OH + P. Enz-R-O S Enz-R-O-S
 CATALISI COVALENTE Comporta la formazione di intermedi in cui il substrato è legato covalentemente con un residuo amminoacidico del sito attivo dell enzima. La catena laterale re dell enzima può comportarsi
CATALISI COVALENTE Comporta la formazione di intermedi in cui il substrato è legato covalentemente con un residuo amminoacidico del sito attivo dell enzima. La catena laterale re dell enzima può comportarsi
10/30/16. non modificato CAP al 5 e poly-a al 3. RNA messaggero: soggetto a splicing
 procarioti eucarioti poli-cistronico mono-cistronico non modificato CAP al 5 e poly-a al 3 RNA messaggero: procarioti eucarioti policistronico monocistronico non modificato CAP al 5 e poly-a al 3 continuo
procarioti eucarioti poli-cistronico mono-cistronico non modificato CAP al 5 e poly-a al 3 RNA messaggero: procarioti eucarioti policistronico monocistronico non modificato CAP al 5 e poly-a al 3 continuo
Strategie catalitiche
 Strategie catalitiche catalisi acido-base catalisi covalente catalisi da metalli Dati molecolari di alcune proteine Citocromo C (umano) Ribonucleasi A (pancreas di bue) Lisozima (bianco dell uovo) Mioglobina
Strategie catalitiche catalisi acido-base catalisi covalente catalisi da metalli Dati molecolari di alcune proteine Citocromo C (umano) Ribonucleasi A (pancreas di bue) Lisozima (bianco dell uovo) Mioglobina
- utilizzano esclusivamente le reattività chimiche di alcuni residui AA
 Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
Enzimi semplici Enzimi coniugati - utilizzano esclusivamente le reattività chimiche di alcuni residui AA - richiedono la reattività chimica aggiuntiva di COFATTORI o COENZIMI gruppi prostetici COENZIMI
TRASDUZIONE DEL SEGNALE CELLULARE
 TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
TRASDUZIONE DEL SEGNALE CELLULARE RECETTORI I recettori ormonali sono proteine (spesso glicoproteine) capaci di riconoscere e legare l ormone L interazione tra ormone e recettore è estremamente specifica
REGOLAZIONE DEL METABOLISMO GLUCIDICO
 REGOLAZIONE DEL METABOLISMO GLUCIDICO IL METABOLISMO DEL GLICOGENO E FINEMENTE REGOLATO: Quando è attiva la sua sintesi non è attiva la sua demolizione e viceversa I principali ormoni che controllano il
REGOLAZIONE DEL METABOLISMO GLUCIDICO IL METABOLISMO DEL GLICOGENO E FINEMENTE REGOLATO: Quando è attiva la sua sintesi non è attiva la sua demolizione e viceversa I principali ormoni che controllano il
Dimensioni di Genomi
 Dimensioni di Genomi plasmids viruses bacteria fungi plants algae insects mollusks bony fish amphibians Il Genoma umano è costituito da circa 3 miliardi di bp e contiene un numero di geni pari a circa
Dimensioni di Genomi plasmids viruses bacteria fungi plants algae insects mollusks bony fish amphibians Il Genoma umano è costituito da circa 3 miliardi di bp e contiene un numero di geni pari a circa
Casi più complicati. inibizione mista (competitiva e non competitiva)
") Casi più complicati inibizione mista (competitiva e non competitiva) reazioni reversibili reazioni a più substrati effetto del ph inibizione mista (competitiva e non competitiva) K I K I E + I EI ES +
Casi più complicati inibizione mista (competitiva e non competitiva) reazioni reversibili reazioni a più substrati effetto del ph inibizione mista (competitiva e non competitiva) K I K I E + I EI ES +
FATTORI DI CRESCITA COME MESSAGGERI
 RECETTORI ASSOCIATI A PROTEINE CHINASI Sono proteine che hanno sia funzione di recettore che attività enzimatica chinasica. L interazione con il ligando specifico stimola l attività della chinasi e il
RECETTORI ASSOCIATI A PROTEINE CHINASI Sono proteine che hanno sia funzione di recettore che attività enzimatica chinasica. L interazione con il ligando specifico stimola l attività della chinasi e il
REGOLAZIONE DELL ESPRESSIONE GENICA. Controllo trascrizionale in E. coli. Esempio: Lac operon
 REGOLAZIONE DELL ESPRESSIONE GENICA Controllo trascrizionale in E. coli Esempio: Lac operon Nel genoma di un batterio ci sono circa 4000 geni Nel genoma umano ci sono circa 25000 geni. Espressione costitutiva:
REGOLAZIONE DELL ESPRESSIONE GENICA Controllo trascrizionale in E. coli Esempio: Lac operon Nel genoma di un batterio ci sono circa 4000 geni Nel genoma umano ci sono circa 25000 geni. Espressione costitutiva:
- Φ e ψ Presenta 3,6 residui per spira -HB between C=O n residue n and NH n+4. - α-helix
 Structure Basic elements - Φ e ψ -60-50. -Presenta 3,6 residui per spira -HB between C=O n residue n and NH n+4. - α-helix Β-sheet Red oxygen Blu nitrogen white hydrogen black Cα Violet lateral chain -at
Structure Basic elements - Φ e ψ -60-50. -Presenta 3,6 residui per spira -HB between C=O n residue n and NH n+4. - α-helix Β-sheet Red oxygen Blu nitrogen white hydrogen black Cα Violet lateral chain -at
BIOCHIMICA APPLICATA e CLINICA
 BIOCHIMICA APPLICATA e CLINICA Regolazione Enzimatica Biosegnalazione Regolazione Ormonale Specializzazioni metaboliche (Biochimica d Organo) Integrazione del metabolismo BIOCHIMICA APPLICATA e CLINICA
BIOCHIMICA APPLICATA e CLINICA Regolazione Enzimatica Biosegnalazione Regolazione Ormonale Specializzazioni metaboliche (Biochimica d Organo) Integrazione del metabolismo BIOCHIMICA APPLICATA e CLINICA
Regolazione enzimatica Isoenzimi
 Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
Regolazione enzimatica Isoenzimi Gli enzimi regolatori nel metabolismo gruppi di enzimi lavorano insieme per produrre una via metabolica in cui il prodotto del primo enzima diventa il substrato del secondo
Regolazione dell espressione genica
 Regolazione dell espressione genica Ogni processo che porta dal DNA alla proteina funzionale è soggetto a regolazione. Il primo processo relativo alla trascrizione è il più importante punto di regolazione
Regolazione dell espressione genica Ogni processo che porta dal DNA alla proteina funzionale è soggetto a regolazione. Il primo processo relativo alla trascrizione è il più importante punto di regolazione
AMMINOACIDI E PROTEINE
 AMMINOACIDI E PROTEINE 1 AMMINOACIDI Gli amminoacidi sono composti organici composti da atomi di carbonio, idrogeno, ossigeno e azoto e in alcuni casi anche da altri elementi come lo zolfo. Gli amminoacidi
AMMINOACIDI E PROTEINE 1 AMMINOACIDI Gli amminoacidi sono composti organici composti da atomi di carbonio, idrogeno, ossigeno e azoto e in alcuni casi anche da altri elementi come lo zolfo. Gli amminoacidi
30/10/2015. Molte proteine sono. intrinsecamente disordinate. o destrutturate (IDP/IUP), cioè non hanno una struttura
, cioè non hanno una struttura") Molte proteine sono intrinsecamente disordinate o destrutturate (IDP/IUP), cioè non hanno una struttura terziaria e/o secondaria stabile. Le IUP sono caratterizzate da - scarso contenuto in aa apolari,
Molte proteine sono intrinsecamente disordinate o destrutturate (IDP/IUP), cioè non hanno una struttura terziaria e/o secondaria stabile. Le IUP sono caratterizzate da - scarso contenuto in aa apolari,
ENZIMI. Un enzima è un catalizzatore (acceleratore) di reazioni biologiche.
 di reazioni biologiche.") ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
ENZIMI ENZIMI Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Catalizzatore = sostanza in grado di accelerare lo svolgimento di una reazione chimica e quindi di aumentarne la sua velocità,
Regolazione dell espressione genica
 Regolazione dell espressione genica definizioni Gene attivato quando viene trascritto in RNA e il suo messaggio tradotto in molecole proteiche specifiche Espressione genica processo complessivo con cui
Regolazione dell espressione genica definizioni Gene attivato quando viene trascritto in RNA e il suo messaggio tradotto in molecole proteiche specifiche Espressione genica processo complessivo con cui
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita
 -traduzione e indirizzamento delle proteine. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita") MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
MFN0366-A1 (I. Perroteau) -traduzione e indirizzamento delle proteine MFN0366-A1 (I. Perroteau) -traduzione delle proteine trna Traduzione: mrna -------> proteine mrna MFN0366-A1 (I. Perroteau) -traduzione
MFN0366-A1 (I. Perroteau) - il nucleo. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita
 - il nucleo. Solo per uso didattico, vietata la riproduzione, la diffusione o la vendita") 1 Cosa contiene il nucleo? Il nucleo non contiene solo DNA, che costituisce solo il 20% del materiale nucleare, ma anche una grande quantità di proteine chiamate nucleoproteine ed RNA. La maggior parte
1 Cosa contiene il nucleo? Il nucleo non contiene solo DNA, che costituisce solo il 20% del materiale nucleare, ma anche una grande quantità di proteine chiamate nucleoproteine ed RNA. La maggior parte
CONTROLLO DELL ESPRESSIONE GENICA NEI PROCARIOTI
 CONTROLLO DELL ESPRESSIONE GENICA NEI PROCARIOTI Unità trascrizionale E. Coli possiede diversi fattori sigma generale shock da calore carenza di azoto sintesi flagellare stress calore e sali Fattori sigma
CONTROLLO DELL ESPRESSIONE GENICA NEI PROCARIOTI Unità trascrizionale E. Coli possiede diversi fattori sigma generale shock da calore carenza di azoto sintesi flagellare stress calore e sali Fattori sigma
Macromolecole Biologiche. La struttura secondaria (III)
") La struttura secondaria (III) Reverse turn Le proteine globulari hanno una forma compatta, dovuta a numerose inversioni della direzione della catena polipeptidica che le compone. Molte di queste inversioni
La struttura secondaria (III) Reverse turn Le proteine globulari hanno una forma compatta, dovuta a numerose inversioni della direzione della catena polipeptidica che le compone. Molte di queste inversioni
Dal Genotipo al Fenotipo
 Dal Genotipo al Fenotipo Dal Fenotipo normale al Fenotipo patologico Regolazione dell espressione genica Figure 7-1 Molecular Biology of the Cell ( Garland Science 2008) Una cellula differenziata contiene
Dal Genotipo al Fenotipo Dal Fenotipo normale al Fenotipo patologico Regolazione dell espressione genica Figure 7-1 Molecular Biology of the Cell ( Garland Science 2008) Una cellula differenziata contiene
Università di Roma Tor Vergata Scienze della Nutrizione Umana Biochimica della Nutrizione - Prof.ssa Luciana Avigliano A.A.
 Università di Roma Tor Vergata Scienze della Nutrizione Umana Biochimica della Nutrizione - Prof.ssa Luciana Avigliano A.A. 2011-12 Nutrienti quali regolatori delle funzioni cellulari AZIONE REGOLATORIA
Università di Roma Tor Vergata Scienze della Nutrizione Umana Biochimica della Nutrizione - Prof.ssa Luciana Avigliano A.A. 2011-12 Nutrienti quali regolatori delle funzioni cellulari AZIONE REGOLATORIA
A systems biology dynamical model of mammalian G1 cell cycle progression
 Genomica Funzionale e Bioinformatica Corso di Algoritmi per la Bioinformatica A systems biology dynamical model of mammalian G1 cell cycle progression Presentazione di : Del Corvo Marcello Matr. n 719970
Genomica Funzionale e Bioinformatica Corso di Algoritmi per la Bioinformatica A systems biology dynamical model of mammalian G1 cell cycle progression Presentazione di : Del Corvo Marcello Matr. n 719970
I meccanismi di catalisi della sintesi di DNA e RNA sono identici
 I meccanismi di catalisi della sintesi di DNA e RNA sono identici U U OH Ribo-nucleotide trifosfato TRASCRIZIONE Non ha bisogno di innesco Inizia a livello di un promotore La RNA polimerasi è totalmente
I meccanismi di catalisi della sintesi di DNA e RNA sono identici U U OH Ribo-nucleotide trifosfato TRASCRIZIONE Non ha bisogno di innesco Inizia a livello di un promotore La RNA polimerasi è totalmente
Trascrizione. Testi Watson et al. Biologia Molecolare del Gene, Zanichelli Nelson et al, Principi di Biochimica di Lehninger, Zanichelli
 Trascrizione Testi Watson et al. Biologia Molecolare del Gene, Zanichelli Nelson et al, Principi di Biochimica di Lehninger, Zanichelli Maurizio Crestani I componen) fondamentali del DNA Polimero di nucleo.di
Trascrizione Testi Watson et al. Biologia Molecolare del Gene, Zanichelli Nelson et al, Principi di Biochimica di Lehninger, Zanichelli Maurizio Crestani I componen) fondamentali del DNA Polimero di nucleo.di
06_citologia_SER_golgi 1
 1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
1 La sintesi proteica inizia sempre nello stesso modo: aggancio della piccola subunità ribosomale al estremità 5 dell mrna. si aggancio la grande subunità ribosomale In corrispondenza del codone di inizio
La struttura della cromatina
 La struttura della cromatina Istoni e nucleosomi La cromatina Il DNA all interno del nucleo eucariotico è associato a delle proteine. Il complesso DNA-proteina si chiama cromatina. Se nuclei interfasici
La struttura della cromatina Istoni e nucleosomi La cromatina Il DNA all interno del nucleo eucariotico è associato a delle proteine. Il complesso DNA-proteina si chiama cromatina. Se nuclei interfasici
Mediatore chimico. Recettore. Trasduzione del segnale. Risposta della cellula
 Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Mediatore chimico Recettore Trasduzione del segnale Risposta della cellula I mediatori chimici sono prodotti da cellule specializzate e sono diffusi nell organismo da apparati di distribuzione Sistemi
Nucleosomi e trascrizione: Rimodellamento cromatinico e regolazione dell'espressione genica
 Nucleosomi e trascrizione: Rimodellamento cromatinico e regolazione dell'espressione genica La trascrizione di un gene richiede delle modifiche nell'organizzazione della cromatina Figura 4.15C Organizzazione
Nucleosomi e trascrizione: Rimodellamento cromatinico e regolazione dell'espressione genica La trascrizione di un gene richiede delle modifiche nell'organizzazione della cromatina Figura 4.15C Organizzazione
RECETTORI INTRACELLULARI
 RECETTORI INTRACELLULARI RECETTORI NUCLEARI (NR) I recettori nucleari legano gli ormoni lipofili. Gli ormoni lipofili sono gli steroidi, i retinoidi, gli ormoni tiroidei e la vitamina D. Gli steroidi e
RECETTORI INTRACELLULARI RECETTORI NUCLEARI (NR) I recettori nucleari legano gli ormoni lipofili. Gli ormoni lipofili sono gli steroidi, i retinoidi, gli ormoni tiroidei e la vitamina D. Gli steroidi e
Modulo 11 biosegnalazione
 Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
Riconoscimento dell antigene
 Riconoscimento dell antigene Risposta immune adattativa La risposta immune innata è molto efficiente ma non sempre riesce ad impedire il diffondersi dell infezione È essenziale quindi che subentri la risposta
Riconoscimento dell antigene Risposta immune adattativa La risposta immune innata è molto efficiente ma non sempre riesce ad impedire il diffondersi dell infezione È essenziale quindi che subentri la risposta
Chimica Biologica A.A α-elica foglietto β reverse turn
 Chimica Biologica A.A. 2010-2011 α-elica foglietto β reverse turn Str. Secondaria sperimentalmente osservata: Si distinguono fondamentalmente tre tipi di strutture secondarie: α elica foglietto β reverse
Chimica Biologica A.A. 2010-2011 α-elica foglietto β reverse turn Str. Secondaria sperimentalmente osservata: Si distinguono fondamentalmente tre tipi di strutture secondarie: α elica foglietto β reverse
Le proteine sono polimeri lineari costituiti da unità base formate da oltre 40 amminoacidi. Possono assumere forme diverse a seconda della funzione
 Le proteine sono polimeri lineari costituiti da unità base formate da oltre 40 amminoacidi Hanno elevato PM Possono assumere forme diverse a seconda della funzione svolgono molteplici funzioni Tra le proteine
Le proteine sono polimeri lineari costituiti da unità base formate da oltre 40 amminoacidi Hanno elevato PM Possono assumere forme diverse a seconda della funzione svolgono molteplici funzioni Tra le proteine
Il sistema immunitario ha sviluppato un sistema altamente specializzato per la cattura degli antigeni e per la loro presentazione ai linfociti.
 Come riescono pochi linfociti specifici per un antigene microbico a localizzare il microbo? Come riesce il sistema immunitario a produrre le cellule effettrici e le molecole piùadatte a debellare un determinato
Come riescono pochi linfociti specifici per un antigene microbico a localizzare il microbo? Come riesce il sistema immunitario a produrre le cellule effettrici e le molecole piùadatte a debellare un determinato
ALCUNI COMPOSTI CHIMICI INIBISCONO SELETTIVAMENTE LA SINTESI PROTEICA NEI PROCARIOTI E NEGLI EUCARIOTI
 ALCUNI COMPOSTI CHIMICI INIBISCONO SELETTIVAMENTE LA SINTESI PROTEICA NEI PROCARIOTI E NEGLI EUCARIOTI VI SONO COMPOSTI SPECIFICI PER OGNI TAPPA DELLA TRADUZIONE Molti microrganismi sono in grado di sviluppare
ALCUNI COMPOSTI CHIMICI INIBISCONO SELETTIVAMENTE LA SINTESI PROTEICA NEI PROCARIOTI E NEGLI EUCARIOTI VI SONO COMPOSTI SPECIFICI PER OGNI TAPPA DELLA TRADUZIONE Molti microrganismi sono in grado di sviluppare
all codons are used in protein synthesis 20 amino acids 3 termination (stop) codons: UAA, UAG, UGA
 codons: UAA, UAG, UGA") Il Codice Genetico The genetic code consists of 64 triplet codons (A, G, C, U) 4 3 = 64 all codons are used in protein synthesis 20 amino acids 3 termination (stop) codons: UAA, UAG, UGA AUG (methionine)
Il Codice Genetico The genetic code consists of 64 triplet codons (A, G, C, U) 4 3 = 64 all codons are used in protein synthesis 20 amino acids 3 termination (stop) codons: UAA, UAG, UGA AUG (methionine)
Modulo 11 biosegnalazione
 Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
Modulo 11 biosegnalazione La trasduzione del segnale Capacità di ricevere e rispondere a segnali esterni alla membrana è un processo fondamentale per la vita. Organismi pluricellulari si scambiano informazioni:
DESTINI METABOLICI DEL PIRUVATO
 DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
DESTINI METABOLICI DEL PIRUVATO Glicolisi Piruvato Metabolismo aerobico: il piruvato entra nel mitocondrio Acetil-CoA Ciclo di Krebs Catena di trasporto degli elettroni Piruvato Complesso della Piruvato
Cenni al controllo dell espressione genica
 Cenni al controllo dell espressione genica Essentials of Cell Biology Unit 2: How Do Cells Decode Genetic Information into Functional Proteins? Biotecnologie_2013 http://www.nature.com/scitable/ebooks/essentials
Cenni al controllo dell espressione genica Essentials of Cell Biology Unit 2: How Do Cells Decode Genetic Information into Functional Proteins? Biotecnologie_2013 http://www.nature.com/scitable/ebooks/essentials
informazione ed espressione genica
 a.a. 2015-16 CORSO DI LAUREA IN INFERMIERISTICA Dott.ssa Marilena Greco Biologia applicata informazione ed espressione genica Biologia Applicata_M.Greco 1 ACIDI NUCLEICI (DNA, RNA) Gli acidi nucleici trasmettono
a.a. 2015-16 CORSO DI LAUREA IN INFERMIERISTICA Dott.ssa Marilena Greco Biologia applicata informazione ed espressione genica Biologia Applicata_M.Greco 1 ACIDI NUCLEICI (DNA, RNA) Gli acidi nucleici trasmettono
E. Giordano 16/09/2010
 GRUPPO NAZIONALE DI BIOINGEGNERIA XXIX Scuola Annuale BIOLOGIA SINTETICA Bressanone 13-17 settembre 2010 1/41 COSTITUENTI MOLECOLARI DELLO CHASSIS CELLULARE Emanuele GIORDANO II Facoltà di Ingegneria Dipartimento
GRUPPO NAZIONALE DI BIOINGEGNERIA XXIX Scuola Annuale BIOLOGIA SINTETICA Bressanone 13-17 settembre 2010 1/41 COSTITUENTI MOLECOLARI DELLO CHASSIS CELLULARE Emanuele GIORDANO II Facoltà di Ingegneria Dipartimento
TRASCRIZIONE DEL DNA. Formazione mrna
 TRASCRIZIONE DEL DNA Formazione mrna Trascrizione Processo mediante il quale l informazione contenuta in una sequenza di DNA (gene) viene copiata in una sequenza complementare di RNA dall enzima RNA polimerasi
TRASCRIZIONE DEL DNA Formazione mrna Trascrizione Processo mediante il quale l informazione contenuta in una sequenza di DNA (gene) viene copiata in una sequenza complementare di RNA dall enzima RNA polimerasi
CONTROLLO E REGOLAZIONE DELLA TRADUZIONE. PROCARIOTI: trascrizione e traduzione avvengono nello stesso compartimento
 CONTROLLO E REGOLAZIONE DELLA TRADUZIONE PROCARIOTI: trascrizione e traduzione avvengono nello stesso compartimento EUCARIOTI:trascrizione e traduzione in compartimenti diversi RISPOSTA STRINGENTE LA SINTESI
CONTROLLO E REGOLAZIONE DELLA TRADUZIONE PROCARIOTI: trascrizione e traduzione avvengono nello stesso compartimento EUCARIOTI:trascrizione e traduzione in compartimenti diversi RISPOSTA STRINGENTE LA SINTESI
HER2 e carcinoma gastrico
 Modulo 1 HER2 e carcinoma gastrico Cosa è HER2 Con l'acronimo HER2 viene identificato uno dei recettori per i fattori epidermici di crescita, codificato dal cromosoma 17; HER2 appartiene alla famiglia
Modulo 1 HER2 e carcinoma gastrico Cosa è HER2 Con l'acronimo HER2 viene identificato uno dei recettori per i fattori epidermici di crescita, codificato dal cromosoma 17; HER2 appartiene alla famiglia
CROMATINA ISTONI. Proteine relativamente piccole, con forte carica positiva per la presenza degli aminoacidi lisina e arginina
 CROMATINA Complesso molecolare formato da DNA, istoni e proteine non istoniche ISTONI Proteine relativamente piccole, con forte carica positiva per la presenza degli aminoacidi lisina e arginina Si conoscono
CROMATINA Complesso molecolare formato da DNA, istoni e proteine non istoniche ISTONI Proteine relativamente piccole, con forte carica positiva per la presenza degli aminoacidi lisina e arginina Si conoscono
Amminoacidi/peptidi/proteine. Chimica Organica II
 Amminoacidi/peptidi/proteine Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II II Amminoacidi: chiralità
Amminoacidi/peptidi/proteine Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II Amminoacidi Chimica Organica II II Amminoacidi: chiralità
LE BASI AZOTATE PIRIMIDINE PURINE
 LE BASI AZTATE PURIE PIRIMIDIE Le basi azotate sono composti eterociclici aromatici con azoti portanti doppietti basici. Possono avere un solo anello (pirimidine) o due (purine). Le basi adenina, guanina
LE BASI AZTATE PURIE PIRIMIDIE Le basi azotate sono composti eterociclici aromatici con azoti portanti doppietti basici. Possono avere un solo anello (pirimidine) o due (purine). Le basi adenina, guanina
Farmacodinamica. Emilio Russo
 Farmacodinamica Emilio Russo Curve concentrazione-risposta Immediata valutazione dell azione biologica Nessuna informazione sul recettore Interferenza di fenomeni esterni (assorbimento, distribuzione
Farmacodinamica Emilio Russo Curve concentrazione-risposta Immediata valutazione dell azione biologica Nessuna informazione sul recettore Interferenza di fenomeni esterni (assorbimento, distribuzione
Organizzazione del DNA negli organismi superiori. La cromatina (DNA + proteine)
") Organizzazione del DNA negli organismi superiori La cromatina (DNA + proteine) Piccole proteine basiche altamente conservate Gli istoni IL NUCLEOSOMA Il nucleosoma è formato da quattro coppie degli istoni
Organizzazione del DNA negli organismi superiori La cromatina (DNA + proteine) Piccole proteine basiche altamente conservate Gli istoni IL NUCLEOSOMA Il nucleosoma è formato da quattro coppie degli istoni
Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI
 Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI Il percorso di maturazione dei linfociti Sviluppo della specicifità immunologica I linfociti B e T avviano le risposte immunitarie dopo il
Immunologia e Immunologia Diagnostica MATURAZIONE DEI LINFOCITI Il percorso di maturazione dei linfociti Sviluppo della specicifità immunologica I linfociti B e T avviano le risposte immunitarie dopo il
3) La presenza di gruppi funzionali specifici che partecipano alla catalisi (quelli delle catene laterali dei suoi residui amminoacidici e/o quelli
 La presenza di gruppi funzionali specifici che partecipano alla catalisi (quelli delle catene laterali dei suoi residui amminoacidici e/o quelli") ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari (sono in grado di aumentare la velocità delle reazioni biologiche anche di 10 20 volte). Durante
ENZIMI Tutti gli enzimi sono PROTEINE che funzionano da catalizzatori biologici nelle reazioni cellulari (sono in grado di aumentare la velocità delle reazioni biologiche anche di 10 20 volte). Durante
lati esterni altamente Idrofilici
 I due filamenti complementari del DNA sono antiparalleli: uno è in direzione 5-3 e l altro in direzione 3-5. parte interna idrofobica lati esterni altamente Idrofilici APPAIAMENTO DELLE BASI AZOTATE: 2
I due filamenti complementari del DNA sono antiparalleli: uno è in direzione 5-3 e l altro in direzione 3-5. parte interna idrofobica lati esterni altamente Idrofilici APPAIAMENTO DELLE BASI AZOTATE: 2
MOLTE PROTEINE SONO LOCALIZZATE IN COMPARTIMENTI DIVERSI DA QUELLO CITOPLASMATICO. Mitocondri Cloroplasti Perossisomi
 MOLTE PROTEINE SONO LOCALIZZATE IN COMPARTIMENTI DIVERSI DA QUELLO CITOPLASMATICO LA TRASLOCAZIONE E IL PASSAGGIO DI UNA PROTEINA ATTRAVERSO UNA MEMBRANA COTRADUZIONALE Batteri Reticolo endoplasmatico
MOLTE PROTEINE SONO LOCALIZZATE IN COMPARTIMENTI DIVERSI DA QUELLO CITOPLASMATICO LA TRASLOCAZIONE E IL PASSAGGIO DI UNA PROTEINA ATTRAVERSO UNA MEMBRANA COTRADUZIONALE Batteri Reticolo endoplasmatico
espressione genica processo attraverso cui l informazione di un gene viene decodificata
 espressione genica processo attraverso cui l informazione di un gene viene decodificata regolazione o controllo dell espressione genica meccanismi mediante i quali un gene viene selettivamente espresso
espressione genica processo attraverso cui l informazione di un gene viene decodificata regolazione o controllo dell espressione genica meccanismi mediante i quali un gene viene selettivamente espresso
CLASSIFICAZIONE O.T.I.L.Is. Lig
 GLI ENZIMI CHE COSA SONO? Sono delle proteine altamente specializzate con attività catalitica, accelerano le reazioni chimiche rimanendo inalterati al termine della reazione stessa. CLASSIFICAZIONE O.T.I.L.Is.
GLI ENZIMI CHE COSA SONO? Sono delle proteine altamente specializzate con attività catalitica, accelerano le reazioni chimiche rimanendo inalterati al termine della reazione stessa. CLASSIFICAZIONE O.T.I.L.Is.